
Levels of Circulating IgM and IgY Natural Antibodies in Broiler Chicks: Association with Genotype and Farming Systems
 , , , , , ,
, , , , , ,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Development of In-House ELISAs
2.3. Statistical Analysis
3. Results
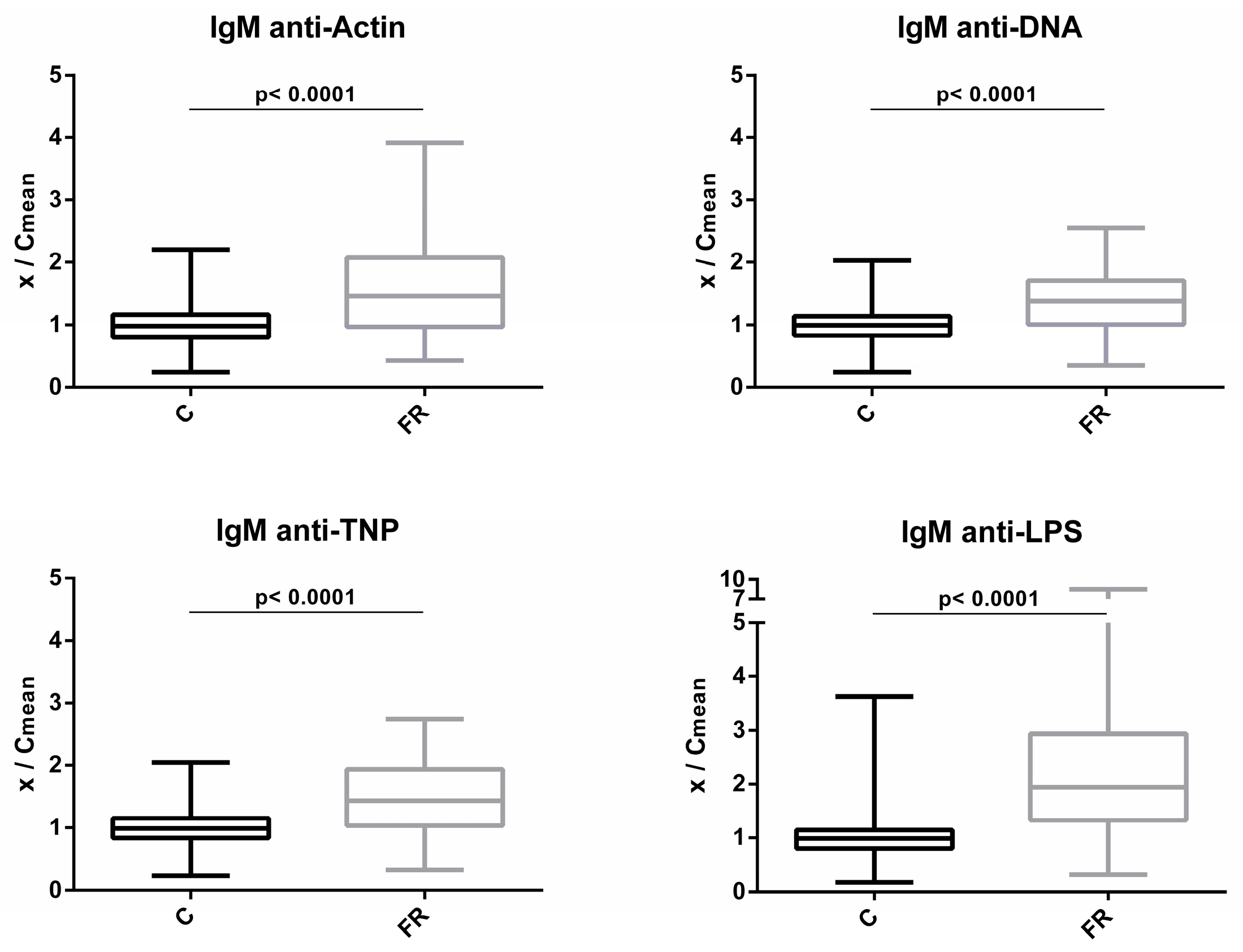
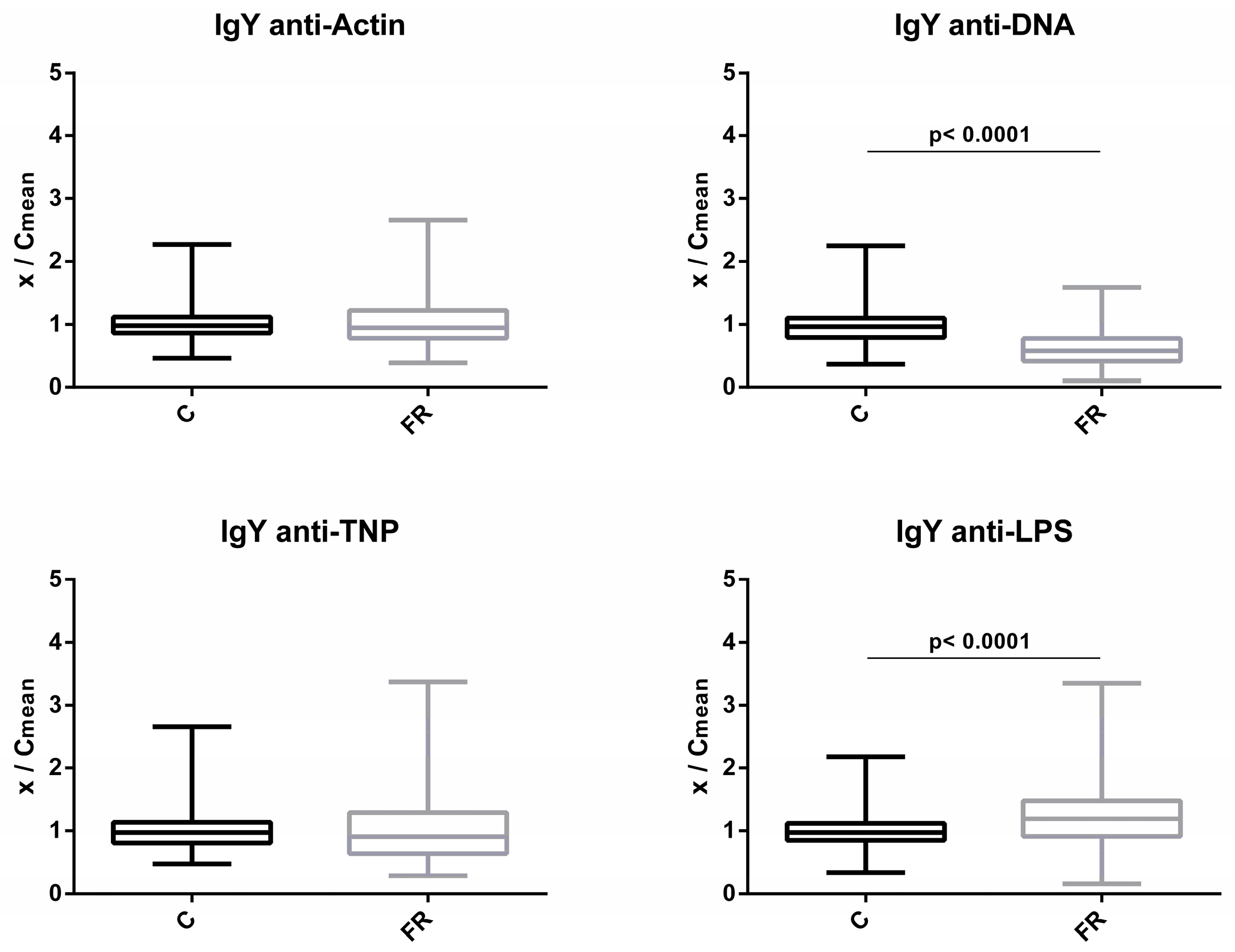
3.1. Comparison of NAb Levels between Free-Range and Conventional Poultry
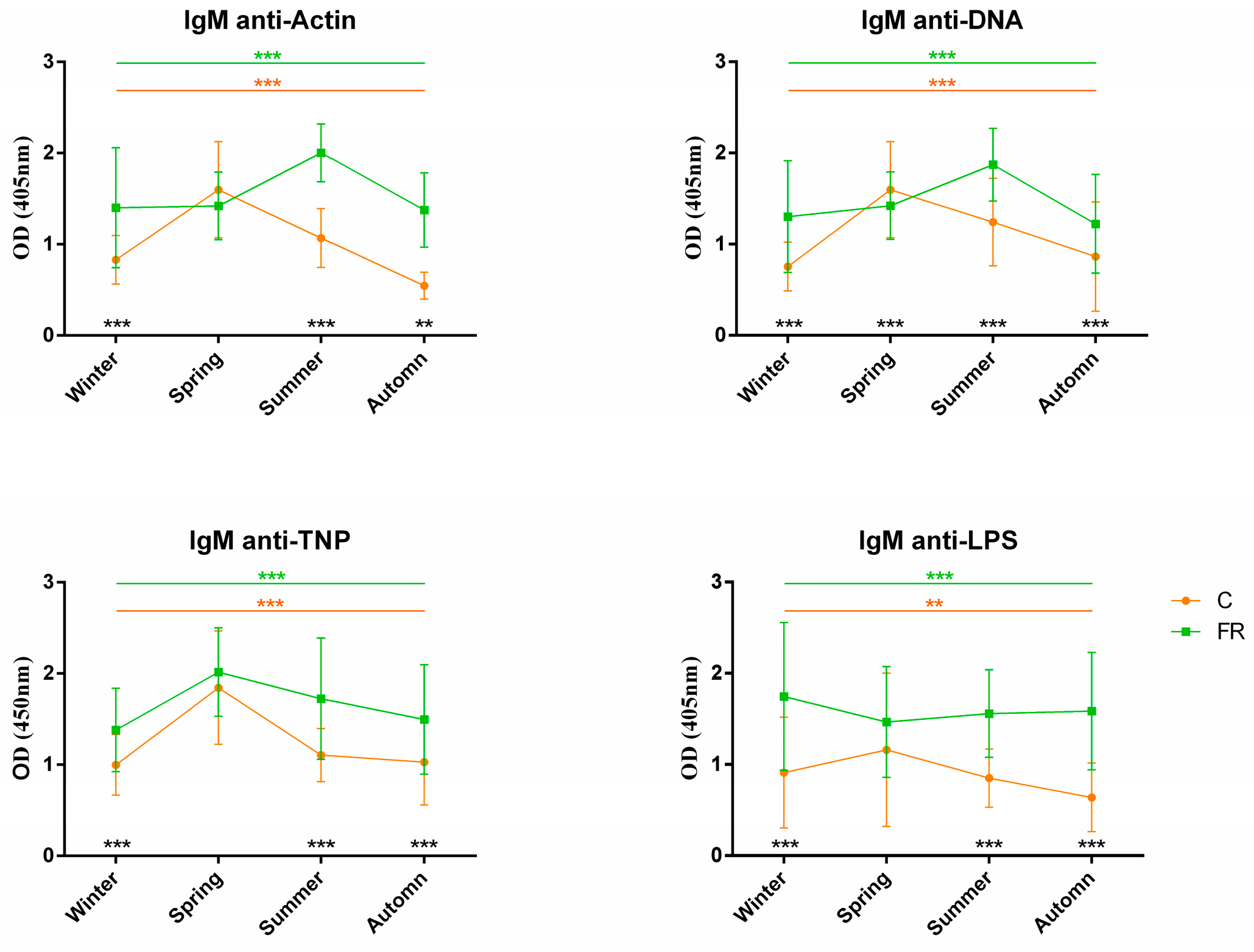
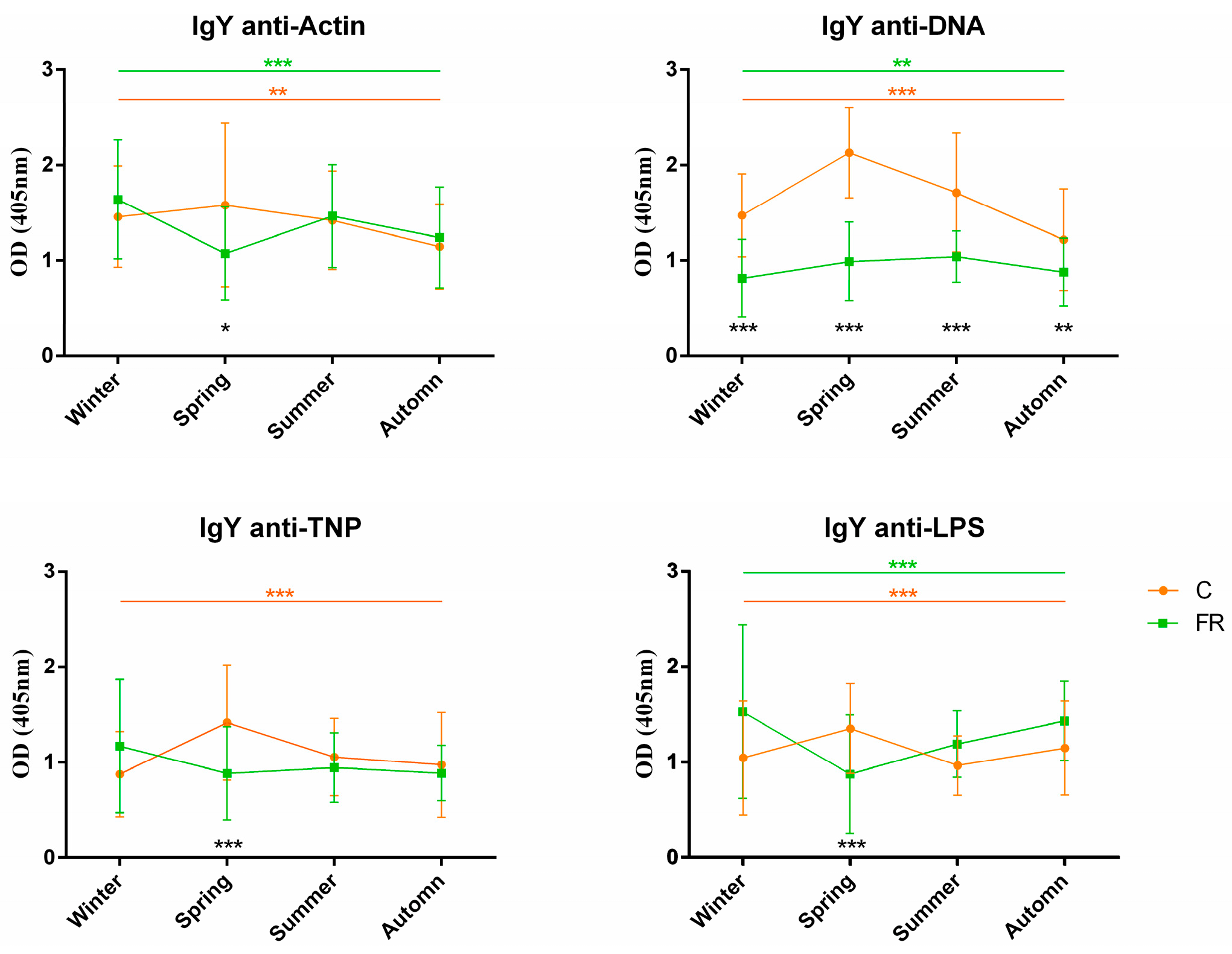
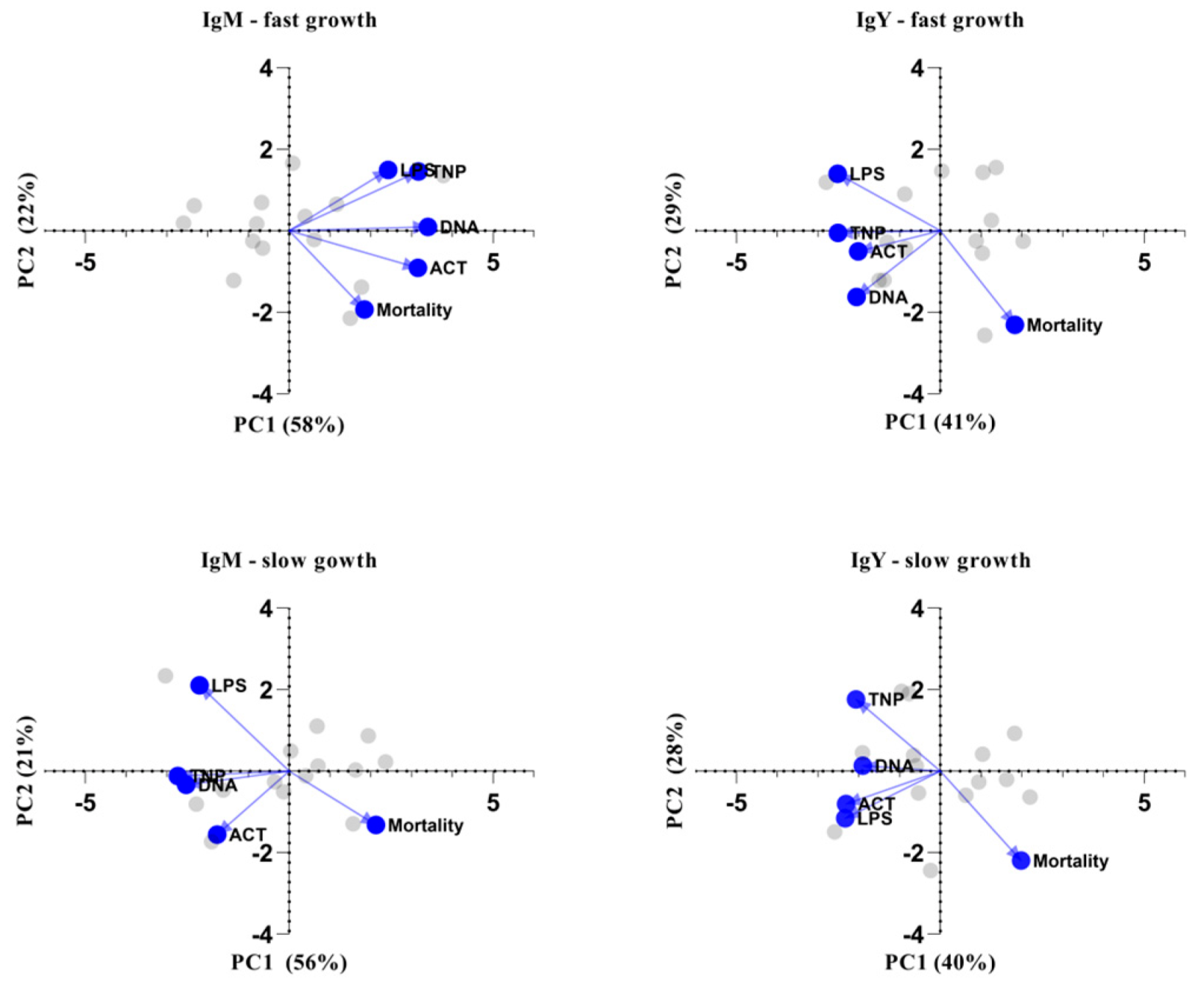
3.2. Association of NAb Levels with Season and Performance
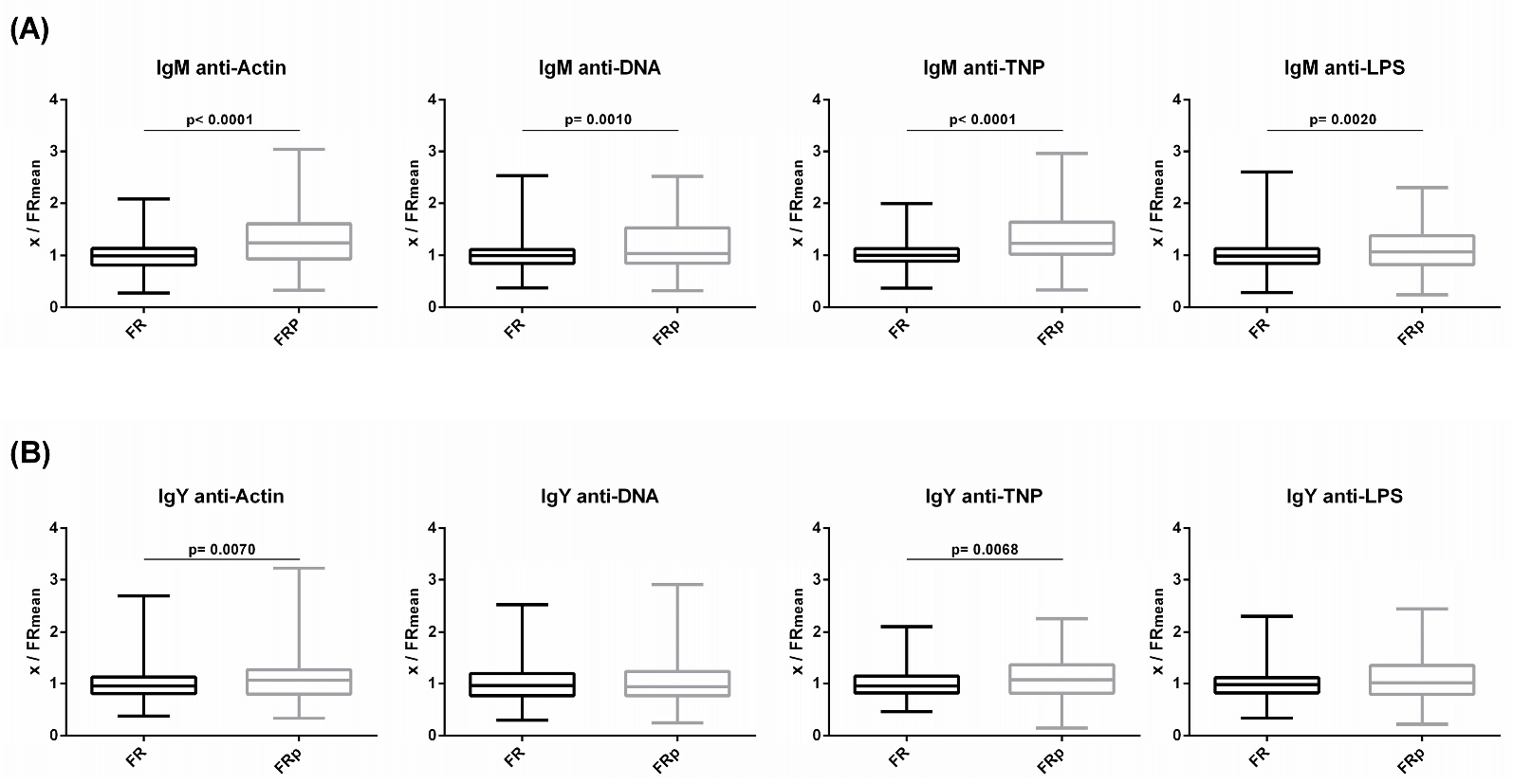
3.3. Comparison of NAb Levels between FR and FRp Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clavijo, V.; Flórez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Hodges, J. Emerging boundaries for poultry production: Challenges, dangers and opportunities. Worlds Poult. Sci. J. 2009, 65, 5–22. [Google Scholar] [CrossRef]
- Hartcher, K.M.; Jones, B. The welfare of layer hens in cage and cage-free housing systems. Worlds Poult. Sci. J. 2017, 73, 767–782. [Google Scholar] [CrossRef]
- Hofmann, T.; Schmucker, S.S.; Bessei, W.; Grashorn, M.; Stefanski, V. Impact of Housing Environment on the Immune System in Chickens: A Review. Animals 2020, 10, 1138. [Google Scholar] [CrossRef] [PubMed]
- Avrameas, S. Natural autoantibodies: From ‘horror autotoxicus’ to ‘gnothi seauton’. Immunol. Today 1991, 12, 154–159. [Google Scholar] [CrossRef]
- Ochsenbein, A.F.; Zinkernagel, R.M. Natural antibodies and complement link innate and acquired immunity. Immunol. Today 2000, 21, 624–630. [Google Scholar] [CrossRef]
- Coutinho, A.; Kazatchkine, M.D.; Avrameas, S. Natural autoantibodies. Curr. Opin. Immunol. 1995, 7, 812–818. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Zhang, Y.; Hu, Y.F.; Wahl, L.M.; Cisar, J.O.; Notkins, A.L. The broad antibacterial activity of the natural antibody repertoire is due to polyreactive antibodies. Cell Host Microb. 2007, 1, 51–61. [Google Scholar] [CrossRef]
