
A Comprehensive Review of Mammalian Pigmentation: Paving the Way for Innovative Hair Colour-Changing Cosmetics


Abstract
:Simple Summary
Abstract
1. Introduction
2. Development and Cycling of the Hair Follicle Pigmentary Unit
3. Molecular and Cellular Biology of Hair Pigmentation
3.1. Melanosomes Biogenesis
3.1.1. Major Players in Melanosome Biogenesis
Premelanosome Protein (PMEL)
Melan-a Protein (MLANA)
G protein-Coupled Receptor 143 (GPR143)
3.1.2. Protein Trafficking to Melanosomes
3.2. Biochemical Pathway of Melanogenesis
3.2.1. Major Players in Melanin Synthesis
Tyrosinase (TYR)
Tyrosinase-Related Protein 1 (TYRP1)
Cystine Transporters
3.2.2. Control of Melanosomal pH
3.3. Transport and Transfer of Melanosomes
Models of Melanin Transfer
4. Regulation of Follicular Melanogenesis
4.1. Pro-Opiomelanocortin (POMC)-Derived Peptides
4.1.1. Corticotropin-Releasing Factor (CRF)
4.1.2. Thyrotropin-Releasing Hormone (TRH)
4.2. WNT Proteins
4.3. Stem Cell Factor (SCF)
4.4. Endothelin 1 (ET-1)
4.5. Neurotransmitters
4.6. Bone Morphogenetic Proteins
4.7. Oestrogens
4.8. Cell Adhesion Molecules
4.9. Non-Coding RNAs
4.10. Other Regulators
5. Diversity of Human Hair Colour
6. Age-Induced Hair Greying
7. Modification of the Hair Fibre Colour
8. Conclusions
- (1)
- Hair colour is one of the most distinctive human phenotypes, being the subject of extensive studies over the last decades. Hair pigmentation is a manifestation of the presence of pigments called melanins.
- (2)
- The colouration of hair shafts results from precise and sequential interactions between different cell populations in the hair follicle, the mini-organ responsible for hair shaft production. Hair pigmentation is strictly coupled to the hair growth cycle, occurring during anagen.
- (3)
- Three steps are vital for the uniform and accurate pigmentation of hair shafts: melanosome biogenesis in neural crest-derived melanocytes, the biochemical synthesis of melanins (melanogenesis) in melanosomes, and transfer of melanin granules to surrounding keratinocytes for incorporation into the forming hair fibres. Those steps are under complex genetic control, with MITF being the transcription regulator of many genes associated with those processes.
- (4)
- Melanosome biogenesis depends on PMEL, MLANA, and GRP143, which form a protein complex in early stages of maturation assuring the proper organelle composition and structure. The maturation of melanosomes also requires the trafficking of several melanogenic proteins, mediated by adaptor-related protein complexes (AP-1 and AP-3), the biogenesis of lysosomal organelle complexes (BLOC-1 and BLOC-2), SNAREs and RABs.
- (5)
- Melanogenesis in vivo produces two chemically distinct types of melanins: the black-to-brown eumelanin and the reddish-brown to yellow pheomelanin. The initiation of melanogenesis requires L-Tyr, which can be directly transported from the extracellular space or synthesized inside melanocytes through the hydroxylation of L-phenylalanine by PAH.
- (6)
- Upon uptake into melanosomes (possibly by SLC7A5), L-Tyr is converted to L-DOPA, which is subsequently oxidized to DQ. Both reactions are catalysed by tyrosinase, the most important enzyme in the melanogenic pathway; TYRP1 appears to ensure its appropriate processing and stabilization. In the presence of cysteine, DQ is used in the production of pheomelanin. When most cysteine is depleted, DQ enters the synthetic pathway of eumelanin. The levels of cysteine inside melanosomes are controlled by cystinosin and presumably SLC7A11.
- (7)
- Tyrosinase exhibits optimal enzymatic activity at a neutral pH. Since the pH of melanosomes is greatly influenced by their internal ionic equilibrium, its regulation may be achieved through the concerted action of a cascade of ion channels and transporters: V-ATPases, NHEs, OCA2, SLC45A2, SLC24A5, TPC2, and TRPML3, among others.
- (8)
- Melanocytes transfer mature melanosomes via dendritic projections. The motility of melanosomes from the perinuclear to the melanocyte periphery occurs along microtubules. The capture and movement of melanosomes in the network of actin filaments of the dendrites is regulated by the RAB27A–MLPH–MYO5A complex.
- (9)
- Currently, several models can explain the transference of melanin from melanocytes to keratinocytes: exocytic-mediated transfer model, cytophagocytic-mediated transfer model, fusion model, and membrane vesicle-mediated transfer model. Although the exact mechanism of transfer is still unclear, PAR2-mediated phagocytosis seems to be a mostly necessary step.
- (10)
- Melanogenesis is a tightly regulated biochemical pathway. It is under the control of several autocrine, paracrine, and endocrine regulators/factors. POMC-derived peptides α-MSH and β-MSH act on melanocortin receptors, activating the cAMP/PKA pathway, upregulating melanogenesis. The stimulation of melanogenesis by POMC-derived β-END occurs through a PKC-dependent pathway; the activation of PKC is also involved in the melanogenic effect of ET-1 upon binding to EDNRB. Increased synthesis of melanin via the binding of WNT proteins to FDZ receptors involves GSK3β and β-catenin. The positive and negative modulation of melanin synthesis is attained once SCF binds to KIT, with the further activation of MAPK signalling pathways (p38, ERK, JNK). A myriad of neurotransmitters, bone morphogenetic proteins, oestrogens, cell adhesion molecules, and non-coding RNAs has also been implicated in the regulation of melanogenesis.
- (11)
- The diversity of human hair colour arises mostly from the quantity and ratio of eumelanin and pheomelanin produced. Mostly eumelanic phenotypes range from black, dark brown, medium brown, light brown, to blond; all of these phenotypes contain small, nearly constant, amounts of pheomelanin with decreasing contents of eumelanin. Red hair is the only phenotype that contains comparable amounts of eumelanin and pheomelanin.
- (12)
- The amount and ratio of the two human hair pigments are determined by the activity of TYR and cysteine content of melanosomes, being influenced by several polymorphisms in a wide range of pigmentary genes (MC1R, ASIP, SLC45A2, SLC24A5, OCA2 SLC7A11, KITL, TPCN2, TYRP1, HERC2, IRF4, among others).
- (13)
- Grey/white hair is one of the most common signs of aging. It is related to the loss of pigment in hair shafts by virtue of a marked reduction in melanogenically active melanocytes at the hair bulb. Some mechanisms have been proposed for this age-related depletion of bulbar melanocytes: ROS-induced depletion, exhaustion of the MelSC reservoir, and defective activation or migration of MelSCs.
- (14)
- Given the enormous social importance attributed to hair colour, the dyeing of hair is today a common practice. Hair dyeing systems are diverse and can be divided into oxidative and non-oxidative, or according to durability, into temporary, semi-permanent, and permanent. On the downside, they have been identified as the source of various adverse health effects and thus, the development of safer ways for hair colour modification is more than ever a pertinent issue.
- (15)
- Hair lightening, darkening, and repigmentation have been reported as a side effect of many drugs used in the treatment of several diseases. These findings raise the possibility of interfering with the physiological process of hair pigmentation as an alternative or in addition to conventional methods for hair colour modification. In this sense, the identification and comprehension of the events underlying the pigmentation of hair is an important starting point for the development of such innovative hair cosmetics to change colour from inside out.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- da França, S.A.; Dario, M.F.; Esteves, V.B.; Baby, A.R.; Velasco, M.V.R. Types of Hair Dye and Their Mechanisms of Action. Cosmetics 2015, 2, 110–126. [Google Scholar] [CrossRef]
- Kim, K.H.; Kabir, E.; Jahan, S.A. The Use of Personal Hair Dye and Its Implications for Human Health. Environ. Int. 2016, 89–90, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.; Sinclair, R. Hair Colouring, Permanent Styling and Hair Structure. J. Cosmet. Dermatol. 2003, 2, 180–185. [Google Scholar] [CrossRef]
- Ahn, H.J.; Lee, W.S. An Ultrastuctural Study of Hair Fiber Damage and Restoration Following Treatment with Permanent Hair Dye. Int. J. Dermatol. 2002, 41, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Tosti, A.; Piraccini, B.M.; van Neste, D.J.J. Telogen Effluvium after Allergic Contact Dermatitis of the Scalp. Arch. Dermatol. 2001, 137, 187–190. [Google Scholar]
- Ishida, W.; Makino, T.; Shimizu, T. Severe Hair Loss of the Scalp Due to a Hair Dye Containing Para Phenylenediamine. ISRN Dermatol. 2011, 2011, 947284. [Google Scholar] [CrossRef]
- Chan, H.P.; Maibach, H.I. Hair Highlights and Severe Acute Irritant Dermatitis (“burn”) of the Scalp. Cutan. Ocul. Toxicol. 2010, 29, 229–233. [Google Scholar] [CrossRef]
- Davari, P.; Maibach, H.I. Contact Urticaria to Cosmetic and Industrial Dyes. Clin. Exp. Dermatol. 2011, 36, 1–5. [Google Scholar] [CrossRef]
- Zhang, Y.; Kim, C.; Zheng, T. Hair Dye Use and Risk of Human Cancer. Front. Biosci. 2012, 4, 516. [Google Scholar] [CrossRef]
- Towle, K.M.; Grespin, M.E.; Monnot, A.D. Personal Use of Hair Dyes and Risk of Leukemia: A Systematic Literature Review and Meta-Analysis. Cancer Med. 2017, 6, 2471–2486. [Google Scholar] [CrossRef]
- Eberle, C.E.; Sandler, D.P.; Taylor, K.W.; White, A.J. Hair Dye and Chemical Straightener Use and Breast Cancer Risk in a Large US Population of Black and White Women. Int. J. Cancer 2020, 147, 383–391. [Google Scholar] [CrossRef]
- Xu, S.; Wang, H.; Liu, Y.; Zhang, C.; Xu, Y.; Tian, F.; Mei, L. Hair Chemicals May Increase Breast Cancer Risk: A Meta-Analysis of 210319 Subjects from 14 Studies. PLoS ONE 2021, 16, e0243792. [Google Scholar] [CrossRef]
- Chang, C.-J.; O’brien, K.M.; Keil, A.P.; Gaston, S.A.; Jackson, C.L.; Sandler, D.P.; White, A.J.; Branch, E. Use of Straighteners and Other Hair Products and Incident Uterine Cancer. J. Natl. Cancer Inst. 2022, 114, 1636–1645. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Diversity of Human Hair Pigmentation as Studied by Chemical Analysis of Eumelanin and Pheomelanin. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 1369–1380. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Human Hair Melanins: What We Have Learned and Have Not Learned from Mouse Coat Color Pigmentation. Pigment Cell Melanoma Res. 2010, 24, 63–74. [Google Scholar] [CrossRef]
- Van Neste, D.; Tobin, D.J. Hair Cycle and Hair Pigmentation: Dynamic Interactions and Changes Associated with Aging. Micron 2004, 35, 193–200. [Google Scholar] [CrossRef]
- Tobin, D.J. The Cell Biology of Human Hair Follicle Pigmentation. Pigment Cell Melanoma Res. 2011, 24, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J.; Hordinsky, M.; Bernard, B.A. Hair Pigmentation: A Research Update. J. Investig. Dermatol. Symp. Proc. 2005, 10, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hong, L.; Wakamatsu, K.; Ito, S.; Adhyaru, B.; Cheng, C.-Y.; Bowers, C.R.; Simon, J.D. Comparison of Structural and Chemical Properties of Black and Red Human Hair Melanosomes. Photochem. Photobiol. 2005, 81, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. Human Hair Pigmentation—Biological Aspects. Int. J. Cosmet. Sci. 2008, 30, 233–257. [Google Scholar] [CrossRef]
- Ito, S. A Chemist’s View of Melanogenesis. Pigment Cell Res. 2003, 16, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.; Wortsman, J.; Plonka, P.M.; Schallreuter, K.U.; Paus, R.; Tobin, D.J. Hair Follicle Pigmentation. J. Investig. Dermatol. 2005, 124, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, N.; Berx, G. From Neural Crest Cells to Melanocytes: Cellular Plasticity during Development and Beyond. Cell Mol. Life Sci. 2019, 76, 1919–1934. [Google Scholar] [CrossRef] [PubMed]
- Mayer, T.C. The Migratory Pathway of Neural Crest Cells into the Skin of Mouse Embryos. Dev. Biol. 1973, 34, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Dupin, E.; Le Douarin, N.M. Development of Melanocyte Precursors from the Vertebrate Neural Crest. Oncogene 2003, 22, 3016–3023. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.J.; Erickson, C.A. The Making of a Melanocyte: The Specification of Melanoblasts from the Neural Crest. Pigment Cell Melanoma Res. 2008, 21, 598–610. [Google Scholar] [CrossRef]
- Steingrímsson, E.; Copeland, N.G.; Jenkins, N.A. Melanocytes and the Microphthalmia Transcription Factor Network. Annu. Rev. Genet. 2004, 38, 365–441. [Google Scholar] [CrossRef] [PubMed]
- Mort, R.L.; Jackson, I.J.; Elizabeth Patton, E. The Melanocyte Lineage in Development and Disease. Development 2015, 142, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Opdecamp, K.; Nakayama, A.; Nguyen, M.T.T.; Hodgkinson, C.A.; Pavan, W.J.; Arnheiter, H. Melanocyte Development in Vivo and in Neural Crest Cell Cultures: Crucial Dependence on the Mitf Basic-Helix-Loop-Helix-Zipper Transcription Factor. Development 1997, 124, 2377–2386. [Google Scholar] [CrossRef]
- Nakayama, A.; Nguyen, M.T.T.; Chen, C.C.; Opdecamp, K.; Hodgkinson, C.A.; Arnheiter, H. Mutations in Microphthalmia, the Mouse Homolog of the Human Deafness Gene MITF, Affect Neuroepithelial and Neural Crest-Derived Melanocytes Differently. Mech. Dev. 1998, 70, 155–166. [Google Scholar] [CrossRef]
- Hou, L.; Panthier, J.J.; Arnheiter, H. Signaling and Transcriptional Regulation in the Neural Crest-Derived Melanocyte Lineage: Interactions between KIT and MITF. Development 2000, 127, 5379–5389. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.J.; Erickson, C.A. FOXD3 Regulates the Lineage Switch between Neural Crest-Derived Glial Cells and Pigment Cells by Repressing MITF through a Non-Canonical Mechanism. Development 2009, 136, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- Adameyko, I.; Lallemend, F.; Furlan, A.; Zinin, N.; Aranda, S.; Kitambi, S.S.; Blanchart, A.; Favaro, R.; Nicolis, S.; Lübke, M.; et al. Sox2 and Mitf Cross-Regulatory Interactions Consolidate Progenitor and Melanocyte Lineages in the Cranial Neural Crest. Development 2012, 139, 397–410. [Google Scholar] [CrossRef]
- Kos, R.; Reedy, M.V.; Johnson, R.L.; Erickson, C.A. The Winged-Helix Transcription Factor FoxD3 Is Important for Establishing the Neural Crest Lineage and Repressing Melanogenesis in Avian Embryos. Development 2001, 128, 1467–1479. [Google Scholar] [CrossRef]
- Ignatius, M.S.; Moose, H.E.; El-Hodiri, H.M.; Henion, P.D. Colgate/Hdac1 Repression of Foxd3 Expression Is Required To Permit Mitfa-Dependent Melanogenesis. Dev. Biol. 2008, 313, 568–583. [Google Scholar] [CrossRef]
- Jin, E.J.; Erickson, C.A.; Takada, S.; Burrus, L.W. Wnt and BMP Signaling Govern Lineage Segregation of Melanocytes in the Avian Embryo. Dev. Biol. 2001, 233, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Yasumoto, K.I.; Takada, R.; Takada, S.; Watanabe, K.I.; Udono, T.; Saito, H.; Takahashi, K.; Shibahara, S. Induction of Melanocyte-Specific Microphthalmia-Associated Transcription Factor by Wnt-3a. J. Biol. Chem. 2000, 275, 14013–14016. [Google Scholar] [CrossRef]
- Kawakami, T.; Kimura, S.; Kawa, Y.; Kato, M.; Mizoguchi, M.; Soma, Y. BMP-4 Upregulates Kit Expression in Mouse Melanoblasts Prior to the Kit-Dependent Cycle of Melanogenesis. J. Investig. Dermatol. 2008, 128, 1220–1226. [Google Scholar] [CrossRef]
- Lahav, R.; Dupin, E.; Lecoin, L.; Glavieux, C.; Champeval, D.; Ziller, C.; Le Douarin, N.M. Endothelin 3 Selectively Promotes Survival and Proliferation of Neural Crest-Derived Glial and Melanocytic Precursors in Vitro. Proc. Natl. Acad. Sci. USA 1998, 95, 14214–14219. [Google Scholar] [CrossRef]
- Cable, J.; Jackson, I.J.; Steel, K.P. Mutations at the W Locus Affect Survival of Neural Crest-Derived Melanocytes in the Mouse. Mech. Dev. 1995, 50, 139–150. [Google Scholar] [CrossRef]
- Wehrle-Haller, B.; Weston, J.A. Soluble and Cell-Bound Forms of Steel Factor Activity Play Distinct Roles in Melanocyte Precursor Dispersal and Survival on the Lateral Neural Crest Migration Pathway. Development 1995, 121, 731–742. [Google Scholar] [CrossRef] [PubMed]
- McGill, G.G.; Horstmann, M.; Widlund, H.R.; Du, J.; Motyckova, G.; Nishimura, E.K.; Lin, Y.L.; Ramaswamy, S.; Avery, W.; Ding, H.F.; et al. Bcl2 Regulation by the Melanocyte Master Regulator Mitf Modulates Lineage Survival and Melanoma Cell Viability. Cell 2002, 109, 707–718. [Google Scholar] [CrossRef]
- Baynash, A.G.; Hosoda, K.; Giaid, A.; Richardson, J.A.; Emoto, N.; Hammer, R.E.; Yanagisawa, M. Interaction of Endothelin-3 with Endothelin-B Receptor Is Essential for Development of Epidermal Melanocytes and Enteric Neurons. Cell 1994, 79, 1277–1285. [Google Scholar] [CrossRef]
- Holbrook, K.A.; Underwood, R.A.; Vogel, A.M.; Gown, A.M.; Kimball, H. The Appearance, Density and Distribution of Melanocytes in Human Embryonic and Fetal Skin Revealed by the Anti-Melanoma Monoclonal Antibody, HMB-45. Anat. Embryol. 1989, 180, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Peters, E.M.J.; Tobin, D.J.; Botchkareva, N.; Maurer, M.; Paus, R. Migration of Melanoblasts into the Developing Murine Hair Follicle Is Accompanied by Transient C-Kit Expression. J. Histochem. Cytochem. 2002, 50, 751–766. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.A.; Jackson, I.J. MGF (KIT Ligand) Is a Chemokinetic Factor for Melanoblast Migration into Hair Follicles. Dev. Biol. 2000, 225, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J.; Bystryn, J.C. Different Populations of Melanocytes Are Present in Hair Follicles and Epidermis. Pigment Cell Res. 1996, 9, 304–310. [Google Scholar] [CrossRef]
- Belmadani, A.; Jung, H.; Ren, D.; Miller, R.J. The Chemokine SDF-1/CXCL12 Regulates the Migration of Melanocyte Progenitors in Mouse Hair Follicles. Differentiation 2009, 77, 395–411. [Google Scholar] [CrossRef]
- Botchkareva, N.V.; Khlgatian, M.; Longley, B.J.; Botchkarev, V.A.; Gilchrest, B.A. SCF/C-kit Signaling Is Required for Cyclic Regeneration of the Hair Pigmentation Unit. FASEB J. 2001, 15, 645–658. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin Pigmentation in Mammalian Skin and Its Hormonal Regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Tobin, D.J.; Paus, R. Graying: Gerontobiology of the Hair Follicle Pigmentary Unit. Exp. Gerontol. 2001, 36, 29–54. [Google Scholar] [CrossRef] [PubMed]
- Christoph, T.; Müller-Röver, S.; Audring, H.; Tobin, D.J.; Hermes, B.; Cotsarelis, G.; Rückert, R.; Paus, R. The Human Hair Follicle Immune System: Cellular Composition and Immune Privilege. Br. J. Dermatol. 2000, 142, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Houschyar, K.S.; Borrelli, M.R.; Tapking, C.; Popp, D.; Puladi, B.; Ooms, M.; Chelliah, M.P.; Rein, S.; Pförringer, D.; Thor, D.; et al. Molecular Mechanisms of Hair Growth and Regeneration: Current Understanding and Novel Paradigms. Dermatology 2020, 236, 271–280. [Google Scholar] [CrossRef]
- Slominski, A.; Paus, R.; Plonka, P.; Chakraborty, A.; Maurer, M.; Pruski, D.; Lukiewicz, S. Melanogenesis during the Anagen-Catagen-Telogen Transformation of the Murine Hair Cycle. J. Investig. Dermatol. 1994, 102, 862–869. [Google Scholar] [CrossRef]
- Tobin, D.J.; Slominski, A.; Botchkarev, V.; Paus, R. The Fate of Hair Follicle Melanocytes during the Hair Growth Cycle. J. Investig. Dermatol. Symp. Proc. 1999, 4, 323–332. [Google Scholar] [CrossRef]
- Krause, K.; Foitzik, K. Biology of the Hair Follicle: The Basics. Semin. Cutan. Med. Surg. 2006, 25, 2–10. [Google Scholar] [CrossRef]
- Commo, S.; Bernard, B.A. Melanocyte Subpopulation Turnover During the Human Hair Cycle: An Immunohistochemical Study. Pigment Cell Res. 2000, 13, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Schallreuter, K.U.; Beazley, W.D.; Hibberts, N.A.; Tobin, D.J.; Paus, R.; Wood, J.M. Pterins in Human Hair Follicle Cells and in the Synchronized Murine Hair Cycle. J. Investig. Dermatol. 1998, 111, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Paus, R.; Constantino, R. Differential Expression and Activity of Melanogenesis-Related Proteins During Induced Hair Growth in Mice. J. Investig. Dermatol. 1991, 96, 172–179. [Google Scholar] [CrossRef]
- Tobin, D.J.; Hagen, E.; Botchkarev, V.A.; Paus, R. Do Hair Bulb Melanocytes Undergo Apotosis during Hair Follicle Regression (Catagen)? J. Investig. Dermatol. 1998, 111, 941–947. [Google Scholar] [CrossRef]
- Lindner, G.; Botchkarev, V.A.; Botchkareva, N.V.; Ling, G.; Van Der Veen, C.; Paus, R. Analysis of Apoptosis during Hair Follicle Regression (Catagen). Am. J. Pathol. 1997, 151, 1601–1617. [Google Scholar]
- Mak, S.S.; Moriyama, M.; Nishioka, E.; Osawa, M.; Nishikawa, S.I. Indispensable Role of Bcl2 in the Development of the Melanocyte Stem Cell. Dev. Biol. 2006, 291, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, M.; Osawa, M.; Mak, S.S.; Ohtsuka, T.; Yamamoto, N.; Han, H.; Delmas, V.; Kageyama, R.; Beermann, F.; Larue, L.; et al. Notch Signaling via Hes1 Transcription Factor Maintains Survival of Melanoblasts and Melanocyte Stem Cells. J. Cell Biol. 2006, 173, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Kubic, J.D.; Young, K.P.; Plummer, R.S.; Ludvik, A.E.; Lang, D. Pigmentation PAX-Ways: The Role of Pax3 in Melanogenesis, Melanocyte Stem Cell Maintenance, and Disease. Pigment Cell Melanoma Res. 2008, 21, 627–645. [Google Scholar] [CrossRef]
- Nishimura, E.K. Melanocyte Stem Cells: A Melanocyte Reservoir in Hair Follicles for Hair and Skin Pigmentation. Pigment Cell Melanoma Res. 2011, 24, 401–410. [Google Scholar] [CrossRef]
- Geyfman, M.; Plikus, M.V.; Treffeisen, E.; Andersen, B.; Paus, R. Resting No More: Re-Defining Telogen, the Maintenance Stage of the Hair Growth Cycle. Biol. Rev. 2015, 90, 1179–1196. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.A.; Westgate, G.E.; Jahoda, C.A.B. From Telogen to Exogen: Mechanisms Underlying Formation and Subsequent Loss of the Hair Club Fiber. J. Investig. Dermatol. 2009, 129, 2100–2108. [Google Scholar] [CrossRef] [PubMed]
- Milner, Y.; Sudnik, J.; Filippi, M.; Kizoulis, M.; Kashgarian, M.; Stenn, K. Exogen, Shedding Phase of the Hair Growth Cycle: Characterization of a Mouse Model. J. Investig. Dermatol. 2002, 119, 639–644. [Google Scholar] [CrossRef]
- Du, J.; Miller, A.J.; Widlund, H.R.; Horstmann, M.A.; Ramaswamy, S.; Fisher, D.E. MLANA/MART1 and SILV/PMEL17/GP100 Are Transcriptionally Regulated by MITF in Melanocytes and Melanoma. Am. J. Pathol. 2003, 163, 333–343. [Google Scholar] [CrossRef]
- Vetrini, F.; Auricchio, A.; Du, J.; Angeletti, B.; Fisher, D.E.; Ballabio, A.; Marigo, V. The Microphthalmia Transcription Factor (Mitf) Controls Expression of the Ocular Albinism Type 1 Gene: Link between Melanin Synthesis and Melanosome Biogenesis. Mol. Cell Biol. 2004, 24, 6550–6559. [Google Scholar] [CrossRef]
- Bentley, N.J.; Eisen, T.; Goding, C.R. Melanocyte-Specific Expression of the Human Tyrosinase Promoter: Activation by the Microphthalmia Gene Product and Role of the Initiator. Mol. Cell Biol. 1994, 14, 7996–8006. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, K.; Yokoyama, K.; Shibata, K.; Tomita, Y.; Shibahara, S. Microphthalmia-Associated Transcription Factor as a Regulator for Melanocyte-Specific Transcription of the Human Tyrosinase Gene. Microphthalmia-Associated Transcription Factor as a Regulator for Melanocyte-Specific Transcription of the Human Tyrosinase. Mol. Cell Biol. 1994, 14, 8058–8070. [Google Scholar] [CrossRef]
- Yasumoto, K.I.; Yokoyama, K.; Takahashi, K.; Tomita, Y.; Shibahara, S. Functional Analysis of Microphthalmia-Associated Transcription Factor in Pigment Cell-Specific Transcription of the Human Tyrosinase Family Genes. J. Biol. Chem. 1997, 272, 503–509. [Google Scholar] [CrossRef]
- Yasumoto, K.; Mahalingam, H.; Yokoyama, K.; Shibahara, S. Transcriptional Activation of the Melanocyte-Specific Genes by the Human Homolog of the Mouse Microphthalmia Protein. J. Biochem. 1995, 881, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Bertolotto, C.; Buscà, R.; Abbe, P.; Bille, K.; Aberdam, E.; Ortonne, J.-P.P.; Ballotti, R. Different Cis-Acting Elements Are Involved in the Regulation of TRP1 and TRP2 Promoter Activities by Cyclic AMP: Pivotal Role of M Boxes (GTCATGTGCT) and of Microphthalmia. Mol. Cell Biol. 1998, 18, 694–702. [Google Scholar] [CrossRef]
- Cheli, Y.; Ohanna, M.; Ballotti, R.; Bertolotto, C. Fifteen-Year Quest for Microphthalmia-Associated Transcription Factor Target Genes. Pigment Cell Melanoma Res. 2010, 23, 27–40. [Google Scholar] [CrossRef]
- Du, J.; Fisher, D.E. Identification of Aim-1 as the Underwhite Mouse Mutant and Its Transcriptional Regulation by MITF. J. Biol. Chem. 2002, 277, 402–406. [Google Scholar] [CrossRef]
- Bahadoran, P.; Aberdam, E.; Mantoux, F.; Buscà, R.; Bille, K.; Yalman, N.; de Saint-Basile, G.; Casaroli-Marano, R.; Ortonne, J.P.; Ballotti, R. Rab27a: A Key to Melanosome Transport in Human Melanocytes. J. Cell Biol. 2001, 152, 843–850. [Google Scholar] [CrossRef]
- Aoki, H.; Moro, O. Involvement of Microphthalmia-Associated Transcription Factor (MITF) in Expression of Human Melanocortin-1 Receptor (MC1R). Life Sci. 2002, 71, 2171–2179. [Google Scholar] [CrossRef] [PubMed]
- Sato-Jin, K.; Nishimura, E.K.; Akasaka, E.; Huber, W.; Nakano, H.; Miller, A.; Du, J.; Wu, M.; Hanada, K.; Sawamura, D.; et al. Epistatic Connections between Microphthalmia-associated Transcription Factor and Endothelin Signaling in Waardenburg Syndrome and Other Pigmentary Disorders. FASEB J. 2008, 22, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Hearing, V.J. Update on the Regulation of Mammalian Melanocyte Function and Skin Pigmentation. Expert Rev. Dermatol. 2011, 6, 97–108. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Kothari, S.; Chavan, B.; Spencer, J.D. Regulation of Melanogenesis-Controversies and New Concepts. Exp. Dermatol. 2008, 17, 395–404. [Google Scholar] [CrossRef]
- Chen, T.; Zhao, B.; Liu, Y.; Wang, R.; Yang, Y.; Yang, L.; Dong, C. MITF-M Regulates Melanogenesis in Mouse Melanocytes. J. Dermatol. Sci. 2018, 90, 253–262. [Google Scholar] [CrossRef]
- Sato, S.; Roberts, K.; Gambino, G.; Cook, A.; Kouzarides, T.; Goding, C.R. CBP/P300 as a Co-Factor for the Microphthalmia Transcription Factor. Oncogene 1997, 14, 3083–3092. [Google Scholar] [CrossRef] [PubMed]
- Serre, C.; Busuttil, V.; Botto, J.M. Intrinsic and Extrinsic Regulation of Human Skin Melanogenesis and Pigmentation. Int. J. Cosmet. Sci. 2018, 40, 328–347. [Google Scholar] [CrossRef] [PubMed]
- Shibahara, S.; Yasumoto, K.; Amae, S.; Udono, T.; Watanabe, K. Regulation of Pigment Cell-Specific Gene Expression by MITF. Pigment. Cell Res. 2000, 13, 98–102. [Google Scholar] [CrossRef]
- Vachtenheim, J.; Borovanský, J. “Transcription Physiology” of Pigment Formation in Melanocytes: Central Role of MITF. Exp. Dermatol. 2010, 19, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Levy, C.; Khaled, M.; Fisher, D.E. MITF: Master Regulator of Melanocyte Development and Melanoma Oncogene. Trends Mol. Med. 2006, 12, 406–414. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef]
- Denecker, G.; Vandamme, N.; Akay, Ö.; Koludrovic, D.; Taminau, J.; Lemeire, K.; Gheldof, A.; De Craene, B.; Van Gele, M.; Brochez, L.; et al. Identification of a ZEB2-MITF-ZEB1 Transcriptional Network That Controls Melanogenesis and Melanoma Progression. Cell Death Differ. 2014, 21, 1250–1261. [Google Scholar] [CrossRef]
- Marks, M.S.; Seabra, M.C. The Melanosome: Membrane Dynamics in Black and White. Nat. Rev. Mol. Cell Biol. 2001, 2, 738–748. [Google Scholar] [CrossRef]
- Jimbow, K.; Park, J.S.; Kato, F.; Hirosaki, K.; Toyofuku, K.; Hua, C.; Yamashita, T. Assembly, Target-Signaling and Intracellular Transport of Tyrosinase Gene Family Proteins in the Initial Stage of Melanosome Biogenesis. Pigment Cell Res. 2000, 13, 222–229. [Google Scholar] [CrossRef]
- Park, H.Y.; Kosmadaki, M.; Yaar, M.; Gilchrest, B.A. Cellular Mechanisms Regulating Human Melanogenesis. Cell. Mol. Life Sci. 2009, 66, 1493–1506. [Google Scholar] [CrossRef]
- Schiaffino, M.V. Signaling Pathways in Melanosome Biogenesis and Pathology. Int. J. Biochem. Cell Biol. 2010, 42, 1094–1104. [Google Scholar] [CrossRef]
- Kushimoto, T.; Basrur, V.; Valencia, J.; Matsunaga, J.; Vieira, W.D.; Ferrans, V.J.; Muller, J.; Appella, E.; Hearing, V.J. A Model for Melanosome Biogenesis Based on the Purification and Analysis of Early Melanosomes. Proc. Natl. Acad. Sci. USA 2001, 98, 10698–10703. [Google Scholar] [CrossRef]
- Sitaram, A.; Marks, M.S. Mechanisms of Protein Delivery to Melanosomes in Pigment Cells. Physiology 2012, 27, 85–99. [Google Scholar] [CrossRef]
- Hurbain, I.; Geerts, W.J.C.; Boudier, T.; Marco, S.; Verkleij, A.J.; Marks, M.S.; Raposo, G. Electron Tomography of Early Melanosomes: Implications for Melanogenesis and the Generation of Fibrillar Amyloid Sheets. Proc. Natl. Acad. Sci. USA 2008, 105, 19726–19731. [Google Scholar] [CrossRef]
- Berson, J.F.; Harper, D.C.; Tenza, D.; Raposo, G.; Marks, M.S. Pmel17 Initiates Premelanosome Morphogenesis within Multivesicular Bodies. Mol. Biol. Cell 2001, 12, 3451–3464. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Tenza, D.; Murphy, D.M.; Berson, J.F.; Marks, M.S. Distinct Protein Sorting and Localization to Premelanosomes, Melanosomes, and Lysosomes in Pigmented Melanocytic Cells. J. Cell Biol. 2001, 152, 809–824. [Google Scholar] [CrossRef] [PubMed]
- Bissig, C.; Rochin, L.; van Niel, G. PMEL Amyloid Fibril Formation: The Bright Steps of Pigmentation. Int. J. Mol. Sci. 2016, 17, 1438. [Google Scholar] [CrossRef] [PubMed]
- Giordano, F.; Bonetti, C.; Surace, E.M.; Marigo, V.; Raposo, G. The Ocular Albinism Type 1 (OA1) G-Protein-Coupled Receptor Functions with MART-1 at Early Stages of Melanogenesis to Control Melanosome Identity and Composition. Hum. Mol. Genet. 2009, 18, 4530–4545. [Google Scholar] [CrossRef]
- Lee, Z.H.; Hou, L.; Moellmann, G.; Kuklinska, E.; Antol, K.; Fraser, M.; Halaban, R.; Kwon, B.S. Characterization and Subcellular Localization of Human Pmel 17/Silver, a 100-KDa (Pre)Melanosomal Membrane Protein Associated with 5,6,-Dihydroxyindole-2-Carboxylic Acid (DHICA) Converting Activity. J. Investig. Dermatol. 1996, 106, 605–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimbow, K.; Ishida, O.; Ito, S.; Hori, Y.; Witkop, C.J.; King, R.A. Combined Chemical and Electron Microscopic Studies of Pheomelanosomes in Human Red Hair. J. Investig. Dermatol. 1983, 81, 506–511. [Google Scholar] [CrossRef]
- D’Alba, L.; Shawkey, M.D. Melanosomes: Biogenesis, Properties, and Evolution of an Ancient Organelle. Physiol. Rev. 2019, 99, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Jimbow, K.; Hua, C.; Gomez, P.F.; Hirosaki, K.; Shinoda, K.; Salopek, T.G.; Matsusaka, H.; Jin, H.Y.; Yamashita, T. Intracellular Vesicular Trafficking of Tyrosinase Gene Family Protein in Eu- and Pheomelanosome Biogenesis. Pigment Cell Res. 2000, 13 (Suppl. 8), 110–117. [Google Scholar] [CrossRef] [PubMed]
- Furumura, M.; Sakai, C.; Potterf, S.B.; Vieira, W.D.; Barsh, G.S.; Hearing, V.J. Characterization of Genes Modulated during Pheomelanogenesis Using Differential Display. Proc. Natl. Acad. Sci. USA 1998, 95, 7374–7378. [Google Scholar] [CrossRef]
- Chakraborty, A.K.; Platt, J.T.; Kim, R.K.; Kwon, B.S.; Bennett, D.C.; Pawelek, J.M. Polymerization of 5,6-Dihydroxyindole-2-Carboxylic Acid to Melanin by the Pmel 17/Silver Locus Protein. Eur. J. Biochem. 1996, 236, 180–188. [Google Scholar] [CrossRef]
- Ho, T.; Watt, B.; Spruce, L.A.; Seeholzer, S.H.; Marks, M.S. The Kringle-like Domain Facilitates Post-Endoplasmic Reticulum Changes to Premelanosome Protein (PMEL) Oligomerization and Disulfide Bond Configuration and Promotes Amyloid Formation. J. Biol. Chem. 2016, 291, 3595–3612. [Google Scholar] [CrossRef]
- Maresh, G.A.; Wang, W.C.; Beam, K.S.; Malacko, A.R.; Hellström, I.; Hellström, K.E.; Marquardt, H. Differential Processing and Secretion of the Melanoma-Associated ME20 Antigen. Arch. Biochem. Biophys. 1994, 311, 95–102. [Google Scholar] [CrossRef]
- Berson, J.F.; Theos, A.C.; Harper, D.C.; Tenza, D.; Raposo, G.; Marks, M.S. Proprotein Convertase Cleavage Liberates a Fibrillogenic Fragment of a Resident Glycoprotein to Initiate Melanosome Biogenesis. J. Cell Biol. 2003, 161, 521–533. [Google Scholar] [CrossRef]
- Leonhardt, R.M.; Vigneron, N.; Rahner, C.; Cresswell, P. Proprotein Convertases Process Pmel17 during Secretion. J. Biol. Chem. 2011, 286, 9321–9337. [Google Scholar] [CrossRef]