- Rhee, K.J.; Sethupathi, P.; Driks, A.; Lanning, D.K.; Knight, K.L. Role of commensal bacteria in development of gut-associated lymphoid tissues and preimmune antibody repertoire. J. Immunol. 2004, 172, 1118–1124. [Google Scholar] [CrossRef]
- Lammers, A.; Klomp, M.E.V.; Nieuwland, M.G.B.; Savelkoul, H.F.J.; Parmentier, H.K. Adoptive transfer of natural antibodies to nonimmunized chickens affects subsequent antigen-specific humoral and cellular immune responses. Dev. Comp. Immunol. 2004, 28, 51–60. [Google Scholar] [CrossRef]
- Berghof, T.V.; De Vries Reilingh, G.; Nieuwland, M.G.; Parmentier, H.K. Effect of aging and repeated intratracheal challenge on levels of cryptic and overt natural antibodies in poultry. Poult. Sci. 2010, 89, 227–235. [Google Scholar] [CrossRef]
- Neu, N.; Hala, K.; Wick, G. “Natural” chicken antibodies to red blood cells are mainly directed against the B-G antigen, and their occurrence is independent of spontaneous autoimmune thyroiditis. Immunogenetics 1984, 19, 269–277. [Google Scholar] [CrossRef]
- Siwek, M.; Buitenhuis, B.; Cornelissen, S.; Nieuwland, M.; Knol, E.F.; Crooijamans, R.; Groenen, M.; Parmentier, H.; van der Poel, J. Detection of QTL for innate: Non-specific antibody levels binding LPS and LTA in two independent populations of laying hens. Dev. Comp. Immunol. 2006, 30, 659–666. [Google Scholar] [CrossRef]
- Ismiraj, M.R.; Arts, J.A.J.; Parmentier, H.K. Maternal Transfer of Natural (Auto-) Antibodies in Chickens. Poult. Sci. 2019, 98, 2380–2391. [Google Scholar] [CrossRef]
- Wondmeneh, E.; Van Arendonk, J.A.M.; Van der Waaij, E.H.; Ducro, B.J.; Parmentier, H.K. High natural antibody titers of indigenous chickens are related with increased hazard in confinement. Poult. Sci. 2015, 94, 1493–1498. [Google Scholar] [CrossRef]