- Rochin, L.; Hurbain, I.; Serneels, L.; Fort, C.; Watt, B.; Leblanc, P.; Marks, M.S.; De Strooper, B.; Raposo, G.; Van Niel, G. BACE2 Processes PMEL to Form the Melanosome Amyloid Matrix in Pigment Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 10658–10663. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, M.; Hozumi, Y.; Suzuki, T. ADAM Protease Inhibitors Reduce Melanogenesis by Regulating PMEL17 Processing in Human Melanocytes. J. Dermatol. Sci. 2015, 78, 133–142. [Google Scholar] [CrossRef]
- Theos, A.C.; Berson, J.F.; Theos, S.C.; Herman, K.E.; Harper, D.C.; Tenza, D.; Sviderskaya, E.V.; Lamoreux, M.L.; Bennett, D.C.; Raposo, G.; et al. Dual Loss of ER Export and Endocytic Signals with Altered Melanosome Morphology in the Silver Mutation of Pmel17. Mol. Biol. Cell 2006, 17, 3598–3612. [Google Scholar] [CrossRef]
- van Niel, G.; Charrin, S.; Simoes, S.; Romao, M.; Rochin, L.; Saftig, P.; Marks, M.S.; Rubinstein, E.; Raposo, G. The Tetraspanin CD63 Regulates ESCRT-Independent and -Dependent Endosomal Sorting during Melanogenesis. Dev. Cell 2011, 21, 708–721. [Google Scholar] [CrossRef]
- van Niel, G.; Bergam, P.; Di Cicco, A.; Hurbain, I.; Lo Cicero, A.; Dingli, F.; Palmulli, R.; Fort, C.; Potier, M.C.; Schurgers, L.J.; et al. Apolipoprotein E Regulates Amyloid Formation within Endosomes of Pigment Cells. Cell Rep. 2015, 13, 43–51. [Google Scholar] [CrossRef]
- Hoashi, T.; Watabe, H.; Muller, J.; Yamaguchi, Y.; Vieira, W.D.; Hearing, V.J. MART-1 Is Required for the Function of the Melanosomal Matrix Protein PMEL17/GP100 and the Maturation of Melanosomes. J. Biol. Chem. 2005, 280, 14006–14016. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, M.V.; Baschirotto, C.; Pellegrini, G.; Montalti, S.; Tacchetti, C.; De Luca, M.; Ballabio, A. The Ocular Albinism Type 1 Gene Product Is a Membrane Glycoprotein Localized to Melanosomes. Proc. Natl. Acad. Sci. USA 1996, 93, 9055–9060. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, T.; Jolly, R.; Martin-Martin, B.; Seabra, M.C.; Piccirillo, R.; Schiaffino, M.V.; Futter, C.E. Expression of OA1 Limits the Fusion of a Subset of MVBs with Lysosomes—A Mechanism Potentially Involved in the Initial Biogenesis of Melanosomes. J. Cell Sci. 2014, 127, 700. [Google Scholar] [CrossRef]
- Falletta, P.; Bagnato, P.; Bono, M.; Monticone, M.; Schiaffino, M.V.; Bennett, D.C.; Goding, C.R.; Tacchetti, C.; Valetti, C. Melanosome-Autonomous Regulation of Size and Number: The OA1 Receptor Sustains PMEL Expression. Pigment Cell Melanoma Res. 2014, 27, 565–579. [Google Scholar] [CrossRef] [PubMed]
- Maass, P.; Bunde, A.; Ingram, M.D. Ion Transport in Pigmentation. Arch. Biochem. Biophys. 2014, 563, 35–41. [Google Scholar] [CrossRef]
- Liu, T.F.; Kandala, G.; Setaluri, V. PDZ Domain Protein GIPC Interacts with the Cytoplasmic Tail of Melanosomal Membrane Protein Gp75 (Tyrosinase-Related Protein-1). J. Biol. Chem. 2001, 276, 35768–35777. [Google Scholar] [CrossRef] [Green Version]
- Kedlaya, R.; Kandala, G.; Liu, T.F.; Maddodi, N.; Devi, S.; Setaluri, V. Interactions between GIPC-APPL and GIPC-TRP1 Regulate Melanosomal Protein Trafficking and Melanogenesis in Human Melanocytes. Arch. Biochem. Biophys. 2011, 508, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Salas-Cortes, L.; Ye, F.; Tenza, D.; Wilhelm, C.; Theos, A.; Louvard, D.; Raposo, G.; Coudrier, E. Myosin Lb Modulates the Morphology and the Protein Transport within Multi-Vesicular Sorting Endosomes. J. Cell Sci. 2005, 118, 4823–4832. [Google Scholar] [CrossRef] [PubMed]
- Loubéry, S.; Delevoye, C.; Louvard, D.; Raposo, G.; Coudrier, E. Myosin VI Regulates Actin Dynamics and Melanosome Biogenesis. Traffic 2012, 13, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Delevoye, C.; Hurbain, I.; Tenza, D.; Sibarita, J.B.; Uzan-Gafsou, S.; Ohno, H.; Geerts, W.J.C.; Verkleij, A.J.; Salamero, J.; Marks, M.S.; et al. AP-1 and KIF13A Coordinate Endosomal Sorting and Positioning during Melanosome Biogenesis. J. Cell Biol. 2009, 187, 247–264. [Google Scholar] [CrossRef]
- Theos, A.C.; Martina, A.; Hurbain, I.; Peden, A.A.; Sviderskaya, E.V.; Stewart, A.; Robinson, M.S.; Bennett, D.C.; Cutler, D.F.; Bonifacino, J.S.; et al. Functions of Adaptor Protein (AP)-3 and AP-1 in Tyrosinase Sorting from Endosomes to Melanosomes. Mol. Biol. Cell 2005, 16, 5356–5372. [Google Scholar] [CrossRef] [PubMed]
- Huizing, M.; Sarangarajan, R.; Strovel, E.; Zhao, Y.; Gahl, W.A.; Boissy, R.E. AP-3 Mediates Tyrosinase but Not TRP-1 Trafficking in Human Melanocytes. Mol. Biol. Cell 2001, 12, 2075–2085. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, K.; Valencia, J.C.; Kushimoto, T.; Costin, G.E.; Virador, V.M.; Vieira, W.D.; Ferrans, V.J.; Hearing, V.J. The Etiology of Oculocutaneous Albinism (OCA) Type II: The Pink Protein Modulates the Processing and Transport of Tyrosinase. Pigment Cell Res. 2002, 15, 217–224. [Google Scholar] [CrossRef]
- Potterf, S.B.; Furumura, M.; Sviderskaya, E.V.; Santis, C.; Bennett, D.C.; Hearing, V.J. Normal Tyrosine Transport and Abnormal Tyrosinase Routing in Pink-Eyed Dilution Melanocytes. Exp. Cell Res. 1998, 244, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, D.J.; Rodriguez-Fernandez, I.A.; Dell’Angelica, E.C. Functional Interactions between OCA2 and the Protein Complexes BLOC-1, BLOC-2, and AP-3 Inferred from Epistatic Analyses of Mouse Coat Pigmentation. Pigment Cell Melanoma Res. 2011, 24, 275–281. [Google Scholar] [CrossRef]
- Chen, K.; Manga, P.; Orlow, S.J. Pink-Eyed Dilution Protein Controls the Processing of Tyrosinase. Mol. Biol. Cell 2002, 13, 1953–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manga, P.; Pifko-Hirst, S.; Zhou, B.K.; Orlow, S.J.; Boissy, R.E. Mislocalization of Melanosomal Proteins in Melanocytes from Mice with Oculocutaneous Albinism Type 2. Exp. Eye Res. 2001, 72, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Sitaram, A.; Piccirillo, R.; Palmisano, I.; Harper, D.C.; Dell’Angelica, E.C.; Schiaffino, M.V.; Marks, M.S. Localization to Mature Melanosomes by Virtue of Cytoplasmic Dileucine Motifs Is Required for Human OCA2 Function. Mol. Biol. Cell 2009, 20, 1464–1477. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, S.; Jaume, A.; Jani, R.A.; Delevoye, C.; Raposo, G.; Marks, M.S. Type II Phosphatidylinositol 4-Kinases Function Sequentially in Cargo Delivery from Early Endosomes to Melanosomes. J. Cell Biol. 2022, 221, e202110114. [Google Scholar] [CrossRef]
- Petris, M.J. The Menkes Copper Transporter Is Required for the Activation of Tyrosinase. Hum. Mol. Genet. 2000, 9, 2845–2851. [Google Scholar] [CrossRef]
- Rao, S.; Setty, G.; Tenza, D.; Sviderskaya, E.V.; Bennett, D.C.; Raposo, G.; Marks, M.S. Cell-Specific ATP7A Transport Sustains Copper-Dependent Tyrosinase Activity in Melanosomes. Nature 2008, 454, 1142–1146. [Google Scholar]
- Truschel, S.T.; Simoes, S.; Rao, S.; Setty, G.; Harper, D.C.; Tenza, D.; Thomas, P.C.; Herman, K.E.; Sackett, S.D.; David, C.; et al. ESCRT-I Function Is Required for Tyrp1 Transport from Early Endosomes to the Melanosome Limiting Membrane. Traffic 2010, 10, 1318–1336. [Google Scholar] [CrossRef] [PubMed]
- Yatsu, A.; Ohbayashi, N.; Tamura, K.; Fukuda, M. Syntaxin-3 Is Required for Melanosomal Localization of Tyrp1 in Melanocytes. J. Investig. Dermatol. 2013, 133, 2237–2246. [Google Scholar] [CrossRef]
- Jani, R.A.; Purushothaman, L.K.; Rani, S.; Bergam, P.; Curie, I.; Recherche, C. De STX13 Regulates Cargo Delivery from Recycling Endosomes during Melanosome Biogenesis Accepted Manuscript Journal of Cell Science Accepted Manuscript. J. Cell Sci. 2015, 128, 3263–3276. [Google Scholar]
- Fukuda, M. Rab GTPases: Key Players in Melanosome Biogenesis, Transport, and Transfer. Pigment Cell Melanoma Res. 2021, 34, 222–235. [Google Scholar] [CrossRef]
- Tamura, K.; Ohbayashi, N.; Maruta, Y.; Kanno, E.; Itoh, T.; Fukuda, M. Varp Is a Novel Rab32/38-Binding Protein That Regulates Tyrp1 Trafficking in Melanocytes. Mol. Biol. Cell 2009, 20, 2900–2908. [Google Scholar] [CrossRef]
- Wasmeier, C.; Romao, M.; Plowright, L.; Bennett, D.C.; Raposo, G.; Seabra, M.C. Rab38 and Rab32 Control Post-Golgi Trafficking of Melanogenic Enzymes. J. Cell Biol. 2006, 175, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.K.; Delevoye, C.; Acosta-Ruiz, A.; Hurbain, I.; Romao, M.; Hesketh, G.G.; Goff, P.S.; Sviderskaya, E.V.; Bennett, D.C.; Luzio, J.P.; et al. BLOC-1 and BLOC-3 Regulate VAMP7 Cycling to and from Melanosomes via Distinct Tubular Transport Carriers. J. Cell Biol. 2016, 214, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Hirosaki, K.; Yamashita, T.; Wada, I.; Jin, H.Y.; Jimbow, K. Tyrosinase and Tyrosinase-Related Protein 1 Require Rab7 for Their Intracellular Transport. J. Investig. Dermatol. 2002, 119, 475–480. [Google Scholar] [CrossRef]
- Hida, T.; Sohma, H.; Kokai, Y.; Kawakami, A.; Hirosaki, K.; Okura, M.; Tosa, N.; Yamashita, T.; Jimbow, K. Rab7 Is a Critical Mediator in Vesicular Transport of Tyrosinase-Related Protein 1 in Melanocytes. J. Dermatol. 2011, 38, 432–441. [Google Scholar] [CrossRef]
- Gomez, P.F.; Luo, D.; Hirosaki, K.; Shinoda, K.; Yamashita, T.; Suzuki, J.I.; Otsu, K.; Ishikawa, K.; Jimbow, K. Identification of Rab7 as a Melanosome-Associated Protein Involved in the Intracellular Transport of Tyrosinase-Related Protein 1. J. Investig. Dermatol. 2001, 117, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Sakane, F.; Imai, S.I.; Yasuda, S.; Kai, M.; Kanoh, H.; Jin, H.Y.; Hirosaki, K.; Yamashita, T.; Fisher, D.E.; et al. Rab7 Regulates Maturation of Melanosomal Matrix Protein Gp100/Pmel17/Silv. J. Investig. Dermatol. 2008, 128, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Marubashi, S.; Shimada, H.; Fukuda, M.; Ohbayashi, N. RUTBC1 Functions as a GTPase-Activating Protein for Rab32/38 and Regulates Melanogenic Enzyme Trafficking in Melanocytes. J. Biol. Chem. 2016, 291, 1427–1440. [Google Scholar] [CrossRef]
- Stenmark, H. Rab GTPases as Coordinators of Vesicle Traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef]
- Le, L.; Sirés-Campos, J.; Raposo, G.; Delevoye, C.; Marks, M.S. Melanosome Biogenesis in the Pigmentation of Mammalian Skin. Integr. Comp. Biol. 2021, 61, 1517–1545. [Google Scholar] [CrossRef]
- Simon, J.D.; Peles, D.N. The Red and the Black. Acc. Chem. Res. 2010, 43, 1452–1460. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Wakamatsu, K. Quantitative Analysis of Eumelanin and Pheomelanin in Humans, Mice, and Other Animals: A Comparative Review. Pigment Cell Res. 2003, 16, 523–531. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Chemistry of Mixed Melanogenesis—Pivotal Roles of Dopaquinone. Photochem. Photobiol. 2008, 84, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.D.; Peles, D.; Wakamatsu, K.; Ito, S. Current Challenges in Understanding Melanogenesis: Bridging Chemistry, Biological Control, Morphology, and Function. Pigment Cell Melanoma Res. 2009, 22, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Schallreuter, K.; Slominski, A.; Jm, P.; Jimbow, K.; Ba, G.; Pawelek, J.M.; Gilchrest, B.A. What Controls Melanogenesis ? Exp. Dermatol. 1998, 7, 143–150. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Schulz-Douglas, V.; Bünz, A.; Beazley, W.; Körner, C. Pteridines in the Control of Pigmentation. J. Investig. Dermatol. 1997, 109, 31–35. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Wood, J.M.; Pittelkow, M.R.; Gütlich, M.; Lemke, K.R.; Rödl, W.; Swanson, N.N.; Hitzemann, K.; Ziegler, I. Regulation of Melanin Biosynthesis in the Human Epidermis by Tetrahydrobiopterin. Science 1994, 263, 1444–1446. [Google Scholar] [CrossRef]
- Harada, T.; Kagamiyama, H.; Hatakeyama, K. Feedback Regulation Mechanisms for the Control of GTP Cyclohydrolase I Activity. Science 1993, 260, 1507–1510. [Google Scholar] [CrossRef]
- Wood, J.M.; Schallreuter-Wood, K.U.; Lindsey, N.J.; Callaghan, S.; Gardner, M.L.G. A Specific Tetrahydrobiopterin Binding Domain on Tyrosinase Controls Melanogenesis. Biochem. Biophys. Res. Commun. 1995, 206, 480–485. [Google Scholar] [CrossRef]
- García-Molina, F.; Munoz-Munoz, J.L.; Acosta, J.R.; García-Ruiz, P.A.; Tudela, J.; García-Cánovas, F.; Rodríguez-López, J.N. Melanogenesis Inhibition by Tetrahydropterines. Biochim. Biophys. Acta Proteins Proteom. 2009, 1794, 1766–1774. [Google Scholar] [CrossRef]
- Schallreuter-Wood, K.U.; Wood, J.M. Control of Melanogenesis in the Human Epidermis by the Redox-Status of Tetrahydrobiopterins. Pteridines 1995, 6, 101–103. [Google Scholar] [CrossRef]
- Gaudel, C.; Soysouvanh, F.; Leclerc, J.; Bille, K.; Husser, C.; Montcriol, F.; Bertolotto, C.; Ballotti, R. Regulation of Melanogenesis by the Amino Acid Transporter SLC7A5. J. Investig. Dermatol. 2020, 140, 2253–2259.e4. [Google Scholar] [CrossRef]
- Hearing, V.J. Determination of Melanin Synthetic Pathways. J. Investig. Dermatol. 2011, 131, E8–E11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, S.; Suzuki, N.; Takebayashi, S.; Commo, S.; Wakamatsu, K. Neutral PH and Copper Ions Promote Eumelanogenesis after the Dopachrome Stage. Pigment Cell Melanoma Res. 2013, 26, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Land, E.J.; Riley, P.A. Spontaneous Redox Reactions of Dopaquinone and the Balance between the Eumelanic and Phaeomelanic Pathways. Pigment Cell Res 2000, 13, 273–277. [Google Scholar] [CrossRef]
- Wakamatsu, K.; Ohtara, K.; Ito, S. Chemical Analysis of Late Stages of Pheomelanogenesis: Conversion of Dihydrobenzothiazine to a Benzothiazole Structure. Pigment Cell Melanoma Res. 2009, 22, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Benathan, M.; Labidi, F. Cysteine-Dependent 5-S-Cysteinyldopa Formation and Its Regulation by Glutathione in Normal Epidermal Melanocytes. Arch. Dermatol. Res. 1996, 288, 697–702. [Google Scholar] [CrossRef]
- Land, E.J.; Ito, S.; Wakamatsu, K.; Riley, P.A. Rate Constants for the First Two Chemical Steps of Eumelanogenesis. Pigment Cell Res. 2003, 16, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Aroca, P.; Solano, F.; Salina, C.; García-Borrón, J.C.; Lozano, J.A. Regulation of the Final Phase of Mammalian Melanogenesis: The Role of Dopachrome Tautomerase and the Ratio between 5,6-dihydroxyindole-2-carboxylic Acid and 5,6-dihydroxyindole. Eur. J. Biochem. 1992, 208, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Kroumpouzos, G.; Urabe, K.; Kobayashi, T.; Sakai, C.; Hearing, V.J. Functional Analysis of the Slaty Gene Product (TRP2) as Dopachrome Tautomerase and the Effect of a Point Mutation on Its Catalytic Function. Biochem. Biophys. Res. Commun. 1994, 202, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V.J. A Second Tyrosinase-Related Protein, TRP-2, Is a Melanogenic Enzyme Termed DOPAchrome Tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [CrossRef]
- Commo, P.; Gaillard, O.; Thibaut, S.; Bernard, B.A. Absence of TRP-2 in Melanogenic Melanocytes of Human Hair. Pigment Cell Res. 2004, 17, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Ozeki, H.; Ito, S.; Wakamatsu, K. Chemical Characterization of Melanins in Sheep Wool and Human Hair. Pigment Cell Res. 1996, 9, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Körner, A.; Pawelek, J. Mammalian Tyrosinase Catalyzes Three Reactions in the Biosynthesis of Melanin. Science 1982, 217, 1163–1165. [Google Scholar] [CrossRef]
- Hearing, V.J. Biochemical Control of Melanogenesis and Melanosomal Organization. J. Investig. Dermatol. Symp. Proc. 1999, 4, 24–28. [Google Scholar] [CrossRef]
- Zhao, H.; Eling, D.J.; Medrano, E.E.; Boissy, R.E. Retroviral Infection with Human Tyrosinase-Related Protein-1 (TRP-1) CDNA Upregulates Tyrosinase Activity and Melanin Synthesis in a TRP-1-Deficient Melanoma Cell Line. J. Investig. Dermatol. 1996, 106, 744–752. [Google Scholar] [CrossRef]
- Martinez-Esparza, M.; Jiménez-Cervantes, C.; García-Borrón, J.C.; Lozano, J.A.; Del Marmol, V.; Ghanem, G.; Solano, F. Comparison of TRPs from Murine and Human Malignant Melanocytes. Pigment Cell Res. 1997, 10, 229–235. [Google Scholar] [CrossRef]
- Jimenez-Cervantes, C.; Solano, F.; Kobayashi, T.; Urabe, K.; Hearing, V.J.; Lozano, J.A.; Garcia-Borron, J.C. A New Enzymatic Function in the Melanogenic Pathway. The 5,6- Dihydroxyindole-2-Carboxylic Acid Oxidase Activity of Tyrosinase-Related Protein-1 (TRP1). J. Biol. Chem. 1994, 269, 17993–18000. [Google Scholar] [CrossRef]
- Dolinska, M.B.; Wingfield, P.T.; Young, K.L.; Sergeev, Y.V. The TYRP1-Mediated Protection of Human Tyrosinase Activity Does Not Involve Stable Interactions of Tyrosinase Domains. Pigment Cell Melanoma Res. 2019, 32, 753–765. [Google Scholar] [CrossRef]