- Sun, Y.; Parmentier, H.K.; Frankena, K.; Van der Poel, J.J. Natural antibody isotypes as predictors of survival in laying hens. Poult. Sci. 2011, 90, 2263–2274. [Google Scholar] [CrossRef]
- Dighiero, G.; Lymberi, P.; Holmberg, D.; Lundquist, I.; Coutinho, A.; Avrameas, S. High frequency of natural autoantibodies in normal newborn mice. J. Immunol. 1985, 134, 765–771. [Google Scholar] [CrossRef]
- Diaz Carrasco, J.M.; Redondo, L.M.; Redondo, E.A.; Dominguez, J.E.; Chacana, A.P.; Fernandez Miyakawa, M.E. Use of Plant Extracts as an Effective Manner to Control Clostridium perfringens Induced Necrotic Enteritis in Poultry. Biomed Res. Int. 2016, 2016, 3278359. [Google Scholar] [CrossRef]
- Redondo, L.M.; Chacana, P.A.; Dominguez, J.E.; Fernandez Miyakawa, M.E. Perspectives in the use of tannins as alternative to antimicrobial growth promoter factors in poultry. Front. Microbiol. 2014, 5, 118. [Google Scholar] [CrossRef]
- Lutz, H.U. Homeostatic roles of naturally occurring antibodies: An overview. J. Autoimmun. 2007, 29, 287–294. [Google Scholar] [CrossRef]
- Gonzalez, R.; Matsiota, P.; Torchy, C.; De Kinkelin, P.; Avrameas, S. Natural anti-TNP antibodies from rainbow trout interfere with viral infection in vitro. Res. Immunol. 1989, 140, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Gunti, S.; Notkins, A.L. Polyreactive Antibodies: Function and Quantification. J. Infect. Dis. 2015, 212 (Suppl. S1), S42–S46. [Google Scholar] [CrossRef] [PubMed]
- EC (European Commission). Council Directive 2007/43/EC of 28 June 2007 Laying down Minimum Rules for the Protection of Chickens Kept for Meat Production (Text with EEA Relevance); European Commission: Brussels, Belgium, 2007. [Google Scholar]
- Bhanja, S.; Bhadauria, P. Behaviour and welfare concepts in laying hens and their association with housing systems. Indian J. Poult. Sci. 2018, 53, 1–10. [Google Scholar] [CrossRef]