- Boissy, R.E.; Sakai, C.; Zhao, H.; Kobayashi, T.; Hearing, V.J. Human Tyrosinase Related Protein-1 (TRP-1) Does Not Function as a DHICA Oxidase Activity in Contrast to Murine TRP-1. Exp. Dermatol. 1998, 7, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Olivares, C.; Jiménez-Cervantes, C.; Lozano, J.A.; Solano, F.; García-Borrón, J.C. The 5,6-Dihydroxyindole-2-Carboxylic Acid (DHICA) Oxidase Activity of Human Tyrosinase. Biochem. J. 2001, 354 Pt 1, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hebert, D.N. Tyrosinase Maturation through the Mammalian Secretory Pathway: Bringing Color to Life. Pigment Cell Res. 2006, 19, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Wichers, H.J.; Soler-Lopez, M.; Dijkstra, B.W. Structure and Function of Human Tyrosinase and Tyrosinase-Related Proteins. Chem.—A Eur. J. 2018, 24, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Moellmann, G.; Kuklinska, E.; Bomirski, A.; Pawelek, J. Positive Regulation of Melanin Pigmentation by Two Key Substrates of the Melanogenic Pathway, L-Tyrosine and L-Dopa. J. Cell Sci. 1988, 89, 287–296. [Google Scholar] [CrossRef]
- Wilczek, A.; Mishima, Y. Inhibitory Effects of Melanin Monomers, Dihydroxyindole-2-Carboxylic Acid (DHICA) and Dihydroxyindole (DHI) on Mammalian Tyrosinase, With a Special Reference to the Role of DHICA/DHI Ratio in Melanogenesis. Pigment Cell Res. 1995, 8, 105–112. [Google Scholar] [CrossRef]
- Kobayashi, T.; Imokawa, G.; Bennett, D.C.; Hearing, V.J. Tyrosinase Stabilization by Tyrp1 (the Brown Locus Protein). J. Biol. Chem. 1999, 273, 31801–31805. [Google Scholar] [CrossRef]
- Manga, P.; Sato, K.; Ye, L.; Beermann, F.; Lynn Lamoreux, M.; Orlow, S.J. Mutational Analysis of the Modulation of Tyrosinase by Tyrosinase-Related Proteins 1 and 2 in Vitro. Pigment Cell Res. 2000, 13, 364–374. [Google Scholar] [CrossRef]
- Chintala, S.; Li, W.; Lamoreux, M.L.; Ito, S.; Wakamatsu, K.; Sviderskaya, E.V.; Bennett, D.C.; Park, Y.; Gahl, W.A.; Huizing, M.; et al. Slc7a11 Gene Controls Production of Pheomelanin Pigment and Proliferation of Cultured Cells. Proc. Natl. Acad. Sci. USA 2005, 102, 10964–10969. [Google Scholar] [CrossRef]
- Yang, N.; Mu, L.I.N.; Zhao, B.; Wang, M.; Hu, S.; Zhao, B.I.N. RNAi-Mediated SLC7A11 Knockdown Inhibits Melanogenesis-Related Genes Expression in Rabbit Skin Fibroblasts. J. Genet. 2018, 97, 463–468. [Google Scholar] [CrossRef]
- Chiaverini, C.; Sillard, L.; Flori, E.; Ito, S.; Briganti, S.; Wakamatsu, K.; Fontas, E.; Berard, E.; Cailliez, M.; Cochat, P.; et al. Cystinosin Is a Melanosomal Protein That Regulates Melanin Synthesis. FASEB J. 2012, 26, 3779–3789. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.N.; Lee, J.C. PH-Dependent Fibril Maturation of a Pmel17 Repeat Domain Isoform Revealed by Tryptophan Fluorescence. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Ancans, J.; Tobin, D.J.; Hoogduijn, M.J.; Smit, N.P.; Wakamatsu, K.; Thody, A.J. Melanosomal PH Controls Rate of Melanogenesis, Eumelanin/Phaeomelanin Ratio and Melanosome Maturation in Melanocytes and Melanoma Cells. Exp. Cell Res. 2001, 268, 26–35. [Google Scholar] [CrossRef]
- Tabata, H.; Kawamura, N.; Sun-Wada, G.H.; Wada, Y. Vacuolar-Type H+-ATPase with the A3 Isoform Is the Proton Pump on Premature Melanosomes. Cell Tissue Res. 2008, 332, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, K.; Nagao, A.; Watanabe, M.; Nakao, K.; Ito, S. Pheomelanogenesis Is Promoted at a Weakly Acidic PH. Pigment Cell Melanoma Res. 2017, 30, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Ota, K.; Nardin, C.; Feldman, M.; Widman, A.; Wind, O.; Simon, A.; Reilly, M.; Levin, L.R.; Buck, J.; et al. Mammalian Pigmentation Is Regulated by a Distinct CAMP-Dependent Mechanism That Controls Melanosome PH. Sci. Signal 2018, 11, eaau7987. [Google Scholar] [CrossRef]
- Dooley, C.M.; Schwarz, H.; Mueller, K.P.; Mongera, A.; Konantz, M.; Neuhauss, S.C.F.; Nüsslein-Volhard, C.; Geisler, R. Slc45a2 and V-ATPase Are Regulators of Melanosomal PH Homeostasis in Zebrafish, Providing a Mechanism for Human Pigment Evolution and Disease. Pigment Cell Melanoma Res. 2013, 26, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Ancans, J.; Thody, A.J. Activation of Melanogenesis by Vacuolar Type H+-ATPase Inhibitors in Amelanotic, Tyrosinase Positive Human and Mouse Melanoma Cells. FEBS Lett. 2000, 478, 57–60. [Google Scholar] [CrossRef]
- Smith, D.R.; Spaulding, D.T.; Glenn, H.M.; Fuller, B.B. The Relationship between Na+/H+ Exchanger Expression and Tyrosinase Activity in Human Melanocytes. Exp. Cell Res. 2004, 298, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Puri, N.; Gardner, J.M.; Brilliant, M.H. Aberrant PH of Melanosomes in Pink-Eyed Dilution (p) Mutant Melanocytes. J. Investig. Dermatol. 2000, 115, 607–613. [Google Scholar] [CrossRef]
- Brilliant, M.H. The Mouse p (Pink-Eyed Dilution) and Human P Genes, Oculocutaneous Albinism Type 2 (OCA2), and Melanosomal PH. Pigment Cell Res. 2001, 14, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Bellono, N.W.; Escobar, I.E.; Lefkovith, A.J.; Marks, M.S.; Oancea, E. An Intracellular Anion Channel Critical for Pigmentation. Elife 2014, 3, e04543. [Google Scholar] [CrossRef] [PubMed]
- Weinert, S.; Jabs, S.; Hohensee, S.; Chan, W.L.; Kornak, U.; Jentsch, T.J. Transport Activity and Presence of ClC-7/Ostm1 Complex Account for Different Cellular Functions. EMBO Rep. 2014, 15, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Koroma, D.C.; Scales, J.L.; Trotman, J.C.; Wakamatsu, K.; Ito, S.; Oancea, E. The Lysosomal Chloride-Proton Exchanger CLC7 Functions in Melanosomes as a Negative Regulator of Human Pigmentation. bioRxiv 2021, 2021.02.05.430016. [Google Scholar] [CrossRef]
- Han, H.Y.; Lee, J.R.; Xu, W.A.; Hahn, M.J.; Yang, J.M.; Park, Y.D. Effect of Cl−on Tyrosinase: Complex Inhibition Kinetics and Biochemical Implication. J. Biomol. Struct. Dyn. 2007, 25, 165–171. [Google Scholar] [CrossRef]
- Bin, B.H.; Bhin, J.; Yang, S.H.; Shin, M.; Nam, Y.J.; Choi, D.H.; Shin, D.W.; Lee, A.Y.; Hwang, D.; Cho, E.G.; et al. Membrane-Associated Transporter Protein (MATP) Regulates Melanosomal PH and Influences Tyrosinase Activity. PLoS ONE 2015, 10, e0129273. [Google Scholar] [CrossRef]
- Costin, G.E.; Valencia, J.C.; Vieira, W.D.; Lamoreux, M.L.; Hearing, V.J. Tyrosinase Processing and Intracellular Trafficking Is Disrupted in Mouse Primary Melanocytes Carrying the Underwhite (Uw) Mutation. A Model for Oculocutaneous Albinism (OCA) Type 4. J. Cell Sci. 2003, 116 Pt 15, 3203–3212. [Google Scholar] [CrossRef]
- Ginger, R.S.; Askew, S.E.; Ogborne, R.M.; Wilson, S.; Ferdinando, D.; Dadd, T.; Smith, A.M.; Kazi, S.; Szerencsei, R.T.; Winkfein, R.J.; et al. SLC24A5 Encodes a Trans -Golgi Network Protein with Potassium-Dependent Sodium-Calcium Exchange Activity That Regulates Human Epidermal Melanogenesis. J. Biol. Chem. 2008, 283, 5486–5495. [Google Scholar] [CrossRef]
- Lamason, R.L.; Mohideen, M.; Mest, J.; Wong, A.; Norton, H.; Aros, M.; Jurynec, M.; Mao, X.; Humphreville, V.; Humbert, J.; et al. SLC24A5, a Putative Cation Exchanger, Affects Pigmentation in Zebrafish and Humans. Science 2005, 310, 1782–1786. [Google Scholar] [CrossRef]
- Bush, W.D.; Simon, J.D. Quantification of Ca2+ Binding to Melanin Supports the Hypothesis That Melanosomes Serve a Functional Role in Regulating Calcium Homeostasis. Pigment Cell Res. 2007, 20, 134–139. [Google Scholar] [CrossRef]
- Hoogduijn, M.J.; Smit, N.P.; Van Der Laarse, A.; Van Nieuwpoort, A.F.; Wood, J.M.; Thody, A.J. Melanin Has a Role in Ca2+ Homeostasis in Human Melanocytes. Pigment Cell Res. 2003, 16, 127–132. [Google Scholar] [CrossRef]
- Daniele, T.; Hurbain, I.; Vago, R.; Casari, G.; Raposo, G.; Tacchetti, C.; Schiaffino, M.V. Mitochondria and Melanosomes Establish Physical Contacts Modulated by Mfn2 and Involved in Organelle Biogenesis. Curr. Biol. 2014, 24, 393–403. [Google Scholar] [CrossRef]
- Tanwar, J.; Saurav, S.; Basu, R.; Singh, J.B.; Priya, A.; Dutta, M.; Santhanam, U.; Joshi, M.; Madison, S.; Singh, A.; et al. Mitofusin-2 Negatively Regulates Melanogenesis by Modulating Mitochondrial ROS Generation. Cells 2022, 11, 701. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-Y. The Receptor for Activated C-Kinase-I (RACK-I) Anchors Activated PKC- on Melanosomes. J. Cell Sci. 2004, 117, 3659–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.Y.; Russakovsky, V.; Ohno, S.; Gilchrest, B.A. The β Isoform of Protein Kinase C Stimulates Human Melanogenesis by Activating Tyrosinase in Pigment Cells. J. Biol. Chem. 1993, 268, 11742–11749. [Google Scholar] [CrossRef]
- Wu, H.; Park, H.Y. Protein Kinase C-β-Mediated Complex Formation between Tyrosinase and TRP-1. Biochem. Biophys. Res. Commun. 2003, 311, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Schallreuter, K.U.; Wood, J.M. The Importance of L -Phenylalanine Transport and Its Autocrine Turnover to L -Tyrosine for Melanogenesis in Human Epidermal Melanocytes. Biochem. Biophys. Res. Commun. 1999, 262, 423–428. [Google Scholar] [CrossRef]
- Noh, T.K.; Bang, S.H.; Lee, Y.J.; Cho, H.I.; Jung, M.Y.; Kim, I.; Leem, C.H.; Chang, S.E. The Ion Channel Activator CyPPA Inhibits Melanogenesis via the GSK3β/β-Catenin Pathway. Chem. Biol. Interact. 2019, 300, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.G.; Nair, N.; Begum, G.; Joshi, N.B.; Sinkar, V.P.; Vora, S. Melanocyte-Keratinocyte Interaction Induces Calcium Signalling and Melanin Transfer to Keratinocytes. Pigment Cell Res. 2007, 20, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Oancea, E.; Vriens, J.; Brauchi, S.; Jun, J.; Splawski, I.; Clapham, D.E. TRPM1 Forms Ion Channels Associated with Melanin Content in Melanocytes. Sci. Signal 2009, 2, ra21. [Google Scholar] [CrossRef]
- Lu, S.; Slominski, A.; Yang, S.E.; Sheehan, C.; Ross, J.; Carlson, J.A. The Correlation of TRPM1 (Melastatin) MRNA Expression with Microphthalmia-Associated Transcription Factor (MITF) and Other Melanogenesis-Related Proteins in Normal and Pathological Skin, Hair Follicles and Melanocytic Nevi. J. Cutan. Pathol. 2010, 37, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.; Markandeya, Y.; Maddodi, N.; Dhingra, A.; Vardi, N.; Balijepalli, R.C.; Setaluri, V. Metabotropic Glutamate Receptor 6 Signaling Enhances TRPM1 Calcium Channel Function and Increases Melanin Content in Human Melanocytes. Pigment Cell Melanoma Res. 2013, 26, 348–356. [Google Scholar] [CrossRef]
- Xu, H.; Delling, M.; Li, L.; Dong, X.; Clapham, D.E. Activating Mutation in a Mucolipin Transient Receptor Potential Channel Leads to Melanocyte Loss in Varitint-Waddler Mice. Proc. Natl. Acad. Sci. USA 2007, 104, 18321–18326. [Google Scholar] [CrossRef]
- Stanisz, H.; Stark, A.; Kilch, T.; Schwarz, E.C.; Müller, C.S.L.; Peinelt, C.; Hoth, M.; Niemeyer, B.A.; Vogt, T.; Bogeski, I. ORAI1 Ca2+channels Control Endothelin-1-Induced Mitogenesis and Melanogenesis in Primary Human Melanocytes. J. Investig. Dermatol. 2012, 132, 1443–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calcraft, P.J.; Ruas, M.; Pan, Z.; Cheng, X.; Arredouani, A.; Hao, X.; Tang, J.; Rietdorf, K.; Teboul, L.; Chuang, K.T.; et al. NAADP Mobilizes Calcium from Acidic Organelles through Two-Pore Channels. Nature 2009, 459, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, X.; Dong, X.P.; Samie, M.; Li, X.; Cheng, X.; Goschka, A.; Shen, D.; Zhou, Y.; Harlow, J.; et al. TPC Proteins Are Phosphoinositide- Activated Sodium-Selective Ion Channels in Endosomes and Lysosomes. Cell 2012, 151, 372–383. [Google Scholar] [CrossRef]
- Bellono, N.W.; Escobar, I.E.; Oancea, E. A Melanosomal Two-Pore Sodium Channel Regulates Pigmentation. Sci. Rep. 2016, 6, 26570. [Google Scholar] [CrossRef]
- Sirés-Campos, J.; Lambertos, A.; Delevoye, C.; Raposo, G.; Bennett, D.C.; Sviderskaya, E.; Jiménez-Cervantes, C.; Olivares, C.; García-Borrón, J.C. Mahogunin Ring Finger 1 Regulates Pigmentation by Controlling the PH of Melanosomes in Melanocytes and Melanoma Cells. Cell. Mol. Life Sci. 2021, 79, 47. [Google Scholar] [CrossRef]
- Hume, A.N.; Seabra, M.C. Melanosomes on the Move: A Model to Understand Organelle Dynamics. Biochem. Soc. Trans. 2011, 39, 1191–1196. [Google Scholar] [CrossRef]
- Hara, M.; Yaar, M.; Byers, H.R.; Goukassian, D.; Fine, R.E.; Gonsalves, J.; Gilchrest, B.A. Kinesin Participates in Melanosomal Movement along Melanocyte Dendrites. J. Investig. Dermatol. 2000, 114, 438–443. [Google Scholar] [CrossRef]
- Byers, H.R.; Yaar, M.; Eller, M.S.; Jalbert, N.L.; Gilchrest, B.A. Role of Cytoplasmic Dynein in Melanosome Transport in Human Melanocytes. J. Investig. Dermatol. 2000, 114, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Vancoillie, G.; Lambert, J.; Mulder, A.; Koerten, H.K.; Mommaas, A.M.; Van Oostveldt, P.; Naeyaert, J.M. Kinesin and Kinectin Can Associate with the Melanosomal Surface and Form a Link with Microtubules in Normal Human Melanocytes. J. Investig. Dermatol. 2000, 114, 421–429. [Google Scholar] [CrossRef]
- Ishida, M.; Ohbayashi, N.; Fukuda, M. Rab1A Regulates Anterograde Melanosome Transport by Recruiting Kinesin-1 to Melanosomes through Interaction with SKIP. Sci. Rep. 2015, 5, 8238. [Google Scholar] [CrossRef]
- Ohbayashi, N.; Maruta, Y.; Ishida, M.; Fukuda, M. Melanoregulin Regulates Retrograde Melanosome Transport through Interaction with the RILP-P150Glued Complex in Melanocytes. J. Cell Sci. 2012, 125 Pt 6, 1508–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, T.; Ohbayashi, N.; Fukuda, M. The Rab Interacting Lysosomal Protein (RILP) Homology Domain Functions as a Novel Effector Domain for Small GTPase Rab36: Rab36 Regulates Retrograde Melanosome Transport in Melanocytes. J. Biol. Chem. 2012, 287, 28619–28631. [Google Scholar] [CrossRef] [PubMed]
- Chiaverini, C.; Beuret, L.; Flori, E.; Busca, R.; Abbe, P.; Bille, K.; Bahadoran, P.; Ortonne, J.P.; Bertolotto, C.; Ballotti, R. Microphthalmia-Associated Transcription Factor Regulates RAB27A Gene Expression and Controls Melanosome Transport. J. Biol. Chem. 2008, 283, 12635–12642. [Google Scholar] [CrossRef]
- Wu, X.; Wang, F.; Rao, K.; Sellers, J.R.; Hammer, J.A. Rab27a Is an Essential Component of Melanosome Receptor for Myosin Va. Mol. Biol. Cell 2002, 13, 1735–1749. [Google Scholar] [CrossRef]
- Wu, X.; Bowers, B.; Wei, Q.; Kocher, B.; Hammer, J.A. Myosin V Associates with Melanosomes in Mouse Melanocytes: Evidence That Myosin V Is an Organelle Motor. J. Cell Sci. 1997, 110 Pt 7, 847–859. [Google Scholar] [CrossRef]
- Strom, M.; Hume, A.N.; Tarafder, A.K.; Barkagianni, E.; Seabra, M.C. A Family of Rab27-Binding Proteins: Melanophilin Links Rab27a and Myosin Va Function in Melanosome Transport. J. Biol. Chem. 2002, 277, 25423–25430. [Google Scholar] [CrossRef]
- Wu, X.; Rao, K.; Bowers, M.B.; Copeland, N.G.; Jenkins, N.A.; Hammer, J.A. Rab27a Enables Myosin Va-Dependent Melanosome Capture by Recruiting the Myosin to the Organelle. J. Cell Sci. 2001, 114 Pt 6, 1091–1100. [Google Scholar] [CrossRef]
- Ishida, M.; Arai, S.P.; Ohbayashi, N.; Fukuda, M. The GTPase-Deficient Rab27A(Q78L) Mutant Inhibits Melanosome Transport in Melanocytes through Trapping of Rab27A Effector Protein Slac2-a/Melanophilin in Their Cytosol: Development of a Novel Melanosome-Targeting Tag. J. Biol. Chem. 2014, 289, 11059–11067. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Kuroda, T.S.; Mikoshiba, K. Slac2-a/Melanophilin, the Missing Link between Rab27 and Myosin Va: Implications of a Tripartite Protein Complex for Melanosome Transport. J. Biol. Chem. 2002, 277, 12432–12436. [Google Scholar] [CrossRef]
- Hume, A.N.; Collinson, L.M.; Rapak, A.; Gomes, A.Q.; Hopkins, C.R.; Seabra, M.C. Rab27a Regulates the Peripheral Distribution of Melanosomes in Melanocytes. J. Cell Biol. 2001, 152, 795–808. [Google Scholar] [CrossRef]
- Kuroda, T.S.; Fukuda, M. Rab27A-Binding Protein Slp2-a Is Required for Peripheral Melanosome Distribution and Elongated Cell Shape in Melanocytes. Nat. Cell Biol. 2004, 6, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Matesic, L.E.; Yip, R.; Reuss, A.E.; Swing, D.A.; O’Sullivan, T.N.; Fletcher, C.F.; Copeland, N.G.; Jenkins, N.A. Mutations in Mlph, Encoding a Member of the Rab Effector Family, Cause the Melanosome Transport Defects Observed in Leaden Mice. Proc. Natl. Acad. Sci. USA 2001, 98, 10238–10243. [Google Scholar] [CrossRef]