- Shimmura, T.; Hirahara, S.; Azuma, T.; Suzuki, T.; Eguchi, Y.; Uetake, K.; Tanaka, T. Multi-factorial investigation of various housing systems for laying hens. Br. Poult. Sci. 2010, 51, 31–42. [Google Scholar] [CrossRef]
- Scholz, B.; Rönchen, S.; Hamann, H.; Pendl, H.; Distl, O. Effect of housing system, group size and perch position on H/L-ratio in laying hens. Eur. Poult. Sci. 2008, 72, 174–180. [Google Scholar]
- Awad, E.A.; Najaa, M.; Zulaikha, Z.A.; Zulkifli, I.; Soleimani, A.F. Effects of heat stress on growth performance, selected physiological and immunological parameters, caecal microflora, and meat quality in two broiler strains. Asian-Australas J. Anim. Sci. 2020, 33, 778–787. [Google Scholar] [CrossRef]
- Baumgarth, N.; Tung, J.W.; Herzenberg, L.A. Inherent specificities in natural antibodies: A key to immune defense against pathogen invasion. Semin. Immunopathol. 2005, 26, 347–362. [Google Scholar] [CrossRef]
- Owen, J.P.; Waite, J.L.; Holden, K.Z.; Clayton, D.H. Does antibody binding to diverse antigens predict future infection? Parasite Immunol. 2014, 36, 573–584. [Google Scholar] [CrossRef]
- Van Altena, S.E.C.; Peen, M.A.; Van der Linden, F.H.; Parmentier, H.K.; Savelkoul, H.F.J.; Tijhaar, E.J. Bovine natural antibodies in antibody-dependent bactericidal activity against Escherichia coli and Salmonella Typhimurium and risk of mastitis. Vet. Immunol. Immunopathol. 2016, 171, 21–27. [Google Scholar] [CrossRef]
- Jayasekera, J.P.; Moseman, E.A.; Carroll, M.C. Natural antibody and complement mediate neutralization of influenza virus in the absence of prior immunity. J. Virol. 2007, 81, 3487–3494. [Google Scholar] [CrossRef]
- Michel, C.; Gonzalez, R.; Bonjour, E.; Avrameas, S. A concurrent increasing of natural antibodies and enhancement of resistance to furunculosis in rainbow trout. Ann. Rech. Vet. 1990, 21, 211–218. [Google Scholar]
- Ujvari, B.; Madsen, T. Do natural antibodies compensate for humoral immunosenescence in tropical pythons? Funct. Ecol. 2011, 25, 813–817. [Google Scholar] [CrossRef]
- Pap, P.; Vágási, C.; Vincze, O.; Osváth, G.; Veres-Szászka, J.; Czirják, G. Physiological pace of life: The link between constitutive immunity, developmental period, and metabolic rate in European birds. Oecologia 2015, 177, 147–158. [Google Scholar] [CrossRef]
- Parmentier, H.K.; Lammers, A.; Hoekman, J.J.; de Vries Reilingh, G.; Zaanen, I.T.; Savelkoul, H.F. Different levels of natural antibodies in chickens divergently selected for specific antibody responses. Dev. Comp. Immunol. 2004, 28, 39–49. [Google Scholar] [CrossRef]
- Reyneveld, G.I.; Savelkoul, H.F.J.; Parmentier, H.K. Current Understanding of Natural Antibodies and Exploring the Possibilities of Modulation Using Veterinary Models. A Review. Front Immunol. 2020, 11, 2139. [Google Scholar] [CrossRef]
- Lymberi, P.; Zannikou, M.; Hatzioannou, A. Natural Autoantibodies in Health and Disease. Compr. Pharmacol. 2022, 342–368. [Google Scholar] [CrossRef]
- Berland, R.; Wortis, H.H. Origins and functions of B-1 cells with notes on the role of CD5. Annu. Rev. Immunol. 2002, 20, 253–300. [Google Scholar] [CrossRef]
- Avrameas, S.; Dighiero, G.; Lymberi, P.; Guilbert, B. Studies on natural antibodies and autoantibodies. Ann. Immunol. 1983, 134D, 103–113. [Google Scholar]
- Mouthon, L.; Lacroix-Desmazes, S.; Nobrega, A.; Barreau, C.; Coutinho, A.; Kazatchkine, M.D. The self-reactive antibody repertoire of normal human serum IgM is acquired in early childhood and remains conserved throughout life. Scand. J. Immunol. 1996, 44, 243–251. [Google Scholar] [CrossRef]
- Hooijkaas, H.; Benner, R.; Pleasants, J.R.; Wostman, B.S. Isotypes and specificities of immunoglobulins produced by germ-free mice fed chemically defined ultrafiltered “antigen-free” diet. Eur. J. Immunol. 1984, 14, 1127–1130. [Google Scholar] [CrossRef]