- Kuroda, T.S.; Ariga, H.; Fukuda, M. The Actin-Binding Domain of Slac2-a / Melanophilin Is Required for Melanosome Distribution in Melanocytes. Mol. Cell. Biol. 2003, 23, 5245. [Google Scholar] [CrossRef]
- Jo, C.S.; Park, H.I.; Jung, H.J.; Park, J.I.; Lee, J.E.; Myung, C.H.; Hwang, J.S. A Novel Function of Prohibitin on Melanosome Transport in Melanocytes. Theranostics 2020, 10, 3880–3891. [Google Scholar] [CrossRef]
- Wei, Q.; Wu, X.; Hammer, J.A. The Predominant Defect in Dilute Melanocytes Is in Melanosome Distribution and Not Cell Shape, Supporting a Role for Myosin V in Melanosome Transport. J. Muscle Res. Cell Motil. 1997, 18, 517–527. [Google Scholar] [CrossRef]
- Van Den Bossche, K.; Naeyaert, J.M.; Lambert, J. The Quest for the Mechanism of Melanin Transfer. Traffic 2006, 7, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, R.; Takahashi, Y. Intercellular Transfer of Organelles during Body Pigmentation. Curr. Opin. Genet. Dev. 2017, 45, 132–138. [Google Scholar] [CrossRef]
- Moreiras, H.; Bento-Lopes, L.; Neto, M.V.; Escrevente, C.; Cabaço, L.C.; Hall, M.J.; Ramalho, J.S.; Seabra, M.C.; Barral, D.C. Melanocore Uptake by Keratinocytes Occurs through Phagocytosis and Involves Protease-Activated Receptor-2 Internalization. Traffic 2022, 23, 331–345. [Google Scholar] [CrossRef]
- Hall, M.J.; Lopes-Ventura, S.; Neto, M.V.; Charneca, J.; Zoio, P.; Seabra, M.C.; Oliva, A.; Barral, D.C. Reconstructed Human Pigmented Skin/Epidermis Models Achieve Epidermal Pigmentation through Melanocore Transfer. Pigment Cell Melanoma Res. 2022, 35, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Moreiras, H.; Seabra, M.C.; Barral, D.C. Melanin Transfer in the Epidermis: The Pursuit of Skin Pigmentation Control Mechanisms. Int. J. Mol. Sci. 2021, 22, 4466. [Google Scholar] [CrossRef]
- Swift, J.A. Transfer of Melanin Granules from Melanocytes to the Cortical Cells of Human Hair. Nature 1964, 203, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Tarafder, A.K.; Bolasco, G.; Correia, M.S.; Pereira, F.J.C.; Iannone, L.; Hume, A.N.; Kirkpatrick, N.; Picardo, M.; Torrisi, M.R.; Rodrigues, I.P.; et al. Rab11b Mediates Melanin Transfer between Donor Melanocytes and Acceptor Keratinocytes via Coupled Exo/Endocytosis. J. Investig. Dermatol. 2014, 134, 1056–1066. [Google Scholar] [CrossRef]
- Yamamoto, O.; Bhawan, J. Three Modes of Melanosome Transfers in Caucasian Facial Skin: Hypothesis Based on an Ultrastructural Study. Pigment Cell Res. 1994, 7, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Rachinger, N.; Mittag, N.; Böhme-Schäfer, I.; Xiang, W.; Kuphal, S.; Bosserhoff, A.K. Alpha-Synuclein and Its Role in Melanocytes. Cells 2022, 11, 2087. [Google Scholar] [CrossRef]
- Moreiras, H.; Pereira, F.J.C.; Neto, M.V.; Bento-Lopes, L.; Festas, T.C.; Seabra, M.C.; Barral, D.C. The Exocyst Is Required for Melanin Exocytosis from Melanocytes and Transfer to Keratinocytes. Pigment Cell Melanoma Res. 2020, 33, 366–371. [Google Scholar] [CrossRef]
- Benito-Martínez, S.; Salavessa, L.; Raposo, G.; Marks, M.S.; Delevoye, C. Melanin Transfer and Fate within Keratinocytes in Human Skin Pigmentation. Integr. Comp. Biol. 2021, 61, 1546–1555. [Google Scholar] [CrossRef]
- Singh, S.K.; Kurfurst, R.; Nizard, C.; Schnebert, S.; Perrier, E.; Tobin, D.J. Melanin Transfer in Human Skin Cells Is Mediated by Filopodia—A Model for Homotypic and Heterotypic Lysosome-Related Organelle Transfer. FASEB J. 2010, 24, 3756–3769. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.; Leopardi, S.; Printup, S.; Madden, B.C. Filopodia Are Conduits for Melanosome Transfer to Keratinocytes. J. Cell Sci. 2002, 115 Pt 7, 1441–1451. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes Are Transferred from Melanocytes to Keratinocytes through the Processes of Packaging, Release, Uptake, and Dispersion. J. Investig. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Niki, Y.; Yoshida, M.; Ito, M.; Akiyama, K.; Kim, J.-H.; Yoon, T.-J.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Involvement of Pigment Globules Containing Multiple Melanosomes in the Transfer of Melanosomes from Melanocytes to Keratinocytes. Cell Logist. 2011, 1, 12–20. [Google Scholar] [CrossRef]
- Singh, S.K.; Baker, R.; Sikkink, S.K.; Nizard, C.; Schnebert, S.; Kurfurst, R.; Tobin, D.J. E-Cadherin Mediates Ultraviolet Radiation- and Calcium-Induced Melanin Transfer in Human Skin Cells. Exp. Dermatol. 2017, 26, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, K.A.; Hamilton, N.A.; Moores, M.T.; Brown, D.L.; Ohbayashi, N.; Cairncross, O.; Cook, A.L.; Smith, A.G.; Misaki, R.; Fukuda, M.; et al. The Recycling Endosome Protein Rab17 Regulates Melanocytic Filopodia Formation and Melanosome Trafficking. Traffic 2011, 12, 627–643. [Google Scholar] [CrossRef]
- Seiberg, M. Keratinocyte-Melanocyte Interactions during Melanosome Transfer. Pigment Cell Res. 2001, 14, 236–242. [Google Scholar] [CrossRef]
- Seiberg, M.; Paine, C.; Sharlow, E.; Andrade-Gordon, P.; Costanzo, M.; Eisinger, M.; Shapiro, S.S. The Protease-Activated Receptor 2 Regulates Pigmentation via Keratinocyte-Melanocyte Interactions. Exp Cell Res. 2000, 254, 25–32. [Google Scholar] [CrossRef]
- Sharlow, E.R.; Paine, C.S.; Babiarz, L.; Eisinger, M.; Shapiro, S.; Seiberg, M. The Protease-Activated Receptor-2 Upregulates Keratinocyte Phagocytosis. J. Cell Sci. 2000, 113, 3093–3101. [Google Scholar] [CrossRef]
- Babiarz-Magee, L.; Chen, N.; Seiberg, M.; Lin, C.B. The Expression and Activation of Protease-Activated Receptor-2 Correlate with Skin Color. Pigment Cell Res. 2004, 17, 241–251. [Google Scholar] [CrossRef]
- Sakuraba, K.; Hayashi, N.; Kawashima, M.; Imokawa, G. Down-Regulated PAR-2 Is Associated in Part with Interrupted Melanosome Transfer in Pigmented Basal Cell Epithelioma. Pigment Cell Res. 2004, 17, 371–378. [Google Scholar] [CrossRef]
- Seiberg, M.; Paine, C.; Sharlow, E.; Andrade-Gordon, P.; Costanzo, M.; Eisinger, M.; Shapiro, S.S. Inhibition of Melanosome Transfer Results in Skin Lightening. J. Investig. Dermatol. 2000, 115, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.; Leopardi, S.; Printup, S.; Malhi, N.; Seiberg, M.; LaPoint, R. Proteinase-Activated Receptor-2 Stimulates Prostaglandin Production in Keratinocytes: Analysis of Prostaglandin Receptors on Human Melanocytes and Effects of PGE2 and PGF2α on Melanocyte Dendricity. J. Investig. Dermatol. 2004, 122, 1214–1224. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, D.S.; Sohn, H.; Lee, E.J.; Oh, S.H. PAR-2 Is Involved in Melanogenesis by Mediating Stem Cell Factor Production in Keratinocytes. Exp. Dermatol. 2016, 25, 487–489. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Z.; Wu, W.; Liu, Y.; Xiao, Y.; Qi, D.; Zhao, G.; Zhou, M.; Wang, H.; Liu, J.; et al. TRPA1 Promotes Melanosome Phagocytosis in Keratinocytes via PAR-2/CYLD Axis. J. Dermatol. Sci. 2022, 106, 181–188. [Google Scholar] [CrossRef]
- Finlayson, L.; Barnard, I.R.M.; McMillan, L.; Ibbotson, S.H.; Brown, C.T.A.; Eadie, E.; Wood, K. Depth Penetration of Light into Skin as a Function of Wavelength from 200 to 1000 Nm. Photochem. Photobiol. 2022, 98, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, M.; Krebs, R.; Anders, A.; Heinrich, U.; Tronnier, H. Wavelength-Dependent Penetration Depths of Ultraviolet Radiation in Human Skin. J. Biomed. Opt. 2008, 13, 044030. [Google Scholar] [CrossRef]
- Lepselter, J.; Elman, M. Biological and Clinical Aspects in Laser Hair Removal. J. Dermatolog. Treat. 2004, 15, 72–83. [Google Scholar] [CrossRef]
- Schauer, E.; Trautinger, F.; Köck, A.; Schwarz, A.; Bhardwaj, R.; Simon, M.; Ansel, J.C.; Schwarz, T.; Luger, T.A. Proopiomelanocortin-Derived Peptides Are Synthesized and Released by Human Keratinocytes. J. Clin. Investig. 1994, 93, 2258–2262. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, K.; Kauser, S.; Pritchard, L.E.; Warhurst, A.; Oliver, R.L.; Slominski, A.; Wei, E.T.; Thody, A.J.; Tobin, D.J.; White, A. Proopiomelanocortin (POMC), the ACTH/Melanocortin Precursor, Is Secreted by Human Epidermal Keratinocytes and Melanocytes and Stimulates Melanogenesis. FASEB J. 2007, 21, 1844–1856. [Google Scholar] [CrossRef]
- Slominski, A.; Botchkareva, N.V.; Botchkarev, V.A.; Chakraborty, A.; Luger, T.; Uenalan, M.; Paus, R. Hair Cycle-Dependent Production of ACTH in Mouse Skin. Biochim. Biophys. Acta Mol. Cell Res. 1998, 1448, 147–152. [Google Scholar] [CrossRef]
- Ermak, G.; Slominski, A. Production of POMC, CRH-R1, MC1, and MC2 Receptor MRNA and Expression of Tyrosinase Gene in Relation to Hair Cycle and Dexamethasone Treatment in the C57BL/6 Mouse Skin. J. Investig. Dermatol. 1997, 108, 160–165. [Google Scholar] [CrossRef]
- Benjannet, S.; Rondeau, N.; Day, R.; Chrétien, M.; Seidah, N.G. PC1 and PC2 Are Proprotein Convertases Capable of Cleaving Proopiomelanocortin at Distinct Pairs of Basic Residues. Proc. Natl. Acad. Sci. USA 1991, 88, 3564–3568. [Google Scholar] [CrossRef] [PubMed]
- Kauser, S.; Thody, A.J.; Schallreuter, K.U.; Gummer, C.L.; Tobin, D.J. Β-Endorphin As a Regulator of Human Hair Follicle Melanocyte Biology. J. Investig. Dermatol. 2004, 123, 184–195. [Google Scholar] [CrossRef]
- Tsatmali, M.; Ancans, J.; Thody, A.J. Melanocyte Function and Its Control by Melanocortin Peptides. J. Histochem. Cytochem. 2002, 50, 125–133. [Google Scholar] [CrossRef]
- McLeod, S.D.; Smith, C.; Mason, R.S. Stimulation of Tyrosinase in Human Melanocytes by Pro-Opiomelanocortin- Derived Peptides. J. Endocrinol. 1995, 146, 439–447. [Google Scholar] [CrossRef]
- Hunt, G.; Todd, C.; Kyne, S.; Thody, A.J. ACTH Stimulates Melanogenesis in Cultured Human Melanocytes. J. Endocrinol. 1994, 140, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.E.H.H.; Lee, M.S. In Vivo Effects of MSH on Tyrosinase and Melanogenesis of Pigmentary System. Yale J. Biol. Med. 1973, 46, 493–499. [Google Scholar]
- Suzuki, I.; Cone, R.D.; Im, S.; Nordlund, J.; Abdel-Malek, Z.A. Binding of Melanotropic Hormones to the Melanocortin Receptor MC1R on Human Melanocytes Stimulates Proliferation and Melanogenesis. Endocrinology 1996, 137, 1627–1633. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Malek, Z.; Scott, M.C.; Suzuki, I.; Tada, A.; Im, S.; Lamoreux, L.; Ito, S.; Barsh, G.; Hearing, V.J. The Melanocortin-1 Receptor Is a Key Regulator of Human Cutaneous. Pigment Cell Res. 2000, 13, 156–162. [Google Scholar] [CrossRef]
- Spencer, J.D.; Schallreuter, K.U. Regulation of Pigmentation in Human Epidermal Melanocytes by Functional High-Affinity. Endocrinology 2009, 150, 1250–1258. [Google Scholar] [CrossRef]
- Weatherhead, B.; Logan, A. Interaction of α-Melanocyte-Stimulating Hormone, Melatonin, Cyclic AMP and Cyclic GMP in the Control of Melanogenesis in Hair Follicle Melanocytes in Vitro. J. Endocrinol. 1981, 90, 89–96. [Google Scholar] [CrossRef]
- Moorthy, B.S.; Gao, Y.; Anand, G.S. Phosphodiesterases Catalyze Hydrolysis of CAMP-Bound to Regulatory Subunit of Protein Kinase A and Mediate Signal Termination. Mol. Cell. Proteom. 2011, 10, M110.002295. [Google Scholar] [CrossRef]
- Ballotti, R.; Inserm, U. Cyclic AMP a Key Messenger in the Regulation of Skin Pigmentation. Pigment Cell Res. 2000, 13, 60–69. [Google Scholar]
- Herraiz, C.; Garcia-Borron, J.C.; Jiménez-Cervantes, C.; Olivares, C. MC1R Signaling. Intracellular Partners and Pathophysiological Implications. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2448–2461. [Google Scholar] [CrossRef]
- Horike, N.; Kumagai, A.; Shimono, Y.; Onishi, T.; Itoh, Y.; Kitagawa, K.; Hatano, O.; Takagi, H.; Susumu, T.; Teraoka, H. Downregulation of SIK2 Expression Promotes the Melanogenic Program in Mice. Pigment Cell Melanoma Res. 2010, 23, 809–819. [Google Scholar] [CrossRef]
- Yun, C.Y.; Hong, S.D.; Lee, Y.H.; Lee, J.; Jung, D.E.; Kim, G.H.; Kim, S.H.; Jung, J.K.; Kim, K.H.; Lee, H.; et al. Nuclear Entry of CRTC1 as Druggable Target of Acquired Pigmentary Disorder. Theranostics 2019, 9, 646–660. [Google Scholar] [CrossRef] [PubMed]
- Shoag, J.; Haq, R.; Zhang, M.; Liu, L.; Rowe, G.C.; Jiang, A.; Koulisis, N.; Farrel, C.; Amos, C.I.; Wei, Q.; et al. PGC-1 Coactivators Regulate MITF and the Tanning Response. Mol. Cell 2013, 49, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Grabacka, M.; Wieczorek, J.; Michalczyk-Wetula, D.; Malinowski, M.; Wolan, N.; Wojcik, K.; Plonka, P.M. Peroxisome Proliferator-Activated Receptor α (PPARα) Contributes to Control of Melanogenesis in B16 F10 Melanoma Cells. Arch. Dermatol. Res. 2017, 309, 141–157. [Google Scholar] [CrossRef]
- Kang, H.Y.; Chung, E.; Lee, M.; Cho, Y.; Kang, W.H. Expression and Function of Peroxisome Proliferator-Activated Receptors in Human Melanocytes. Br. J. Dermatol. 2004, 150, 462–468. [Google Scholar] [CrossRef]
- Chen, J.H.; Chang, J.L.; Chen, P.R.; Chuang, Y.J.; Tang, S.T.; Pan, S.F.; Lin, T.B.; Chen, K.H.; Chen, M.J. Inhibition of Peroxisome Proliferator-Activated Receptor Gamma Prevents the Melanogenesis in Murine B16/F10 Melanoma Cells. Biomed. Res. Int. 2014, 2014, 695797. [Google Scholar] [CrossRef]
- Lee, J.S.; Choi, Y.M.; Kang, H.Y. PPAR-Gamma Agonist, Ciglitazone, Increases Pigmentation and Migration of Human Melanocytes. Exp. Dermatol. 2007, 16, 118–123. [Google Scholar] [CrossRef]
- Khaled, M.; Larribere, L.; Bille, K.; Aberdam, E.; Ortonne, J.P.; Ballotti, R.; Bertolotto, C. Glycogen Synthase Kinase 3β Is Activated by CAMP and Plays an Active Role in the Regulation of Melanogenesis. J. Biol. Chem. 2002, 277, 33690–33697. [Google Scholar] [CrossRef] [PubMed]
- Khaled, M.; Larribere, L.; Bille, K.; Ortonne, J.P.; Ballotti, R.; Bertolotto, C. Microphthalmia Associated Transcription Factor Is a Target of the Phosphatidylinositol-3-Kinase Pathway. J. Investig. Dermatol. 2003, 121, 831–836. [Google Scholar] [CrossRef]
- Oka, M.; Nagai, H.; Ando, H.; Fukunaga, M.; Matsumura, M.; Araki, K.; Ogawa, W.; Miki, T.; Sakaue, M.; Tsukamoto, K.; et al. Regulation of Melanogenesis through Phosphatidylinositol 3-Kinase-Akt Pathway in Human G361 Melanoma Cells. J. Investig. Dermatol. 2000, 115, 699–703. [Google Scholar] [CrossRef]
- Bellei, B.; Pitisci, A.; Catricalà, C.; Larue, L.; Picardo, M. Wnt/β-Catenin Signaling Is Stimulated by α-Melanocyte-Stimulating Hormone in Melanoma and Melanocyte Cells: Implication in Cell Differentiation. Pigment Cell Melanoma Res. 2011, 24, 309–325. [Google Scholar] [CrossRef]
- Buscà, R.; Bertolotto, C.; Ortonne, J.P.; Ballotti, R. Inhibition of the Phosphatidylinositol 3-Kinase/P70(S6)-Kinase Pathway Induces B16 Melanoma Cell Differentiation. J. Biol. Chem. 1996, 271, 31824–31830. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Tyburczy, M.E.; Moss, J.; Darling, T.N.; Widlund, H.R.; Kwiatkowski, D.J. Tuberous Sclerosis Complex Inactivation Disrupts Melanogenesis via MTORC1 Activation. J. Clin. Investig. 2017, 127, 349–364. [Google Scholar] [CrossRef]
- Yang, F.; Yang, L.; Wataya-Kaneda, M.; Yoshimura, T.; Tanemura, A.; Katayama, I. Uncoupling of ER/Mitochondrial Oxidative Stress in MTORC1 Hyperactivation-Associated Skin Hypopigmentation. J. Investig. Dermatol. 2018, 138, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.S.; Lee, S.H.; Yun, H.Y.; Baek, K.J.; Kwon, N.S.; Park, K.C.; Kim, D.S. Involvement of MTOR Signaling in Sphingosylphosphorylcholine-Induced Hypopigmentation Effects. J. Biomed. Sci. 2011, 18, 55. [Google Scholar] [CrossRef] [PubMed]
- Ohcuchi, K.; Banno, Y.; Nakagawa, Y.; Akao, Y.; Nozawa, Y. Negative Regulation of Melanogenesis by Phospholipase D1 through MTOR/P70 S6 Kinase 1 Signaling in Mouse B16 Melanoma Cells. J. Cell Physiol. 2005, 205, 444–451. [Google Scholar] [CrossRef]
- Hah, Y.; Cho, H.Y.; Lim, T.; Park, D.H.; Kim, H.M.; Yoon, J.; Kim, J.G.; Kim, C.Y. Induction of Melanogenesis by Rapamycin in Human MNT-1 Melanoma Cells. Ann. Dermatol. 2012, 24, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Kauser, S.; Schallreuter, K.U.; Thody, A.J.; Gummer, C.; Tobin, D.J. Regulation of Human Epidermal Melanocyte Biology by β-Endorphin. J. Investig. Dermatol. 2003, 120, 1073–1080. [Google Scholar] [CrossRef]
- Kauser, S.; Slominski, A.; Wei, E.T.; Tobin, D.J. Modulation of the Human Hair Follicle Pigmentary Unit by Corticotropin-releasing Hormone and Urocortin Peptides. FASEB J. 2006, 20, 882–895. [Google Scholar] [CrossRef]
- Ito, N.; Ito, T.; Kromminga, A.; Bettermann, A.; Takigawa, M.; Kees, F.; Straub, R.H.; Paus, R. Human Hair Follicles Display a Functional Equivalent of the Hypothalamic-pituitary-adrenal (HPA) Axis and Synthesize Cortisol. FASEB J. 2005, 19, 1332–1334. [Google Scholar] [CrossRef] [PubMed]