- Nobrega, A.; Haury, M.; Grandien, A.; Malanchère, E.; Sundblad, A. Global analysis of antibody repertoires., I.I. Evidence for specificity, self-selection and the immunological “homunculus” of antibodies in normal serum. Eur. J. Immunol. 1993, 23, 2851–2859. [Google Scholar] [CrossRef] [PubMed]
- Berghof, T.V.L.; Matthijs, M.G.R.; Arts, J.A.J.; Bovenhuis, H.; Dwars, R.M.; van der Poel, J.J.; Visker, M.; Parmentier, H.K. Selective breeding for high natural antibody level increases resistance to avian pathogenic Escherichia coli (APEC) in chickens. Dev. Comp. Immunol. 2019, 93, 45–57. [Google Scholar] [CrossRef]
- Hollemans, M.S.; de Vries Reilingh, G.; de Vries, S.; Parmentier, H.K.; Lammers, A. Effects of early nutrition and sanitary conditions on antibody levels in early and later life of broiler chickens. Dev. Comp. Immunol. 2021, 117, 103954. [Google Scholar] [CrossRef] [PubMed]
- De Jong, B.G.; Lammers, A.; Oberendorf, L.A.; Nieuwland, M.G.; Savelkoul, H.F.; Parmentier, H.K. Genetic and phenotypic selection affect natural (auto-) antibody reactivity of chickens. PLoS ONE 2013, 8, e72276. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, H.R.; Gong, J.; Gyles, C.L.; Hayes, M.A.; Zhou, H.; Sanei, B.; Chambers, J.R.; Sharif, S. Probiotics stimulate production of natural antibodies in chickens. Clin. Vaccine Immunol. 2006, 13, 975–980. [Google Scholar] [CrossRef]
- Berghof, T.V.L.; Van der Klein, S.A.S.; Arts, J.A.J.; Parmentier, H.K.; Van der Poel, J.J.; Bovenhuis, H. Genetic and non-genetic inheritance of natural antibodies binding keyhole limpet hemocyanin in a purebred layer chicken line. PLoS ONE 2015, 10, e0131088. [Google Scholar] [CrossRef]
- Zerna, G.; Cameron, T.C.; Toet, H.; Spithill, T.W.; Beddoe, T. Bovine Natural Antibody Relationships to Specific Antibodies and Fasciola hepatica Burdens after Experimental Infection and Vaccination with Glutathione S-Transferase. Vet. Sci. 2022, 9, 58. [Google Scholar] [CrossRef]