- Gáspár, E.; Nguyen-Thi, K.T.; Hardenbicker, C.; Tiede, S.; Plate, C.; Bodó, E.; Knuever, J.; Funk, W.; Bíró, T.; Paus, R.; et al. Thyrotropin-Releasing Hormone Selectively Stimulates Human Hair Follicle Pigmentation. J. Investig. Dermatol. 2011, 131, 2368–2377. [Google Scholar] [CrossRef]
- Paus, R. A Neuroendocrinological Perspective on Human Hair Follicle Pigmentation. Pigment Cell Melanoma Res. 2010, 24, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Van Beek, N.; Meyer, K.; Wenzel, E.; Paus, R. Thyroid Hormones Directly Alter Human Hair Follicle Functions: Anagen Prolongation and Stimulation of Both Hair Matrix Keratinocyte Proliferation and Hair Pigmentation. J. Clin. Endocrinol. Metab. 2008, 93, 4381–4388. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Wu, Y.; Yang, T.; Yang, K.; Wang, R.; Yang, J.; Guo, H. Wnt5a Inhibits the Proliferation and Melanogenesis of Melanocytes. Int. J. Med. Sci. 2013, 10, 699–706. [Google Scholar] [CrossRef]
- Hino, S.; Tanji, C.; Nakayama, K.I.; Kikuchi, A. Phosphorylation of β-Catenin by Cyclic AMP-Dependent Protein Kinase Stabilizes β-Catenin through Inhibition of Its Ubiquitination. Mol. Cell Biol. 2005, 25, 9063–9072. [Google Scholar] [CrossRef]
- Schepsky, A.; Bruser, K.; Gunnarsson, G.J.; Goodall, J.; Hallsson, J.H.; Goding, C.R.; Steingrimsson, E.; Hecht, A. The Microphthalmia-Associated Transcription Factor Mitf Interacts with β-Catenin To Determine Target Gene Expression. Mol. Cell Biol. 2006, 26, 8914–8927. [Google Scholar] [CrossRef]
- Bellei, B.; Flori, E.; Izzo, E.; Maresca, V.; Picardo, M. GSK3 β Inhibition Promotes Melanogenesis in Mouse B16 Melanoma Cells and Normal Human Melanocytes. Cell Signal 2008, 20, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Rubinfeld, B.; Albert, I.; Porfiri, E.; Fiol, C.; Munemitsu, S.; Polakis, P. Binding of GSK3beta to the APC-Beta-Catenin Complex and Regulation of Complex Assembly. Science 1996, 272, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, Y.; Semenov, M.; Han, C.; Baeg, G.H.; Tan, Y.; Zhang, Z.; Lin, X.; He, X. Control of β-Catenin Phosphorylation/Degradation by a Dual-Kinase Mechanism. Cell 2002, 108, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Kandyba, E.; Kobielak, K. Wnt7b Is an Important Intrinsic Regulator of Hair Follicle Stem Cell Homeostasis and Hair Follicle Cycling. Stem Cells 2014, 32, 886–901. [Google Scholar] [CrossRef]
- Guo, H.; Yang, K.; Deng, F.; Xing, Y.; Li, Y.; Lian, X.; Yang, T. Wnt3a Inhibits Proliferation but Promotes Melanogenesis of Melan-a Cells. Int. J. Mol. Med. 2012, 30, 636–642. [Google Scholar] [CrossRef]
- Guo, H.; Yang, K.; Deng, F.; Ye, J.; Xing, Y.; Li, Y.; Lian, X.; Yang, T. Wnt3a Promotes Melanin Synthesis of Mouse Hair Follicle Melanocytes. Biochem. Biophys. Res. Commun. 2012, 420, 799–804. [Google Scholar] [CrossRef]
- Guo, H.; Xing, Y.; Liu, Y.; Luo, Y.; Deng, F.; Yang, T.; Yang, K.; Li, Y. Wnt/β-Catenin Signaling Pathway Activates Melanocyte Stem Cells in Vitro and in Vivo. J. Dermatol. Sci. 2016, 83, 45–51. [Google Scholar] [CrossRef]
- Li, Y.H.; Zhang, K.; Yang, K.; Ye, J.X.; Xing, Y.Z.; Guo, H.Y.; Deng, F.; Lian, X.H.; Yang, T. Adenovirus-Mediated Wnt10b Overexpression Induces Hair Follicle Regeneration. J. Investig. Dermatol. 2013, 133, 42–48. [Google Scholar] [CrossRef]
- Dunn, K.J.; Brady, M.; Ochsenbauer-Jambor, C.; Snyder, S.; Incao, A.; Pavan, W.J. WNT1 and WNT3a Promote Expansion of Melanocytes through Distinct Modes of Action. Pigment Cell Res. 2005, 18, 167–180. [Google Scholar] [CrossRef]
- Zhou, J.; Ling, J.; Song, H.; Lv, B.; Wang, L.; Shang, J.; Wang, Y.; Chang, C.; Ping, F.; Qian, J. Neurokinin-1 Receptor Is a Novel Positive Regulator of Wnt / β-Catenin Signaling in Melanogenesis. Oncotarget 2016, 7, 81268–81280. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Passeron, T.; Hoashi, T.; Watabe, H.; Rouzaud, F.; Yasumoto, K.-I.; Hara, T.; Tohyama, C.; Katayama, I.; Miki, T.; et al. Dickkopf 1 (DKK1) Regulates Skin Pigmentation and Thickness by Affecting Wnt/-Catenin Signaling in Keratinocytes. FASEB J. 2007, 22, 1009–1020. [Google Scholar] [CrossRef]
- Park, T.J.; Kim, M.; Kim, H.; Park, S.Y.; Park, K.C.; Ortonne, J.P.; Kang, H.Y. Wnt Inhibitory Factor (WIF)-1 Promotes Melanogenesis in Normal Human Melanocytes. Pigment Cell Melanoma Res. 2014, 27, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Han, J.H.; Kim, J.H.; Park, T.J.; Kang, H.Y. Secreted Frizzled-Related Protein 2 (SFRP2) Functions as a Melanogenic Stimulator; The Role of SFRP2 in UV-Induced Hyperpigmentary Disorders. J. Investig. Dermatol. 2016, 136, 236–244. [Google Scholar] [CrossRef]
- Li, P.H.; Liu, L.H.; Chang, C.C.; Gao, R.; Leung, C.H.; Ma, D.L.; David Wang, H.M. Silencing Stem Cell Factor Gene in Fibroblasts to Regulate Paracrine Factor Productions and Enhance C-Kit Expression in Melanocytes on Melanogenesis. Int. J. Mol. Sci. 2018, 19, 1475. [Google Scholar] [CrossRef]
- Ashman, L.K. The Biology of Stem Cell Factor and Its Receptor C-Kit. Int. J. Biochem. Cell Biol. 1999, 31, 1037–1051. [Google Scholar] [CrossRef]
- Alexeev, V.; Yoon, K. Distinctive Role of the CKit Receptor Tyrosine Kinase Signaling in Mammalian Melanocytes. J. Investig. Dermatol. 2006, 126, 1102–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwano, T.; Terazawa, S.; Nakajima, H.; Imokawa, G. The Stem Cell Factor-Stimulated Melanogenesis in Human Melanocytes Can Be Abrogated by Interrupting the Phosphorylation of MSK1: Evidence for Involvement of the P38/MSK1/CREB/MITF Axis. Arch. Dermatol. Res. 2018, 310, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Ishida, K. Inhibitors of Intracellular Signaling Pathways That Lead to Stimulated Epidermal Pigmentation: Perspective of Anti-Pigmenting Agents. Int. J. Mol. Sci. 2014, 15, 8293–8315. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Jung, S.-H. Recent Development of Signaling Pathways Inhibitors of Melanogenesis. Cell Signal 2017, 40, 99–115. [Google Scholar] [CrossRef]
- Englaro, W.; Bertolotto, C.; Buscà, R.; Brunet, A.; Pagès, G.; Ortonne, J.P.; Ballotti, R. Inhibition of the Mitogen-Activated Protein Kinase Pathway Triggers B16 Melanoma Cell Differentiation. J. Biol. Chem. 1998, 273, 9966–9970. [Google Scholar] [CrossRef]
- Wu, M.; Hemesath, T.J.; Takemoto, C.M.; Horstmann, M.A.; Wells, A.G.; Price, E.R.; Fisher, D.Z.; Fisher, D.E. C-Kit Triggers Dual Phosphorylations, Which Couple Activation and Degradation of the Essential Melanocyte Factor Mi. Genes Dev. 2000, 14, 301–312. [Google Scholar] [CrossRef]
- Xu, W.; Gong, L.; Haddad, M.M.; Bischof, O.; Campisi, J.; Yeh, E.T.H.; Medrano, E.E. Regulation of Microphthalmia-Associated Transcription Factor MITF Protein Levels by Association with the Ubiquitin-Conjugating Enzyme HUBC9. Exp. Cell Res. 2000, 255, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Rönnstrand, L. Signal Transduction via the Stem Cell Factor Receptor/c-Kit. Cell. Mol. Life Sci. 2004, 61, 2535–2548. [Google Scholar] [CrossRef] [PubMed]
- Schönwasser, D.C.; Marais, R.M.; Marshall, C.J.; Parker, P.J. Activation of the Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase Pathway by Conventional, Novel, and Atypical Protein Kinase C Isotypes. Mol. Cell Biol. 1998, 18, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Englaro, W.; Rezzonico, R.; Durand-Clement, M.; Lallemand, D.; Ortonne, J.; Ballotti, R. Mitogen-Activated Protein Kinase Pathway and AP-1 Are Activated during CAMP-Induced Melanogenesis in B-16 Melanoma Cells. J. Biol. Chem. 1995, 270, 24315–24320. [Google Scholar] [CrossRef] [PubMed]
- Hemesath, T.J.; Price, E.R.; Takemoto, C.; Badalian, T.; Fisher, D.E. MAP Kinase Links the Transcription Factor Microphthalmia to C-Kit Signalling in Melanocytes. Nature 1998, 391, 298–301. [Google Scholar] [CrossRef]
- Higuchi, K.; Kawashima, M.; Ichikawa, Y.; Imokawa, G. Sphingosylphosphorylcholine Is a Melanogenic Stimulator for Human Melanocytes. Pigment Cell Res. 2003, 16, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Park, S.H.; Kwon, S.B.; Park, E.S.; Huh, C.H.; Youn, S.W.; Park, K.C. Sphingosylphosphorylcholine-Induced ERK Activation Inhibits Melanin Synthesis in Human Melanocytes. Pigment Cell Res. 2006, 19, 146–153. [Google Scholar] [CrossRef]
- Kim, D.-S.; Park, S.-H.; Kwon, S.-B.; Kwon, N.S.; Park, K.-C. Sphingosylphosphorylcholine Inhibits Melanin Synthesis via Pertussis Toxin-Sensitive MITF Degradation. J. Pharm. Pharmacol. 2010, 62, 181–187. [Google Scholar] [CrossRef]
- Jeong, H.S.; Park, K.C.; Kim, D.S. PP2A and DUSP6 Are Involved in Sphingosylphosphorylcholine-Induced Hypopigmentation. Mol. Cell Biochem. 2012, 367, 43–49. [Google Scholar] [CrossRef]
- Kim, D.S.; Kim, S.Y.; Chung, J.H.; Kim, K.H.; Eun, H.C.; Park, K.C. Delayed ERK Activation by Ceramide Reduces Melanin Synthesis in Human Melanocytes. Cell Signal 2002, 14, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Dong-Seok, K.I.M.; Sook-Young, K.I.M.; Moon, S.J.; Chung, J.H.O.; Kyu-Han, K.I.M.; Kwang-Hyun, C.H.O.; Park, K.C. Ceramide Inhibits Cell Proliferation through Akt/PKB Inactivation and Decreases Melanin Synthesis in Mel-Ab Cells. Pigment Cell Res. 2001, 14, 110–115. [Google Scholar] [CrossRef]
- Jeong, H.S.; Choi, H.R.; Yun, H.Y.; Baek, K.J.; Kwon, N.S.; Park, K.C.; Kim, D.S. Ceramide PC102 Inhibits Melanin Synthesis via Proteasomal Degradation of Microphthalmia-Associated Transcription Factor and Tyrosinase. Mol. Cell Biochem. 2013, 375, 81–87. [Google Scholar] [CrossRef]
- Kim, D.-S.; Hwang, E.-S.; Lee, J.-E.; Kim, S.-Y.; Kwon, S.-B.; Park, K.-C. Sphingosine-1-Phosphate Decreases Melanin Synthesis via Sustained ERK Activation and Subsequent MITF Degradation. J. Cell Sci. 2003, 116, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Park, S.H.; Jeong, Y.M.; Kwon, S.B.; Miller, A.J.; Fisher, D.E.; Park, K.C. Sphingosine-1-Phosphate Decreases Melanin Synthesis via Microphthalmia-Associated Transcription Factor Phosphorylation through the S1P3 Receptor Subtype. J. Pharm. Pharmacol. 2011, 63, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Park, S.H.; Kwon, S.B.; Youn, S.W.; Park, K.C. Effects of Lysophosphatidic Acid on Melanogenesis. Chem. Phys. Lipids 2004, 127, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Hong, A.R.; Kim, Y.H.; Yoo, H.; Kang, S.W.; Chang, S.E.; Song, Y. JNK Suppresses Melanogenesis by Interfering with CREB-Regulated Transcription Coactivator 3-Dependent MITF Expression. Theranostics 2020, 10, 4017–4029. [Google Scholar] [CrossRef]
- Smalley, K.; Eisen, T. The Involvement of P38 Mitogen-Activated Protein Kinase in the α-Melanocyte Stimulating Hormone (α-MSH)-Induced Melanogenic and Anti-Proliferative Effects in B16 Murine Melanoma Cells. FEBS Lett. 2000, 476, 198–202. [Google Scholar] [CrossRef]
- Bellei, B.; Maresca, V.; Flori, E.; Pitisci, A.; Larue, L.; Picardo, M. P38 Regulates Pigmentation Via Proteasomal Degradation of Tyrosinase. J. Biol. Chem. 2010, 285, 7288–7299. [Google Scholar] [CrossRef]
- Saha, B.; Singh, S.K.; Sarkar, C.; Bera, R.; Ratha, J.; Tobin, D.J.; Bhadra, R. Activation of the Mitf Promoter by Lipid-Stimulated Activation of P38-Stress Signalling to CREB. Pigment Cell Res. 2006, 19, 595–605. [Google Scholar] [CrossRef]
- Imokawa, G.; Yada, Y.; Kimura, M. Signalling Mechanisms of Endothelin-Induced Mitogenesis and Melanogenesis in Human Melanocytes. Biochem. J. 1996, 314 Pt 1, 305–312. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, W.; Yuan, X.; Li, D.; Gu, W.; Gao, T. Endothelin-1 Enhances the Melanogenesis via MITF-GPNMB Pathway. BMB Rep. 2013, 46, 364–369. [Google Scholar] [CrossRef]
- Yada, Y.; Higuchi, K.; Imokawa, G. Effects of Endothelins on Signal Transduction and Proliferation in Human Melanocytes. J. Biol. Chem. 1991, 266, 18352–18357. [Google Scholar] [CrossRef]
- Imokawa, G.; Kobayashi, T.; Miyagishi, M.; Higashi, K.; Yada, Y. The Role of Endothelin-1 in Epidermal Hyperpigmentation and Signaling Mechanisms of Mitogenesis and Melanogenesis. Pigment Cell Res. 1997, 10, 218–228. [Google Scholar] [CrossRef]
- Hara, M.; Yaar, M.; Gilchrest, B.A. Endothelin-1 of Keratinocyte Origin Is a Mediator of Melanocyte Dendricity. J. Investig. Dermatol. 1995, 105, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Noh, M. The Regulation of Epidermal Melanogenesis via CAMP and/or PKC Signaling Pathways: Insights for the Development of Hypopigmenting Agents. Arch. Pharm. Res. 2013, 36, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Perez, J.M.; Laursen, R.; Hara, M.; Gilchrest, B.A. Protein Kinase C-β Activates Tyrosinase by Phosphorylating Serine Residues in Its Cytoplasmic Domain. J. Biol. Chem. 1999, 274, 16470–16478. [Google Scholar] [CrossRef]
- Wu, Q.; Fung, A.H.Y.; Xu, M.L.; Poon, K.; Liu, E.Y.L.; Kong, X.P.; Yao, P.; Xiong, Q.P.; Dong, T.T.X.; Tsim, K.W.K. Microphthalmia-Associated Transcription Factor up-Regulates Acetylcholinesterase Expression during Melanogenesis of Murine Melanoma Cells. J. Biol. Chem. 2018, 293, 14417–14428. [Google Scholar] [CrossRef]
- Hasse, S.; Chernyavsky, A.I.; Grando, S.A.; Paus, R. The M4 Muscarinic Acetylcholine Receptor Play a Key Role in the Control of Murine Hair Follicle Cycling and Pigmentation. Life Sci. 2007, 80, 2248–2252. [Google Scholar] [CrossRef] [PubMed]
- Buchli, R.; Ndoye, A.; Arredondo, J.; Webber, R.J.; Grando, S.A. Identification and Characterization of Muscarinic Acetylcholine Receptor Subtypes Expressed in Human Skin Melanocytes. Mol. Cell Biochem. 2001, 228, 57–72. [Google Scholar] [CrossRef]
- Iyengar, B. Modulation of Melanocytic Activity by Acetylcholine. Acta. Anat. 1989, 136, 139–141. [Google Scholar] [CrossRef]
- Hoogduijn, M.J.; Hitchcock, I.S.; Smit, N.P.M.; Gillbro, J.M.; Schallreuter, K.U.; Genever, P.G. Glutamate Receptors on Human Melanocytes Regulate the Expression of MiTF. Pigment Cell Res. 2006, 19, 58–67. [Google Scholar] [CrossRef]
- Yoshida, M.; Takahashi, Y.; Inoue, S. Histamine Induces Melanogenesis and Morphologic Changes by Protein Kinase A Activation via H2receptors in Human Normal Melanocytes. J. Investig. Dermatol. 2000, 114, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Hirotsu, S.; Nakahara, M.; Uchiwa, H.; Tomita, Y. Histamine Is Involved in Ultraviolet B-Induced Pigmentation of Guinea Pig Skin. J. Investig. Dermatol. 2002, 118, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Park, M.K.; Lee, E.J.; Kim, Y.L.; Kim, H.J.; Kang, J.H.; Kim, H.M.; Lee, A.Y.; Lee, C.H. Histamine Receptor 2-Mediated Growth-Differentiation Factor-15 Expression Is Involved in Histamine-Induced Melanogenesis. Int. J. Biochem. Cell Biol. 2012, 44, 2124–2128. [Google Scholar] [CrossRef] [PubMed]
- McEwan, M.T.; Parsons, P.G. Regulation of Tyrosinase Expression and Activity in Human Melanoma Cells via Histamine Receptors. J. Investig. Dermatol. 1991, 97, 868–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fechner, G.A.; Michel, J.; Sturm, R.A.; Jacobs, J.J.; Parsons, P.G. Reduction of DNA Synthesis, Pigment Synthesis, Pigmentation Gene MRNA and Resistance to UVB in Human Melanoma Cells Treated with Analogues of a Histamine (H2) Agonist. Biochem. Pharmacol. 1994, 48, 121–130. [Google Scholar] [CrossRef]
- Gillbro, J.M.; Marles, L.K.; Hibberts, N.A.; Schallreuter, K.U. Autocrine Catecholamine Biosynthesis and the Β2- Adrenoceptor Signal Promote Pigmentation in Human Epidermal Melanocytes. J. Investig. Dermatol. 2004, 123, 346–353. [Google Scholar] [CrossRef]
- Wu, H.L.; Pang, S.L.; Liu, Q.Z.; Wang, Q.; Cai, M.X.; Shang, J. 5-HT1A/1B Receptors as Targets for Optimizing Pigmentary Responses in C57BL/6 Mouse Skin to Stress. PLoS ONE 2014, 9, e89663. [Google Scholar] [CrossRef]
- Lee, H.J.; Park, M.K.; Kim, S.Y.; Choo, H.Y.P.; Lee, A.Y.; Lee, C.H. Serotonin Induces Melanogenesis via Serotonin Receptor 2A. Br. J. Dermatol. 2011, 165, 1344–1348. [Google Scholar] [CrossRef]
- Liu, L.; Zhong, M.; Dong, J.; Chen, M.; Shang, J.; Yue, Y. 5-Hydroxytryptamine (5-HT) Positively Regulates Pigmentation via Inducing Melanoblast Specification and Melanin Synthesis in Zebrafish Embryos. Biomolecules 2020, 10, 1344. [Google Scholar] [CrossRef]
- Wu, H.; Zhao, Y.; Huang, Q.; Cai, M.; Pan, Q.; Fu, M.; An, X.; Xia, Z.; Liu, M.; Jin, Y.; et al. NK1R/5-HT1AR Interaction Is Related to the Regulation of Melanogenesis. FASEB J. 2018, 32, 3193–3214. [Google Scholar] [CrossRef] [PubMed]
- Ping, F.; Shang, J.; Zhou, J.; Song, J.; Zhang, L. Activation of Neurokinin-1 Receptor by Substance P Inhibits Melanogenesis in B16-F10 Melanoma Cells. Int. J. Biochem. Cell Biol. 2012, 44, 2342–2348. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Feng, J.Y.; Wang, Q.; Shang, J. Calcitonin Gene-Related Peptide Cooperates with Substance P to Inhibit Melanogenesis and Induces Apoptosis of B16F10 Cells. Cytokine 2015, 74, 137–144. [Google Scholar] [CrossRef]
- Zhou, J.; Geng, K.; Ping, F.; Gao, Y.; Liu, L.; Feng, B. Cross-Talk between 5-Hydroxytryptamine and Substance P in the Melanogensis and Apoptosis of B16F10 Melanoma Cells. Eur. J. Pharmacol. 2016, 775, 106–112. [Google Scholar] [CrossRef]
- Park, P.J.; Lee, T.R.; Cho, E.G. Substance P Stimulates Endothelin 1 Secretion via Endothelin-Converting Enzyme 1 and Promotes Melanogenesis in Human Melanocytes. J. Investig. Dermatol. 2015, 135, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.K.; Lin, Z.; Tidwell, W.J.; Li, W.; Slominski, A.T. Melatonin and Its Metabolites Accumulate in the Human Epidermis in Vivo and Inhibit Proliferation and Tyrosinase Activity in Epidermal Melanocytes in Vitro. Mol. Cell Endocrinol. 2015, 404, 1–8. [Google Scholar] [CrossRef]
- Slominski, A. Pruski Melatonin Inhibits Proliferation and Melanogenesis in Rodent Melanoma Cells. Exp. Cell Res. 1993, 206, 189–194. [Google Scholar] [CrossRef]
- Singh, S.K.; Abbas, W.A.; Tobin, D.J. Bone Morphogenetic Proteins Differentially Regulate Pigmentation in Human Skin Cells. J. Cell Sci. 2012, 125, 4306–4319. [Google Scholar] [CrossRef] [PubMed]
- Botchkarev, V.A. Bone Morphogenetic Proteins and Their Antagonists in Skin and Hair Follicle Biology. J. Investig. Dermatol. 2003, 120, 36–47. [Google Scholar] [CrossRef]
- Sharov, A.A.; Fessing, M.; Atoyan, R.; Sharova, T.Y.; Haskell-Luevano, C.; Weiner, L.; Funa, K.; Brissette, J.L.; Gilchrest, B.A.; Botchkarev, V.A. Bone Morphogenetic Protein (BMP) Signaling Controls Hair Pigmentation by Means of Cross-Talk with the Melanocortin Receptor-1 Pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 93–98. [Google Scholar] [CrossRef]
- Park, H.; Wu, C.; Yaar, M.; Stachur, C.M.; Kosmadaki, M.; Gilchrest, B.A. Role of BMP-4 and Its Signaling Pathways in Cultured Human Melanocytes. Int. J. Cell Biol. 2009, 2009, 750482. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M.; Wu, C.; Park, H.Y.; Panova, I.; Schutz, G.; Gilchrest, B.A. Bone Morphogenetic Protein-4, a Novel Modulator of Melanogenesis. J. Biol. Chem. 2006, 281, 25307–25314. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Esparza, M.; Ferrer, C.; Castells, M.T.; García-Borrón, J.C.; Zuasti, A. Transforming Growth Factor Β1 Mediates Hypopigmentation of B16 Mouse Melanoma Cells by Inhibition of Melanin Formation and Melanosome Maturation. Int. J. Biochem. Cell Biol. 2001, 33, 971–983. [Google Scholar] [CrossRef]
- Murakami, M.; Matsuzaki, Æ.F. Regulation of Melanin Synthesis by the TGF- b Family in B16 Melanoma Cells. Mol. Biol. Rep. 2009, 36, 1247–1250. [Google Scholar] [CrossRef]
- Kim, D.S.; Park, S.H.; Park, K.C. Transforming Growth Factor-Β1 Decreases Melanin Synthesis via Delayed Extracellular Signal-Regulated Kinase Activation. Int. J. Biochem. Cell Biol. 2004, 36, 1482–1491. [Google Scholar] [CrossRef] [PubMed]
- Jime, C.; Beermann, F.; Aparicio, P.; Lozano, A.; Garcı, C. Transforming Growth Factor- Beta1 Inhibits Basal Melanogenesis in B16/F10 Mouse Melanoma Cells by Increasing the Rate of Degradation of Tyrosinase and Tyrosinase-Related Protein-1. J. Biol. Chem. 1997, 272, 3967–3972. [Google Scholar]
- Park, J.Y.; Kim, M.; Park, T.J.; Kang, H.Y. TGFβ1 Derived from Endothelial Cells Inhibits Melanogenesis. Pigment Cell Melanoma Res. 2016, 29, 477–480. [Google Scholar] [CrossRef]
- Jian, D.; Jiang, D.; Su, J.; Chen, W.; Hu, X.; Kuang, Y.; Xie, H.; Li, J.; Chen, X. Diethylstilbestrol Enhances Melanogenesis via CAMP-PKA-Mediating up-Regulation of Tyrosinase and MITF in Mouse B16 Melanoma Cells. Steroids 2011, 76, 1297–1304. [Google Scholar] [CrossRef]
- McLeod, S.D.; Ranson, M.; Mason, R.S. Effects of Estrogens on Human Melanocytes in Vitro. J. Steroid Biochem. Mol. Biol. 1994, 49, 9–14. [Google Scholar] [CrossRef]
- Sun, M.; Xie, H.; Tang, Y.; Lin, S.Q.; Li, J.M.; Sun, S.N.; Hu, X.L.; Huang, Y.X.; Shi, W.; Jian, D. G Protein-Coupled Estrogen Receptor Enhances Melanogenesis via CAMP-Protein Kinase (PKA) by Upregulating Microphthalmia-Related Transcription Factor-Tyrosinase in Melanoma. J. Steroid Biochem. Mol. Biol. 2017, 165 Pt B, 236–246. [Google Scholar] [CrossRef]
- Natale, C.A.; Duperret, E.K.; Zhang, J.; Sadeghi, R.; Dahal, A.; O’Brien, K.T.; Cookson, R.; Winkler, J.D.; Ridky, T.W. Sex Steroids Regulate Skin Pigmentation through Nonclassical Membrane-Bound Receptors. Elife 2016, 5, e15104. [Google Scholar] [CrossRef] [PubMed]
- Samuelov, L.; Sprecher, E.; Sugawara, K.; Singh, S.K.; Tobin, D.J.; Tsuruta, D.; Bíró, T.; Kloepper, J.E.; Paus, R. Topobiology of Human Pigmentation: P-Cadherin Selectively Stimulates Hair Follicle Melanogenesis. J. Investig. Dermatol. 2013, 133, 1591–1600. [Google Scholar] [CrossRef] [PubMed]
- Arnette, C.R.; Roth-Carter, Q.R.; Koetsier, J.L.; Broussard, J.A.; Burks, H.E.; Cheng, K.; Amadi, C.; Gerami, P.; Johnson, J.L.; Green, K.J. Keratinocyte Cadherin Desmoglein 1 Controls Melanocyte Behavior through Paracrine Signaling. Pigment Cell Melanoma Res. 2020, 33, 305–317. [Google Scholar] [CrossRef]
- Choi, W.; Wolber, R.; Gerwat, W.; Mann, T.; Batzer, J.; Smuda, C.; Liu, H.; Kolbe, L.; Hearing, V.J. The Fibroblast-Derived Paracrine Factor Neuregulin-1 Has a Novel Role in Regulating the Constitutive Color and Melanocyte Function in Human Skin. J. Cell Sci. 2010, 123, 3102–3111. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Jung, H.; Lee, J.H.; Oh, H.Y.; Kim, O.B.; Han, I.O.; Oh, E.S. Keratinocyte-Derived Laminin-332 Protein Promotes Melanin Synthesis via Regulation of Tyrosine Uptake. J. Biol. Chem. 2014, 289, 21751–21759. [Google Scholar] [CrossRef]
- Shi, Q.; Zhang, W.; Guo, S.; Jian, Z.; Li, S.; Li, K.; Ge, R.; Dai, W.; Wang, G.; Gao, T.; et al. Oxidative Stress-Induced Overexpression of MiR-25: The Mechanism Underlying the Degeneration of Melanocytes in Vitiligo. Cell Death Differ. 2016, 23, 496–508. [Google Scholar] [CrossRef] [Green Version]
- Bemis, L.T.; Chen, R.; Amato, C.M.; Classen, E.H.; Robinson, S.E.; Coffey, D.G.; Erickson, P.F.; Shellman, Y.G.; Robinson, W.A. MicroRNA-137 Targets Microphthalmia-Associated Transcription Factor in Melanoma Cell Lines. Cancer Res. 2008, 68, 1362–1368. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, J.F.; Wang, W.M.; Cheung, F.W.K.; Lu, Y.F.; Ng, C.F.; Kung, H.F.; Liu, W.K. MicroRNA-218 Inhibits Melanogenesis by Directly Suppressing Microphthalmia-Associated Transcription Factor Expression. RNA Biol. 2014, 11, 732–741. [Google Scholar] [CrossRef]
- Wang, P.; Li, Y.; Hong, W.; Zhen, J.; Ren, J.; Li, Z.; Xu, A. The Changes of MicroRNA Expression Profiles and Tyrosinase Related Proteins in MITF Knocked down Melanocytes. Mol. Biosyst. 2012, 8, 2924–2931. [Google Scholar] [CrossRef]
- Cicero, A.L.; Delevoye, C.; Gilles-Marsens, F.; Loew, D.; Dingli, F.; Guéré, C.; André, N.; Vié, K.; Van Niel, G.; Raposo, G. Exosomes Released by Keratinocytes Modulate Melanocyte Pigmentation. Nat. Commun. 2015, 6, 7506. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Choi, S.H.; Kim, C.H.; Lee, C.H.; Lee, T.R.; Lee, A.Y. Reduced MiR-675 in Exosome in H19 RNA-Related Melanogenesis via MITF as a Direct Target. J. Investig. Dermatol. 2014, 134, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, J.; Yang, S.; Ji, K.; Liu, X.; Du, B.; Jia, Q.; Qi, S.; Li, X.; Fan, R. Effect of Silencing MicroRNA-508 by STTM on Melanogenesis in Alpaca (Vicugna pacos). Gene 2018, 678, 343–348. [Google Scholar] [CrossRef]
- Itoh, T.; Fukatani, K.; Nakashima, A.; Suzuki, K. MicroRNA-141-3p and MicroRNA-200a-3p Regulate α-Melanocyte Stimulating Hormone-Stimulated Melanogenesis by Directly Targeting Microphthalmia-Associated Transcription Factor. Sci. Rep. 2020, 10, 2149. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Rao, C.; Li, H.; Chen, Y.; Fan, L.; Geng, H.; Li, S.; Qu, J.; Hou, L. Regulation of Pigmentation by MicroRNAs: MITF-Dependent MicroRNA-211 Targets TGF-β Receptor 2. Pigment Cell Melanoma Res. 2015, 28, 217–222. [Google Scholar] [CrossRef]
- Yi, R.; Poy, M.N.; Stoffel, M.; Fuchs, E. A Skin MicroRNA Promotes Differentiation by Repressing “Stemness. ” Nature 2008, 452, 225–229. [Google Scholar] [CrossRef]
- Noguchi, S.; Kumazaki, M.; Yasui, Y.; Mori, T.; Yamada, N.; Akao, Y. MicroRNA-203 Regulates Melanosome Transport and Tyrosinase Expression in Melanoma Cells by Targeting Kinesin Superfamily Protein 5b. J. Investig. Dermatol. 2014, 134, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhao, Y.; Fan, R.; Chen, T.; Dong, C. MicroRNA-21a-5p Functions on the Regulation of Melanogenesis by Targeting Sox5 in Mouse Skin Melanocytes. Int. J. Mol. Sci. 2016, 17, 959. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, P.; Meng, J.; Ji, Y.; Xu, D.; Chen, T.; Fan, R.; Yu, X.; Yao, J.; Dong, C. MicroRNA-27a-3p Inhibits Melanogenesis in Mouse Skin Melanocytes by Targeting Wnt3a. Int. J. Mol. Sci. 2015, 16, 10921–10933. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Bin, B.H.; Kim, J.; Dong, S.E.; Park, P.J.; Choi, H.; Kim, B.J.; Yu, S.J.; Kang, H.; Kang, H.H.; et al. Novel Inhibitory Function of MiR-125b in Melanogenesis. Pigment Cell Melanoma Res. 2014, 27, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Lee, T.R.; Cho, E.G. SH3BP4, a Novel Pigmentation Gene, Is Inversely Regulated by MiR-125b and MITF. Exp. Mol. Med. 2017, 49, e367. [Google Scholar] [CrossRef] [PubMed]
- Dynoodt, P.; Mestdagh, P.; Van Peer, G.; Vandesompele, J.; Goossens, K.; Peelman, L.J.; Geusens, B.; Speeckaert, R.M.; Lambert, J.L.W.; Van Gele, M.J.L. Identification of MiR-145 as a Key Regulator of the Pigmentary Process. J. Investig. Dermatol. 2012, 133, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.T.; Chen, J.S.; Chang, D.C.; Lin, S.-L. Mir-434-5p Mediates Skin Whitening and Lightening. Clin. Cosmet. Investig. Dermatol. 2008, 1, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Yu, X.; Dong, C. MiR-137 Affects Melanin Synthesis in Mouse Melanocyte by Repressing the Expression of c-Kit and Tyrp2 in SCF/c-Kit Signaling Pathway. Biosci. Biotechnol. Biochem. 2016, 80, 2115–2121. [Google Scholar] [CrossRef]
- Qi, S.; Liu, B.; Zhang, J.; Liu, X.; Dung, C.; Fan, R. Knockdown of MicroRNA-143-5p by STTM Technology Affects Eumelanin and Pheomelanin Production in Melanocytes. Mol. Med. Rep. 2019, 20, 2649–2656. [Google Scholar] [CrossRef]
- Lin, K.-Y.; Chen, C.-M.; Lu, C.-Y.; Cheng, C.-Y. Regulation of MiR-21 Expression in Human Melanoma via UV-Ray-Induced Melanin Pigmentation. Environ. Toxicol. 2017, 32, 2064–2069. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, L.; Gao, H.; Chang, L.; Yu, X.; Zhu, Z.; He, X.; Geng, J.; Dong, Y.; Li, H.; et al. Exosomal MiRNA Derived from Keratinocytes Regulates Pigmentation in Melanocytes. J. Dermatol. Sci. 2019, 93, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Šahmatova, L.; Tankov, S.; Prans, E.; Aab, A.; Hermann, H.; Reemann, P.; Pihlap, M.; Karelson, M.; Abram, K.; Kisand, K.; et al. MicroRNA-155 Is Dysregulated in the Skin of Patients with Vitiligo and Inhibits Melanogenesis-Associated Genes in Melanocytes and Keratinocytes. Acta Derm. Venereol. 2016, 96, 742–747. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, D.; Wang, X.; Mao, Y.; Xu, Z.; Sun, Y.; Mei, X.; Song, J.; Shi, W. Down-Regulation of Exosomal MiR-200c Derived from Keratinocytes in Vitiligo Lesions Suppresses Melanogenesis. J. Cell Mol. Med. 2020, 24, 12164–12175. [Google Scholar] [CrossRef]
- Zeng, Q.; Wang, Q.; Chen, X.; Xia, K.; Tang, J.; Zhou, X.; Cheng, Y.; Chen, Y.; Huang, L.; Xiang, H.; et al. Analysis of LncRNAs Expression in UVB-Induced Stress Responses of Melanocytes. J. Dermatol. Sci. 2016, 81, 53–60. [Google Scholar] [CrossRef]
- Ji, K.; Fan, R.; Zhang, J.; Yang, S.; Dong, C. Long Non-Coding RNA Expression Profile in Cdk5-Knockdown Mouse Skin. Gene 2018, 672, 195–201. [Google Scholar] [CrossRef]
- Pei, S.; Chen, J.; Lu, J.; Hu, S.; Jiang, L.; Lei, L.; Ouyang, Y.; Fu, C.; Ding, Y.; Li, S.; et al. The Long Noncoding RNA UCA1 Negatively Regulates Melanogenesis in Melanocytes. J. Investig. Dermatol. 2020, 140, 152–163.e5. [Google Scholar] [CrossRef]
- Ho, J.C.; Lee, C.H.; Hong, C.H. Targeting Steroid Receptor RNA Activator (SRA), a Long Non-Coding RNA, Enhances Melanogenesis through Activation of TRP1 and Inhibition of P38 Phosphorylation. PLoS ONE 2020, 15, e0237577. [Google Scholar] [CrossRef]
- Hale, L.P. Zinc α-2-Glycoprotein Regulates Melanin Production by Normal and Malignant Melanocytes. J. Investig. Dermatol. 2002, 119, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, T.; Matsuda, M.; Yamashita, Y.; Takehara, M.; Fukuya, M.; Mineda, K.; Maji, D.; Ihn, H.; Adachi, H.; Sobue, G.; et al. Suppression of Melanin Production by Expression of HSP70. J. Biol. Chem. 2010, 285, 13254–13263. [Google Scholar] [CrossRef] [PubMed]
- Murase, D.; Hachiya, A.; Fullenkamp, R.; Beck, A.; Moriwaki, S.; Hase, T.; Takema, Y.; Manga, P. Variation in Hsp70-1A Expression Contributes to Skin Color Diversity. J. Investig. Dermatol. 2016, 136, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Malek, Z.A. Fueling Melanocytes with ATP from Keratinocytes Accelerates Melanin Synthesis. J. Investig. Dermatol. 2019, 139, 1424–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelis, L.; Jovancevic, N.; Veitinger, S.; Mandal, B.; Arndt, H.D.; Neuhaus, E.M.; Hatt, H. Functional Characterization of the Odorant Receptor 51E2 in Human Melanocytes. J. Biol. Chem. 2016, 291, 17772–17786. [Google Scholar] [CrossRef]
- Wojcik, S.; Weidinger, D.; Ständer, S.; Luger, T.; Hatt, H.; Jovancevic, N. Functional Characterization of the Extranasal OR2A4/7 Expressed in Human Melanocytes. Exp. Dermatol. 2018, 27, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Luecke, S.; Backlund, M.; Jux, B.; Esser, C.; Krutmann, J.; Rannug, A. The Aryl Hydrocarbon Receptor (AHR), a Novel Regulator of Human Melanogenesis. Pigment Cell Melanoma Res. 2010, 23, 828–833. [Google Scholar] [CrossRef]
- Pucci, M.; Pasquariello, N.; Battista, N.; Di Tommaso, M.; Rapino, C.; Fezza, F.; Zuccolo, M.; Jourdain, R.; Agrò, A.F.; Breton, L.; et al. Endocannabinoids Stimulate Human Melanogenesis via Type-1 Cannabinoid Receptor. J. Biol. Chem. 2012, 287, 15466–15478. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Nakanishi, Y.; Valenzuela, R.K.; Brilliant, M.H.; Kolbe, L.; Wakamatsu, K. Usefulness of Alkaline Hydrogen Peroxide Oxidation to Analyze Eumelanin and Pheomelanin in Various Tissue Samples: Application to Chemical Analysis of Human Hair Melanins. Pigment Cell Melanoma Res. 2011, 24, 605–613. [Google Scholar] [CrossRef]
- Pavan, W.J.; Sturm, R.A. The Genetics of Human Skin and Hair Pigmentation. Annu. Rev. Genom. Hum. Genet. 2019, 20, 41–72. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.D.; Pairo-Castineira, E.; Rawlik, K.; Canela-Xandri, O.; Rees, J.; Sims, D.; Tenesa, A.; Jackson, I.J. Genome-Wide Study of Hair Colour in UK Biobank Explains Most of the SNP Heritability. Nat. Commun. 2018, 9, 5271. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, K.A.; Shekar, S.L.; Newton, R.A.; James, M.R.; Stow, J.L.; Duffy, D.L.; Sturm, R.A. Receptor Function, Dominant Negative Activity and Phenotype Correlations for MC1R Variant Alleles. Hum. Mol. Genet. 2007, 16, 2249–2260. [Google Scholar] [CrossRef]
- Gerstenblith, M.R.; Goldstein, A.M.; Fargnoli, M.C.; Peris, K.; Landi, M.T. Comprehensive Evaluation of Allele Frequency Differences of MC1R Variants across Populations. Hum. Mutat. 2007, 28, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Naysmith, L.; Waterston, K.; Ha, T.; Flanagan, N.; Bisset, Y.; Ray, A.; Wakamatsu, K.; Ito, S.; Rees, J.L. Quantitative Measures of the Effect of the Melanocortin 1 Receptor on Human Pigmentary Status. J. Investig. Dermatol. 2004, 122, 423–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valverde, P.; Healy, E.; Jackson, I.; Rees, J.L.; Thody, A.J. Variants of the Melanocyte–Stimulating Hormone Receptor Gene Are Associated with Red Hair and Fair Skin in Humans. Nat. Genet. 1995, 11, 328–330. [Google Scholar] [CrossRef]