- Khobondo, J.O.; Nieuwland, M.G.; Webb, L.E.; Bokkers, E.A.; Parmentier, H.K. Natural (auto)antibodies in calves are affected by age and diet. Vet Q. 2015, 35, 64–75. [Google Scholar] [CrossRef]
- De Klerk, B.; Ducro, B.J.; Heuven, H.C.; den Uyl, I.; van Arendonk, J.A.; Parmentier, H.K.; van der Poel, J.J. Phenotypic and genetic relationships of bovine natural antibodies binding keyhole limpet hemocyanin in plasma and milk. J. Dairy Sci. 2015, 98, 2746–2752. [Google Scholar] [CrossRef]
- Van Knegsel, A.T.; de Vries Reilingh, G.; Meulenberg, S.; van den Brand, H.; Dijkstra, J.; Kemp, B.; Parmentier, H.K. Natural antibodies related to energy balance in early lactation dairy cows. J. Dairy Sci. 2007, 90, 5490–5498. [Google Scholar] [CrossRef]
- Chen, Y.; Tibbs-Cortes, L.E.; Ashley, C.; Putz, A.M.; Lim, K.S.; Dyck, M.K.; Fortin, F.; Plastow, G.S.; Dekkers, J.C.M.; Harding, J.C.S. The genetic basis of natural antibody titers of young healthy pigs and relationships with disease resilience. BMC Genom. 2020, 21, 648. [Google Scholar] [CrossRef]
- Zaazaa, A.; Mudalal, S.; Alzuheir, I.; Samara, M.; Jalboush, N.; Fayyad, A.; Petracci, M. The Impact of Thyme and Oregano Essential Oils Dietary Supplementation on Broiler Health, Growth Performance, and Prevalence of Growth-Related Breast Muscle Abnormalities. Animals 2022, 12, 3065. [Google Scholar] [CrossRef]
- Ruan, D.; Fan, Q.; Fouad, A.M.; Sun, Y.; Huang, S.; Wu, A.; Lin, C.; Kuang, Z.; Zhang, C.; Jiang, S. Effects of dietary oregano essential oil supplementation on growth performance, intestinal antioxidative capacity, immunity, and intestinal microbiota in yellow-feathered chickens. J. Anim. Sci. 2021, 99, skab033. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (kg/ton) | Starter (Days 1–17) | Grower (Days 18–35) | Finisher (Days 36–Slaughter) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| C | FR | FRp | C | FR | FRp | C | FR | FRp | |
| Corn | 200 | 329 | 310 | 150 | 423 | 399 | 0 | 650 | 613 |
| Wheat | 392 | 300 | 283 | 478 | 250 | 236 | 714 | 68 | 64 |
| Soya meal | 335 | 314 | 296 | 305 | 273 | 257 | 230 | 236 | 223 |
| Phosphoric acid | 8.5 | 6.0 | 6.0 | 5.8 | 6.8 | 6.4 | 3.8 | 7.0 | 6.0 |
| Limestone | 14 | 14 | 13 | 12 | 13 | 12 | 10 | 11 | 10 |
| Palm oil | 4 | 0 | 0 | 14 | 10 | 9 | 18 | 17 | 16 |
| Soya oil | 25 | 17 | 16 | 19 | 9 | 9 | 10 | 0 | 0 |
| Premix | 20.0 | 19.9 | 18.8 | 16.3 | 15.7 | 14.8 | 14.4 | 11.8 | 11.1 |
| Oregano premix | - | - | 56.6 | - | - | 56.6 | - | - | 57.0 |
| C | FR | FRp | |
|---|---|---|---|
| Mortality% | 3.81 ± 0.29 | 4.34 ± 0.52 | 4.72 ± 0.50 |
| BW | 2.59 ± 0.07 | 2.50 ± 0.06 | 2.46 ± 0.04 |
| FI | 107.00 ± 8.80 | 89.00 ± 5.36 | 88.00 ± 5.29 |
| BWG | 62.00 ± 3.57 a | 36.00 ± 1.81 a | 36.00 ± 1.89 |
| FCR | 1.74 ± 0.02 b | 2.45 ± 0.02 b | 2.48 ± 0.20 |
| EPEF | 312 ± 4.44 b | 145 ± 3.41 b | 141 ± 2.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarrigeorgiou, I.; Stivarou, T.; Tsinti, G.; Patsias, A.; Fotou, E.; Moulasioti, V.; Kyriakou, D.; Tellis, C.; Papadami, M.; Moussis, V.; et al. Levels of Circulating IgM and IgY Natural Antibodies in Broiler Chicks: Association with Genotype and Farming Systems. Biology 2023, 12, 304. https://doi.org/10.3390/biology12020304
Sarrigeorgiou I, Stivarou T, Tsinti G, Patsias A, Fotou E, Moulasioti V, Kyriakou D, Tellis C, Papadami M, Moussis V, et al. Levels of Circulating IgM and IgY Natural Antibodies in Broiler Chicks: Association with Genotype and Farming Systems. Biology. 2023; 12(2):304. https://doi.org/10.3390/biology12020304
Chicago/Turabian StyleSarrigeorgiou, Ioannis, Theodora Stivarou, Gerasimina Tsinti, Apostolos Patsias, Evgenia Fotou, Vasiliki Moulasioti, Dimitra Kyriakou, Constantinos Tellis, Maria Papadami, Vassilios Moussis, and et al. 2023. "Levels of Circulating IgM and IgY Natural Antibodies in Broiler Chicks: Association with Genotype and Farming Systems" Biology 12, no. 2: 304. https://doi.org/10.3390/biology12020304