- Walker, W.P.; Gunn, T.M. Shades of Meaning: The Pigment-Type Switching System as a Tool for Discovery. Pigment Cell Melanoma Res. 2010, 23, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Rees, J.L. Genetics of Hair and Skin Color. Annu Rev Genet 2003, 37, 67–90. [Google Scholar] [CrossRef]
- Sulem, P.; Gudbjartsson, D.F.; Stacey, S.N.; Helgason, A.; Rafnar, T.; Jakobsdottir, M.; Steinberg, S.; Gudjonsson, S.A.; Palsson, A.; Thorleifsson, G.; et al. Two Newly Identified Genetic Determinants of Pigmentation in Europeans. Nat. Genet. 2008, 40, 835–837. [Google Scholar] [CrossRef]
- Le Pape, E.; Wakamatsu, K.; Ito, S.; Wolber, R.; Hearing, V.J. Regulation of Eumelanin / Pheomelanin Synthesis and Visible Pigmentation in Melanocytes by Ligands of the Melanocortin 1 Receptor. Pigment Cell Melanoma Res. 2008, 21, 477–486. [Google Scholar] [CrossRef]
- Siegrist, W.; Willard, D.H.; Wilkison, W.O.; Eberle, A.N. Agouti Protein Inhibits Growth of B16 Melanoma Cells in Vitro by Acting through Melanocortin Receptors. Biochem. Biophys. Res. Commun. 1996, 218, 171–175. [Google Scholar] [CrossRef]
- Suzuki, I.; Tada, A.; Ollmann, M.M.; Barsh, G.S.; Im, S.; Lamoreux, M.L.; Hearing, V.J.; Nordlund, J.J.; Abdel-Malek, Z.A. Agouti Signaling Protein Inhibits Melanogenesis and the Response of Human Melanocytes to α-Melanotropin. J. Investig. Dermatol. 1997, 108, 838–842. [Google Scholar] [CrossRef]
- Siegrist, W.; Drozdz, R.; Cotti, R.; Willard, D.H.; Wilkison, W.O.; Eberle, A.N. Interactions of α-Melanotropin and Agouti on B16 Melanoma Cells: Evidence for Inverse Agonism of Agouti. J. Recept. Signal Transduct. Res. 1997, 17, 75–98. [Google Scholar] [CrossRef] [PubMed]
- Branicki, W.; Liu, F.; Van Duijn, K.; Draus-Barini, J.; Pośpiech, E.; Walsh, S.; Kupiec, T.; Wojas-Pelc, A.; Kayser, M. Model-Based Prediction of Human Hair Color Using DNA Variants. Hum. Genet. 2011, 129, 443–454. [Google Scholar] [CrossRef]
- Lin, B.D.; Mbarek, H.; Willemsen, G.; Dolan, C.V.; Fedko, I.O.; Abdellaoui, A.; De Geus, E.J.; Boomsma, D.I.; Hottenga, J.J. Heritability and Genome-Wide Association Studies for Hair Color in a Dutch Twin Family Based Sample. Genes 2015, 6, 559–576. [Google Scholar] [CrossRef] [Green Version]
- Kenny, E.E.; Timpson, N.J.; Sikora, M.; Yee, M.C.; Moreno-Estrada, A.; Eng, C.; Huntsman, S.; Burchard, E.G.; Stoneking, M.; Bustamante, C.D.; et al. Melanesian Blond Hair Is Caused by an Amino Acid Change in TYRP1. Science 2012, 336, 554. [Google Scholar] [CrossRef]
- Norton, H.L.; Correa, E.A.; Koki, G.; Friedlaender, J.S. Distribution of an Allele Associated with Blond Hair Color across Northern Island Melanesia. Am. J. Phys. Anthropol. 2014, 153, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Ambrosio, A.L.; Boyle, J.A.; Aradi, A.E.; Christian, K.A.; Di Pietro, S.M. TPC2 Controls Pigmentation by Regulating Melanosome PH and Size. Proc. Natl. Acad. Sci. USA 2016, 113, 5622–5627. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.K.; Schludi, V.; Chen, C.C.; Butz, E.; Nguyen, O.N.P.; Müller, M.; Krüger, J.; Kammerbauer, C.; Ben-Johny, M.; Vollmar, A.M.; et al. TPC2 Polymorphisms Associated with a Hair Pigmentation Phenotype in Humans Result in Gain of Channel Function by Independent Mechanisms. Proc. Natl. Acad. Sci. USA 2017, 114, E8595–E8602. [Google Scholar] [CrossRef]
- Guenther, C.A.; Tasic, B.; Luo, L.; Bedell, M.A.; Kingsley, D.M. A Molecular Basis for Classic Blond Hair Color in Europeans. Nat. Genet. 2014, 46, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Kraft, P.; Nan, H.; Guo, Q.; Chen, C.; Qureshi, A.; Hankinson, S.E.; Hu, F.B.; Duffy, D.L.; Zhen, Z.Z.; et al. A Genome-Wide Association Study Identifies Novel Alleles Associated with Hair Color and Skin Pigmentation. PLoS Genet. 2008, 4, e1000074. [Google Scholar] [CrossRef]
- Sulem, P.; Gudbjartsson, D.F.; Stacey, S.N.; Helgason, A.; Rafnar, T.; Magnusson, K.P.; Manolescu, A.; Karason, A.; Palsson, A.; Thorleifsson, G.; et al. Genetic Determinants of Hair, Eye and Skin Pigmentation in Europeans. Nat. Genet. 2007, 39, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Böck, J.; Krogsaeter, E.; Passon, M.; Chao, Y.K.; Sharma, S.; Grallert, H.; Peters, A.; Grimm, C. Human Genome Diversity Data Reveal That L564P Is the Predominant TPC2 Variant and a Prerequisite for the Blond Hair Associated M484L Gain-of-Function Effect. PLoS Genet. 2021, 17, e1009236. [Google Scholar] [CrossRef]
- Branicki, W.; Brudnik, U.; Wojas-pelc, A. Interactions Between HERC2, OCA2 and MC1R May Influence Human Pigmentation Phenotype. Ann. Hum. Genet. 2009, 73, 160–170. [Google Scholar] [CrossRef]
- Graf, J.; Hodgson, R.; Daal, A. Van Single Nucleotide Polymorphisms in the MATP Gene Are Associated With Normal Human Pigmentation Variation. Hum. Mutat. 2005, 25, 278–284. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.D.B.; Nicu, C.; Picard, M.; Chéret, J.; Bedogni, B.; Tobin, D.J.; Paus, R. The Biology of Human Hair Greying. Biol. Rev. 2021, 96, 107–128. [Google Scholar] [CrossRef]
- Jo, S.K.; Lee, J.Y.; Lee, Y.; Kim, C.D.; Lee, J.H.; Lee, Y.H. Three Streams for the Mechanism of Hair Graying. Ann. Dermatol. 2018, 30, 397–401. [Google Scholar] [CrossRef]
- Commo, S.; Gaillard, O.; Bernard, B.A. Human Hair Greying Is Linked to a Specific Depletion of Hair Follicle Melanocytes Affecting Both the Bulb and the Outer Root Sheath. Br. J. Dermatol. 2004, 150, 435–443. [Google Scholar] [CrossRef]
- Hegedus, Z.L. The Probable Involvement of Soluble and Deposited Melanins, Their Intermediates and the Reactive Oxygen Side-Products in Human Diseases and Aging. Toxicology 2000, 145, 85–101. [Google Scholar] [CrossRef]
- Wood, J.M.; Decker, H.; Hartmann, H.; Chavan, B.; Rokos, H.; Spencer, J.D.; Hasse, S.; Thornton, M.J.; Shalbaf, M.; Paus, R.; et al. Senile Hair Graying: H2O2-mediated Oxidative Stress Affects Human Hair Color by Blunting Methionine Sulfoxide Repair. FASEB J. 2009, 23, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Schallreuter, K.U.; Salem, M.M.A.E.L.; Hasse, S.; Rokos, H. The Redox—Biochemistry of Human Hair Pigmentation. Pigment Cell Melanoma Res. 2010, 24, 51–62. [Google Scholar] [CrossRef]
- Arck, P.C.; Overall, R.; Spatz, K.; Liezman, C.; Handjiski, B.; Klapp, B.F.; Birch-Machin, M.A.; Peters, E.M.J.; Arck, P.C.; Overall, R.; et al. Towards a “Free Radical Theory of Graying”: Melanocyte Apoptosis in the Aging Human Hair Follicle Is an Indicator of Oxidative Stress Induced Tissue Damage. FASEB J. 2006, 20, 1567–1569. [Google Scholar] [CrossRef]
- Seiberg, M. Age-Induced Hair Greying—The Multiple Effects of Oxidative Stress. Int J Cosmet Sci 2013, 35, 532–538. [Google Scholar] [CrossRef]
- Ibrahim, L.; Wright, E.A. The Long Term Effect of Repeated Pluckings on the Function of the Mouse Vibrissal Follicles. British J. Dermatol. 1978, 99, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, E.K.; Granter, S.R.; Fisher, D.E. Mechanisms of Hair Graying: Incomplete Melanocyte Stem Cell Maintenance in the Niche. Science 2005, 307, 720–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, E.K.; Suzuki, M.; Igras, V.; Du, J.; Lonning, S.; Miyachi, Y.; Roes, J.; Beermann, F.; Fisher, D.E. Key Roles for Transforming Growth Factor β in Melanocyte Stem Cell Maintenance. Cell Stem Cell 2010, 6, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Valluet, A.; Druillennec, S.; Barbotin, C.; Dorard, C.; Monsoro-Burq, A.H.; Larcher, M.; Pouponnot, C.; Baccarini, M.; Larue, L.; Eychène, A. B-Raf and C-Raf Are Required for Melanocyte Stem Cell Self-Maintenance. Cell Rep. 2012, 2, 774–780. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Namasivayam, V.; Manickam, M.; Jung, S.-H. Inhibitors of Melanogenesis: An Updated Review. J. Med. Chem. 2018, 61, 7395–7418. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Chen, J.; Lu, J.; Yi, L.; Tong, X.; Kang, L.; Pei, S.; Ouyang, Y.; Jiang, L.; Ding, Y.; et al. Roles of Inflammation Factors in Melanogenesis (Review). Mol. Med. Rep. 2020, 21, 1421–1430. [Google Scholar] [CrossRef]
- Bertolini, M.; McElwee, K.; Gilhar, A.; Bulfone-Paus, S.; Paus, R. Hair Follicle Immune Privilege and Its Collapse in Alopecia Areata. Exp. Dermatol. 2020, 29, 703–725. [Google Scholar] [CrossRef]
- Costa, C.; Cavaco-Paulo, A.; Matamá, T. Mapping Hair Follicle-Targeted Delivery by Particle Systems: What Has Science Accomplished so Far? Int. J. Pharm. 2021, 610, 121273. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.H.; Jin, Z.H. Paracrine Regulation of Melanogenesis. Br. J. Dermatol. 2018, 178, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Lona-Durazo, F.; Mendes, M.; Thakur, R.; Funderburk, K.; Zhang, T.; Kovacs, M.A.; Choi, J.; Brown, K.M.; Parra, E.J. A Large Canadian Cohort Provides Insights into the Genetic Architecture of Human Hair Colour. Commun. Biol. 2021, 4, 1253. [Google Scholar] [CrossRef]
- Pośpiech, E.; Wojas-Pelc, A.; Walsh, S.; Liu, F.; Maeda, H.; Ishikawa, T.; Skowron, M.; Kayser, M.; Branicki, W. The Common Occurrence of Epistasis in the Determination of Human Pigmentation and Its Impact on DNA-Based Pigmentation Phenotype Prediction. Forensic. Sci. Int. Genet. 2014, 11, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Fleming, C.; MacKie, R. Alpha Interferon-Induced Hair Discolouration. Br J Dermatol. 1996, 135, 337–338. [Google Scholar] [CrossRef]
- Seckin, D.; Yildiz, A. Repigmentation and Curling of Hair after Acitretin Therapy. Australas. J. Dermatol. 2009, 50, 214–216. [Google Scholar] [CrossRef]
- Ward, P.D.; Miller, H.L.; Shipman, A.R. A Case of Repigmentation and Curling of Hair on Acitretin Therapy. Clin. Exp. Dermatol. 2014, 39, 91–92. [Google Scholar] [CrossRef] [PubMed]
- Rivera, N.; Boada, A.; Bielsa, M.I.; Fernández-Figueras, M.T.; Carcereny, E.; Moran, M.T.; Ferrándiz, C. Hair Repigmentation during Immunotherapy Treatment with an Anti–Programmed Cell Death 1 and Anti–Programmed Cell Death Ligand 1 Agent for Lung Cancer. JAMA Dermatol. 2017, 153, 1162–1165. [Google Scholar] [CrossRef] [PubMed]
- Donovan, J.C.; Price, V.H. Chloroquine-Induced Hair Hypopigmentation. N. Engl. J. Med. 2010, 363, 372. [Google Scholar] [CrossRef]
- Di Giacomo, T.B.; Valente, N.Y.S.; Nico, M.M.S. Chloroquine—Induced Hair Depigmentation. Lupus 2009, 18, 264–266. [Google Scholar] [CrossRef]
- Robinson, A.; Jones, W. Changes in Scalp Hair after Cancer Chemotherapy. Eur. J. Cancer Clin. Oncol. 1989, 25, 155–156. [Google Scholar] [CrossRef]
- Sadighha, A.; Zahed, G. Hair Darkening after Treatment with Cyclosporin in a Patient with Psoriasis. J. Eur. Acad. Dermatol. Venereol. 2008, 2, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Dika, E.; Patrizi, A.; Ribero, S.; Fanti, P.A.; Starace, M.; Melotti, B.; Sperandi, F.; Piraccini, B.M. Hair and Nail Adverse Events during Treatment with Targeted Therapies for Metastatic Melanoma. Eur. J. Dermatol. 2016, 26, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Fujimi, A.; Ibata, S.; Kanisawa, Y.; Shibata, T.; Sakamoto, H.; Yamada, S.; Okuda, T.; Takahashi, S.; Minami, S.; Hashimoto, A. Reversible Skin and Hair Depigmentation during Chemotherapy with Dasatinib for Chronic Myeloid Leukemia. J. Dermatol. 2016, 43, 104–106. [Google Scholar] [CrossRef]
- Rubegni, P.; Sbano, P.; Fimiani, M. A Case of Disseminated Granuloma Annulare Treated with Defibrotide: Complete Clinical Remission and Progressive Hair Darkening. Br. J. Dermatol. 2003, 149, 422–424. [Google Scholar] [CrossRef] [PubMed]
- Sumitomo, C.; Akita, H.; Sugiura, K. Unexpected Side-Effect of Dupilumab: Reversal of Hair Graying. J. Dermatol. 2020, 47, e316–e317. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Chen, H.; Chiu, H. Erlotinib-Induced Hair Pigmentation. Int. J. Dermatol. 2014, 53, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.; Alsaleh, Q.A. Hair Discoloration Caused by Etretinate. Dermatology 1994, 188, 172. [Google Scholar] [CrossRef]
- Nagase, K.; Inoue, T.; Narisawa, Y. Manifest Hair Repigmentation Associated with Etretinate Therapy. J. Dermatol. 2017, 44, e34–e35. [Google Scholar] [CrossRef] [PubMed]
- Vesper, J.L.; Fenske, N.A.; Tampa, M.D. Hair Darkening and New Growth Associated with Etretinate Therapy. J. Am. Acad. Dermatol. 1996, 34, 860. [Google Scholar] [CrossRef] [PubMed]
- Meiler, S.; Gerber, P.A.; Homey, B. Clinical Image: Blonde by Prescription. Arthritis Rheum. 2008, 58, 2286. [Google Scholar] [CrossRef]
- Yun, S.; Song, K.; Hwang, S.; Kim, H.; Lee, N.; Park, J. Hair Graying and Loss Induced by Imatinib Mesylate. J. Dermatol. 2014, 41, 107–108. [Google Scholar] [CrossRef] [PubMed]
- Mariani, S.; Abruzzese, E.; Basciani, S.; Fiore, D.; Persichetti, A.; Watanabe, M.; Spera, G.; Gnessi, L. Reversible Hair Depigmentation in a Patient Treated with Imatinib. Leuk. Res. 2011, 35, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Redondo, P.; Guzmán, M.; Marquina, M.; Pretel, M.; Lloret, L.A.P.; Gorrochategui, A. Repigmentation of Gray Hair After Thyroid Hormone Treatment. Actas. Dermosifiliogr. 2007, 98, 603–610. [Google Scholar] [CrossRef]
- Bellandi, S.; LAmato, L.; Cipollini, E.; Antiga, E.; Brandini, L.; Fabbri, P. Repigmentation of Hair after Latanoprost Therapy. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 2010–2012. [Google Scholar] [CrossRef]
- Dasanu, C.A. Hair Repigmentation Associated with the Use of Lenalidomide: Graying May Not Be an Irreversible Process ! J. Oncol. Pharm. Pract. 2012, 19, 165–169. [Google Scholar] [CrossRef]
- Reynolds, N.J.; Crossley, J.; Ferguson, I.; Peachey, R.D.G. Darkening of White Hair in Parkinson’s Disease. Clin. Exp. Dermatol. 1989, 14, 317–318. [Google Scholar] [CrossRef]
- Zarafonetis, C.J. Darkening of Gray Hair During Para-Amino-Benzoic Acid Therapy. J. Investig. Dermatol. 1950, 15, 399–402. [Google Scholar] [CrossRef]
- Sideras, K.; Menefee, M.E.; Burton, J.K.; Erlichman, C.; Bible, K.C.; Ivy, S.P. Profound Hair and Skin Hypopigmentation in an African American Woman Treated with the Multi-Targeted Tyrosine Kinase Inhibitor Pazopanib. J. Clin. Oncol. 2010, 28, 312–313. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.; Koyama, T.; Kobayashi, K.; Setsu, N.; Kawashima, M.; Kawai, A. Reversible Hair Depigmentation in a Japanese Female Treated with Pazopanib. J. Dermatol. 2014, 41, 1021–1022. [Google Scholar] [CrossRef] [PubMed]
- Dermatol, A. Repigmentation of the White Hair after Systemic Corticosteroids for Bullous Pemphigoid. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 1018–1020. [Google Scholar] [CrossRef]
- Baldo, F.; Magnolato, A.; Barbi, E.; Bruno, I. Selumetinib Side Effects in Children Treated for Plexiform Neurofibromas: First Case Reports of Peripheral Edema and Hair Color Change. BMC Pediatr. 2021, 21, 67. [Google Scholar] [CrossRef]
- Hartmann, J.; Kanz, L. Sunitinib and Periodic Hair Depigmentation Due to Temporary C-KIT Inhibition. Arch. Dermatol. 2008, 144, 1525–1526. [Google Scholar] [CrossRef] [PubMed]
- Hampson, J.; Donnelly, A.; Lewis-Jones, M.; Pye, J. Tamoxifen-Induced Hair Colour Change. Br. J. Dermatol. 1991, 132, 483–485. [Google Scholar] [CrossRef]
- Lovering, S.; Miao, W.; Bailie, T.; Amato, D. Hair Repigmentation Associated with Thalidomide Use for the Treatment of Multiple Myeloma. BMJ Case Rep. 2016, 2016, bcr2016215521. [Google Scholar] [CrossRef]
- Karamizadeh, Z.; Rasekhi, A.R. Gray Hair in Children on Triptorelin Treatment. Int. J. Dermatol. 2008, 47, 601–604. [Google Scholar] [CrossRef]
- Park, S.J.; Ahn, G.R.; Park, J.W.; Seo, S.J. The First Case of Ustekinumab-Associated Hair Repigmentation and a Proposed Mechanism of Action. Ann. Dermatol. 2021, 33, 300–301. [Google Scholar] [CrossRef]
- Gerstner, T.; Lipinski, C.; Longin, E.; König, S. Valproate-Induced Change in Hair Color. J. Am. Acad. Dermatol. 2008, 58, 63–64. [Google Scholar] [CrossRef]
- Read, G. Verapamil and Hair Colour Change. Lancet 1991, 338, 1520. [Google Scholar] [CrossRef] [PubMed]










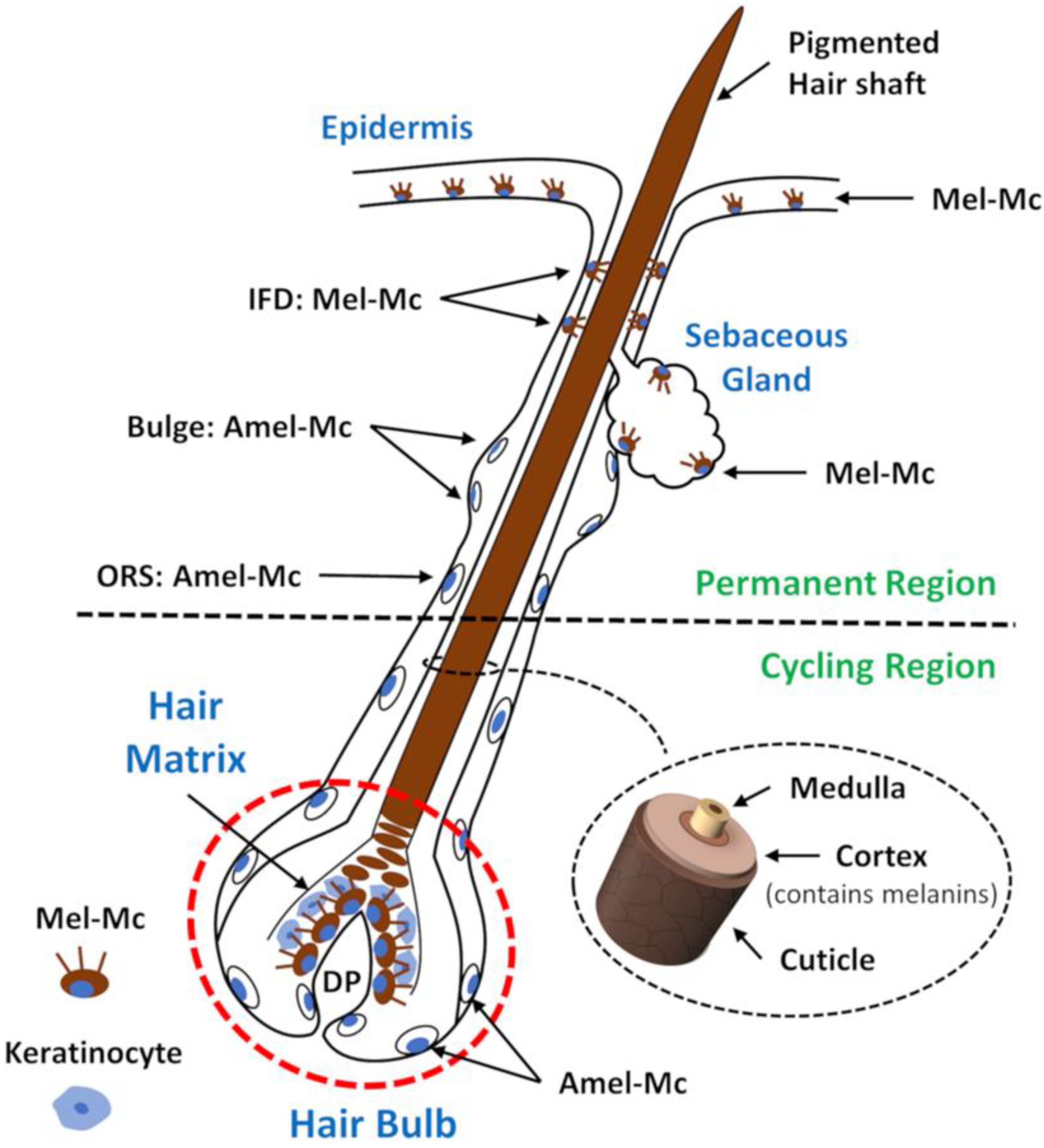{kind=link}
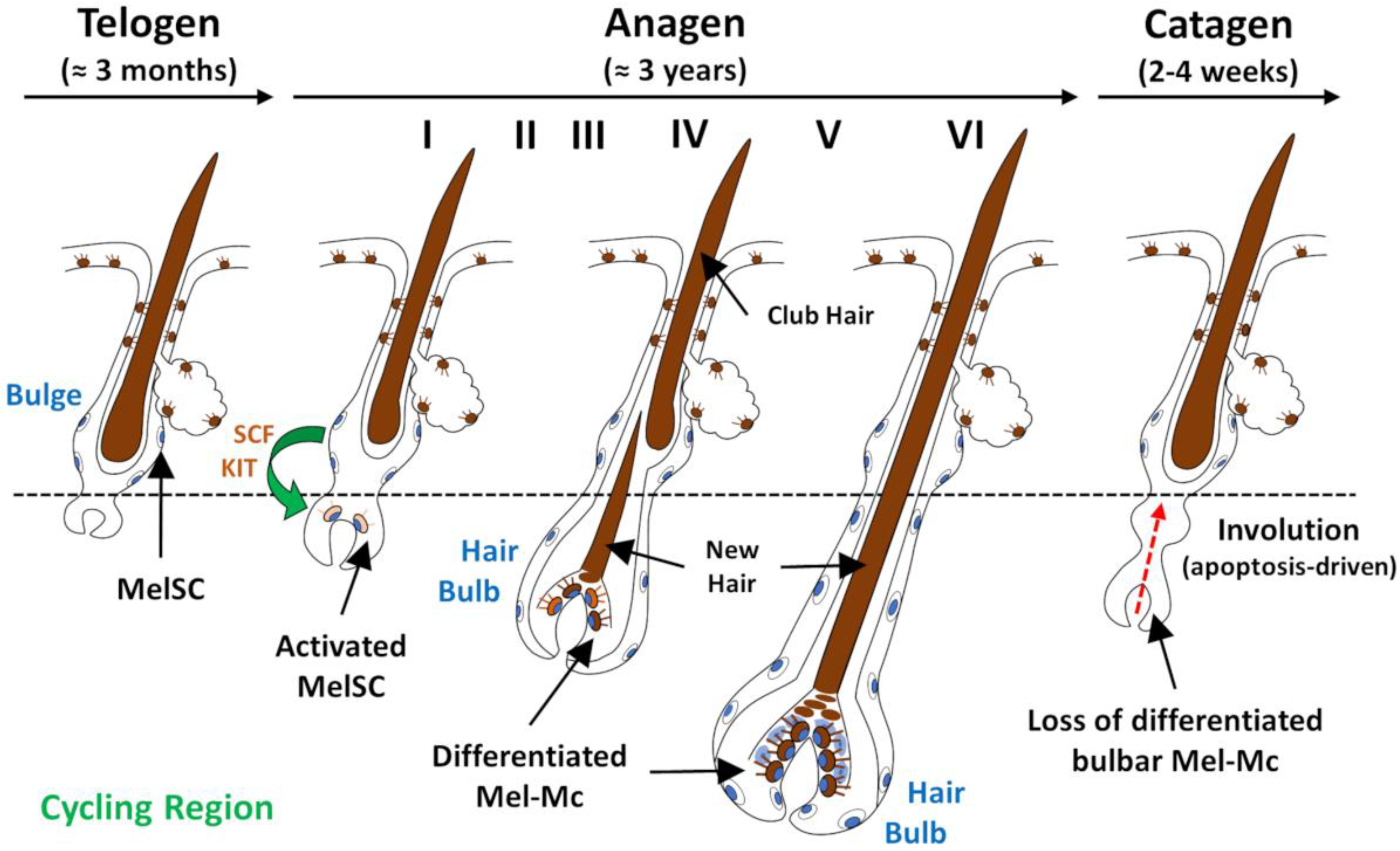{kind=link}
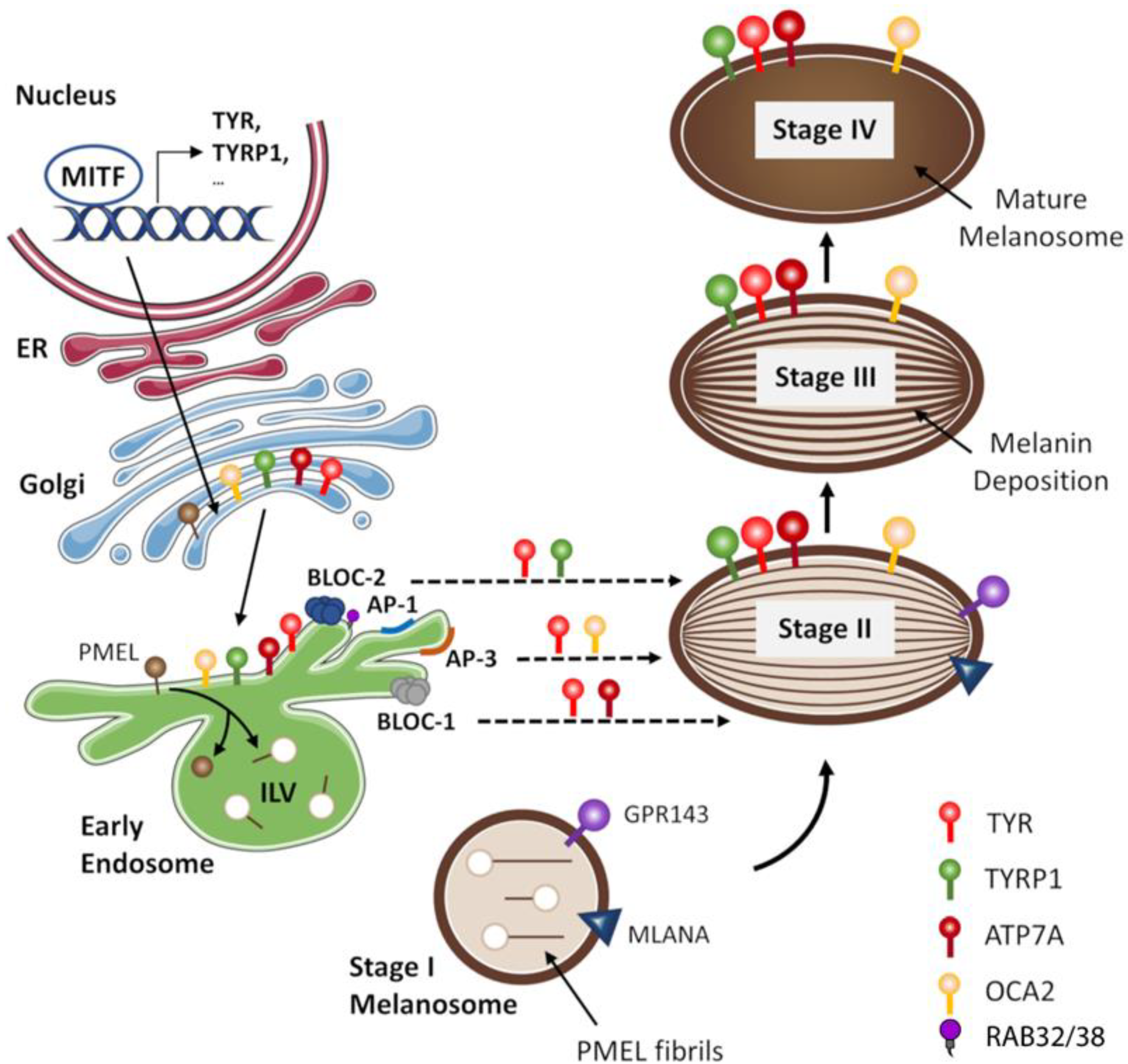{kind=link}
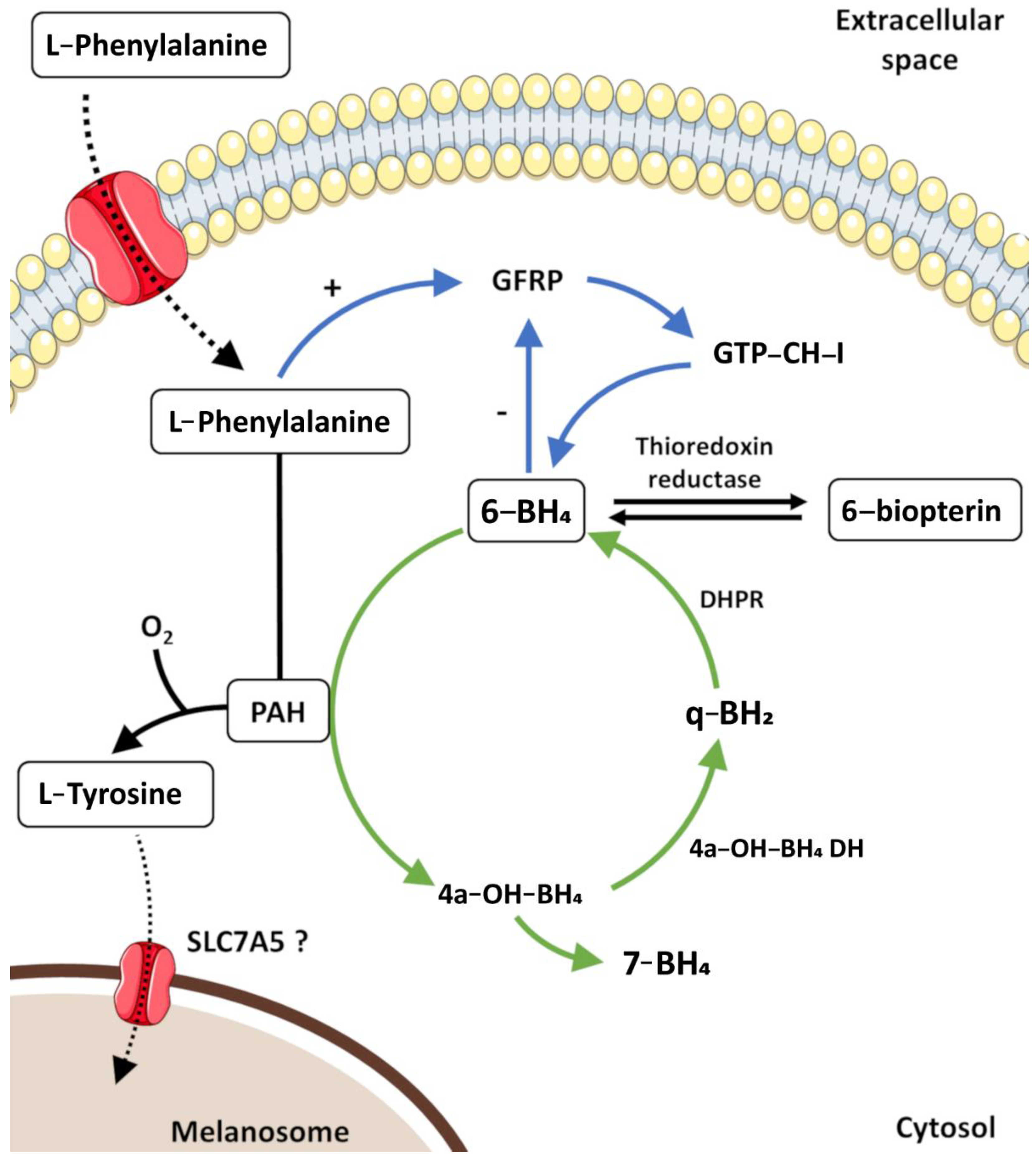{kind=link}
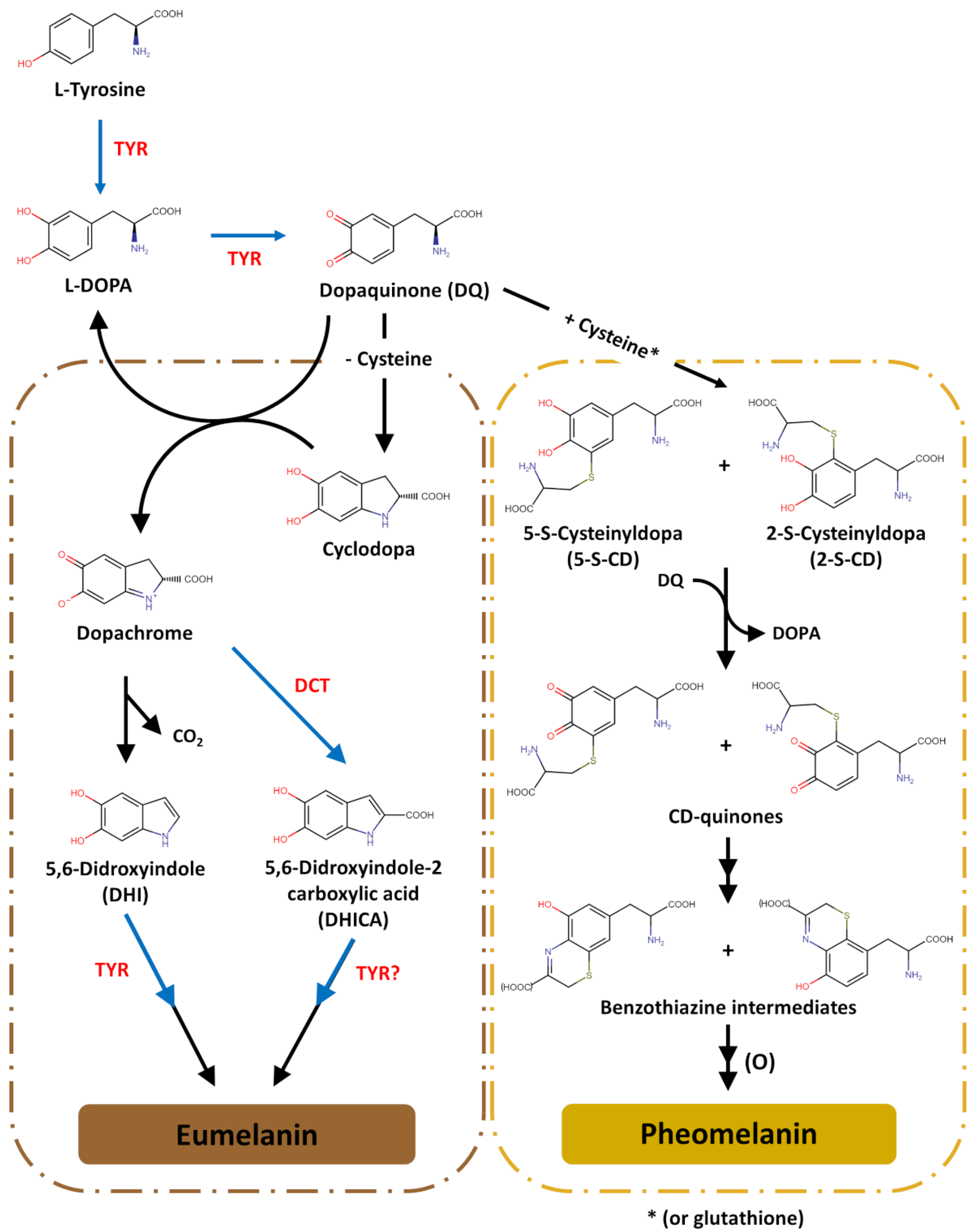{kind=link}
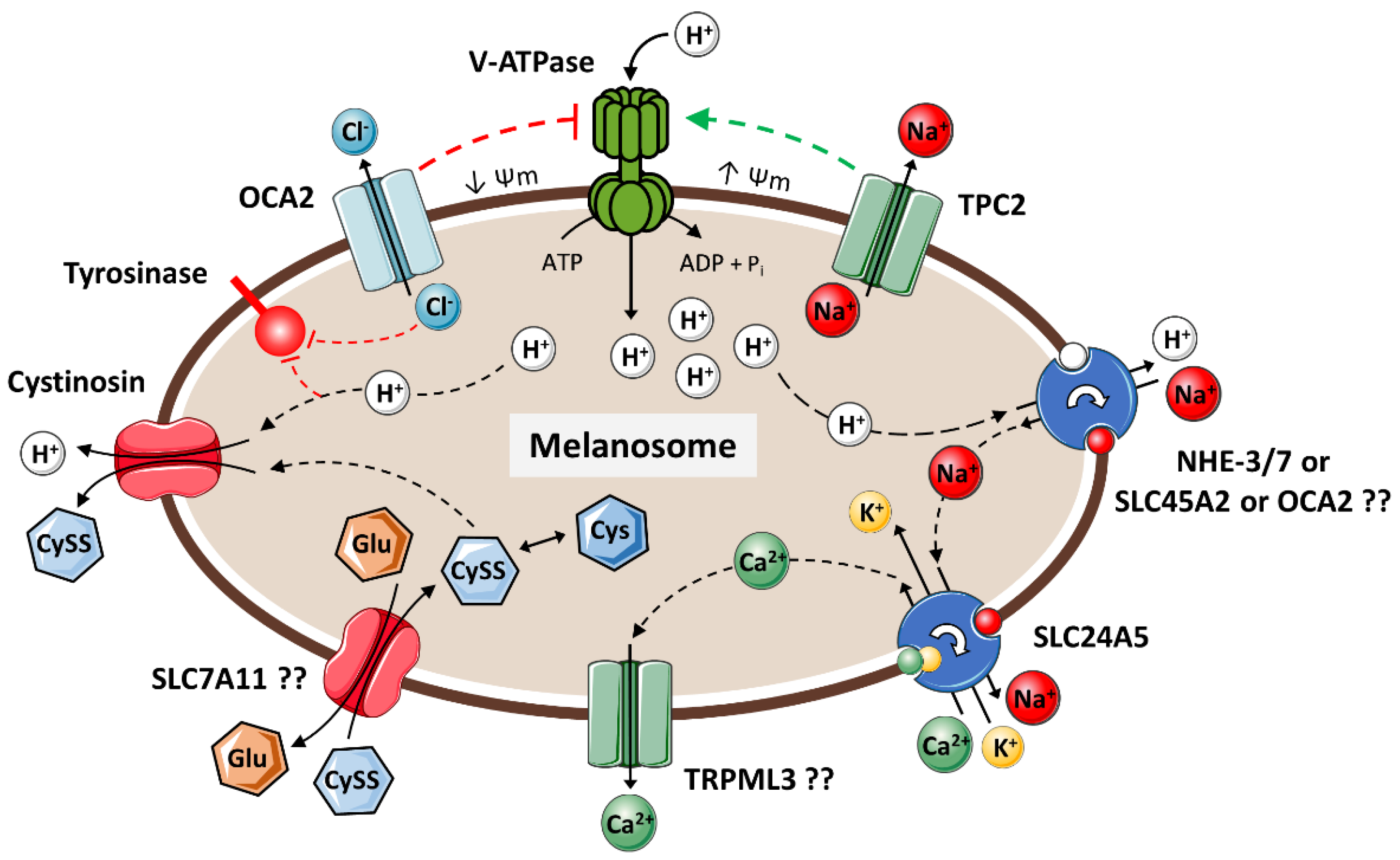{kind=link}
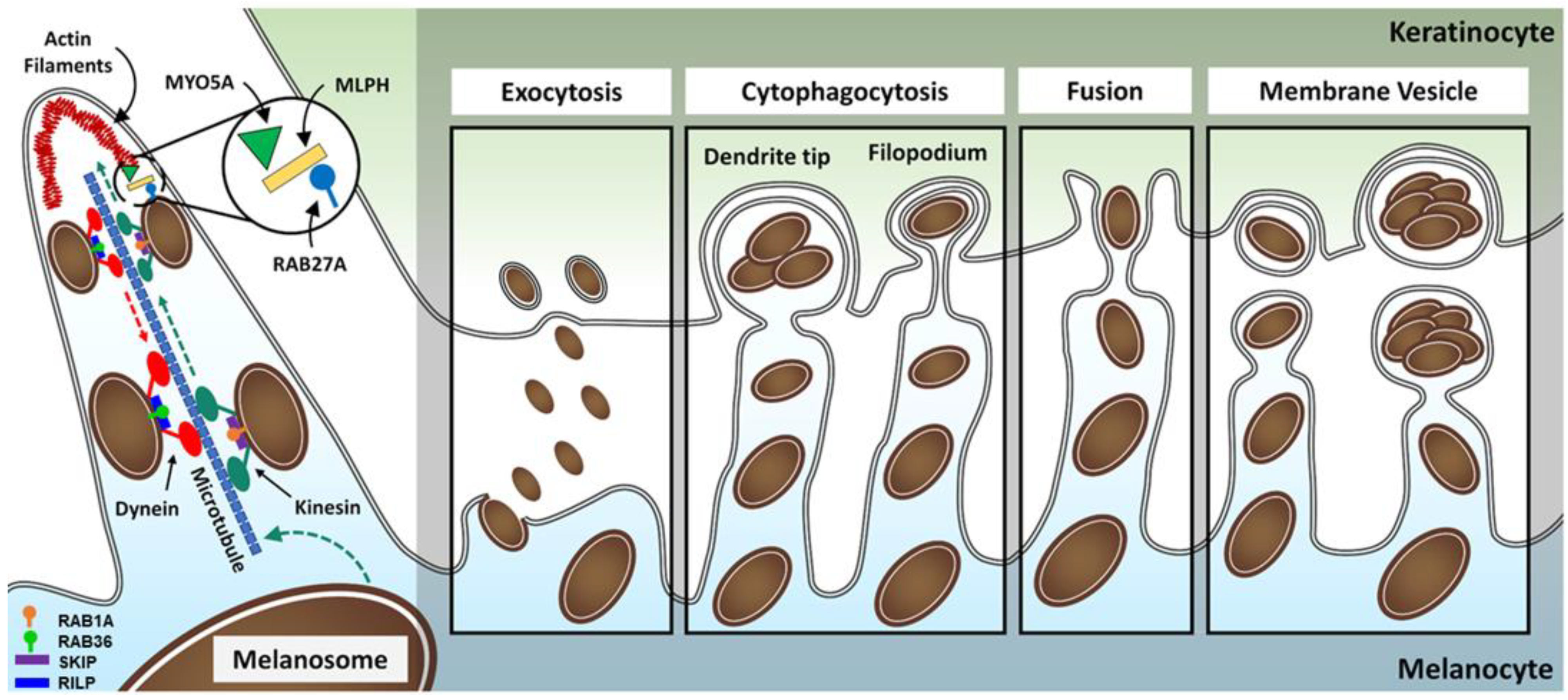{kind=link}
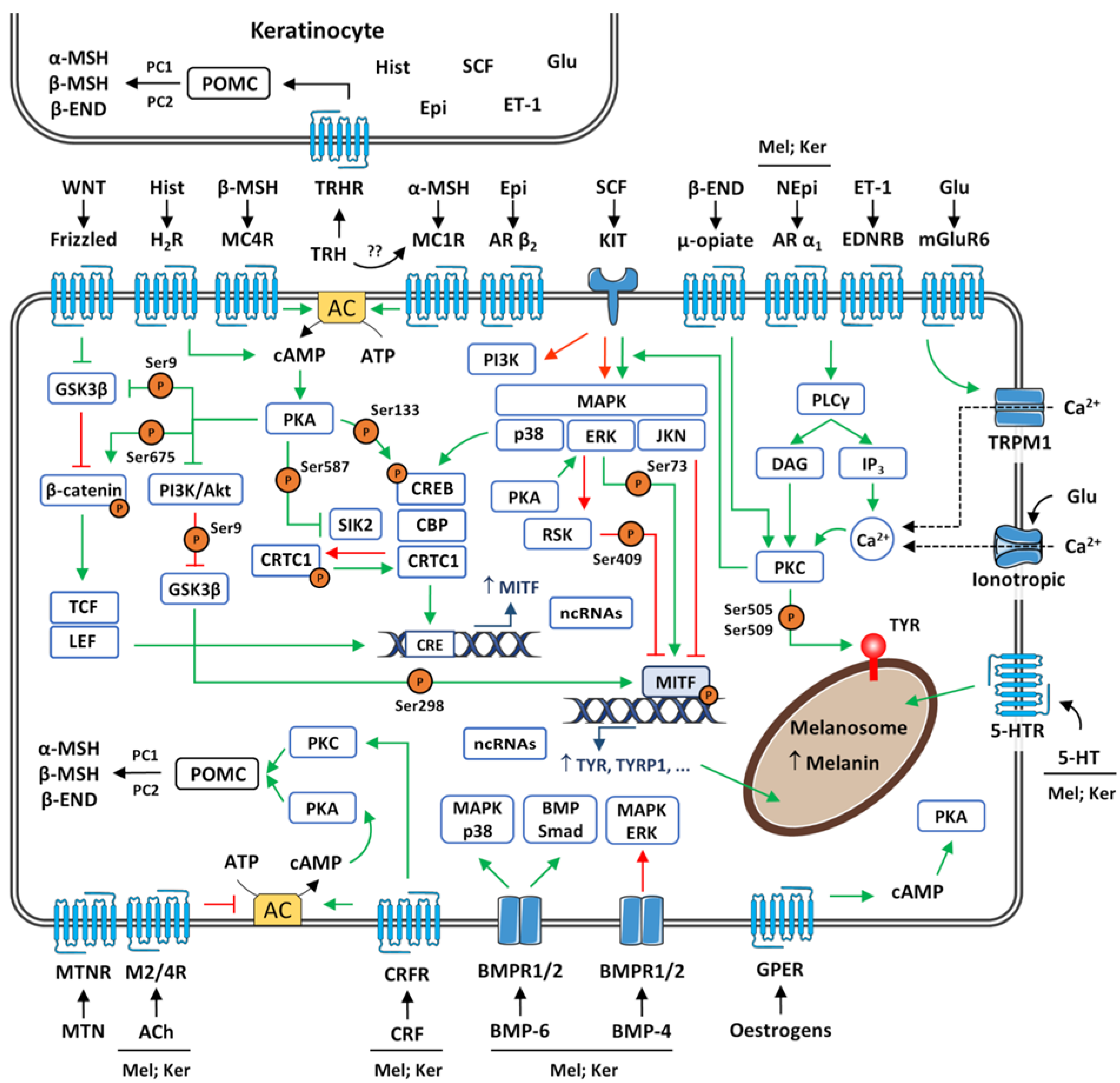{kind=link}
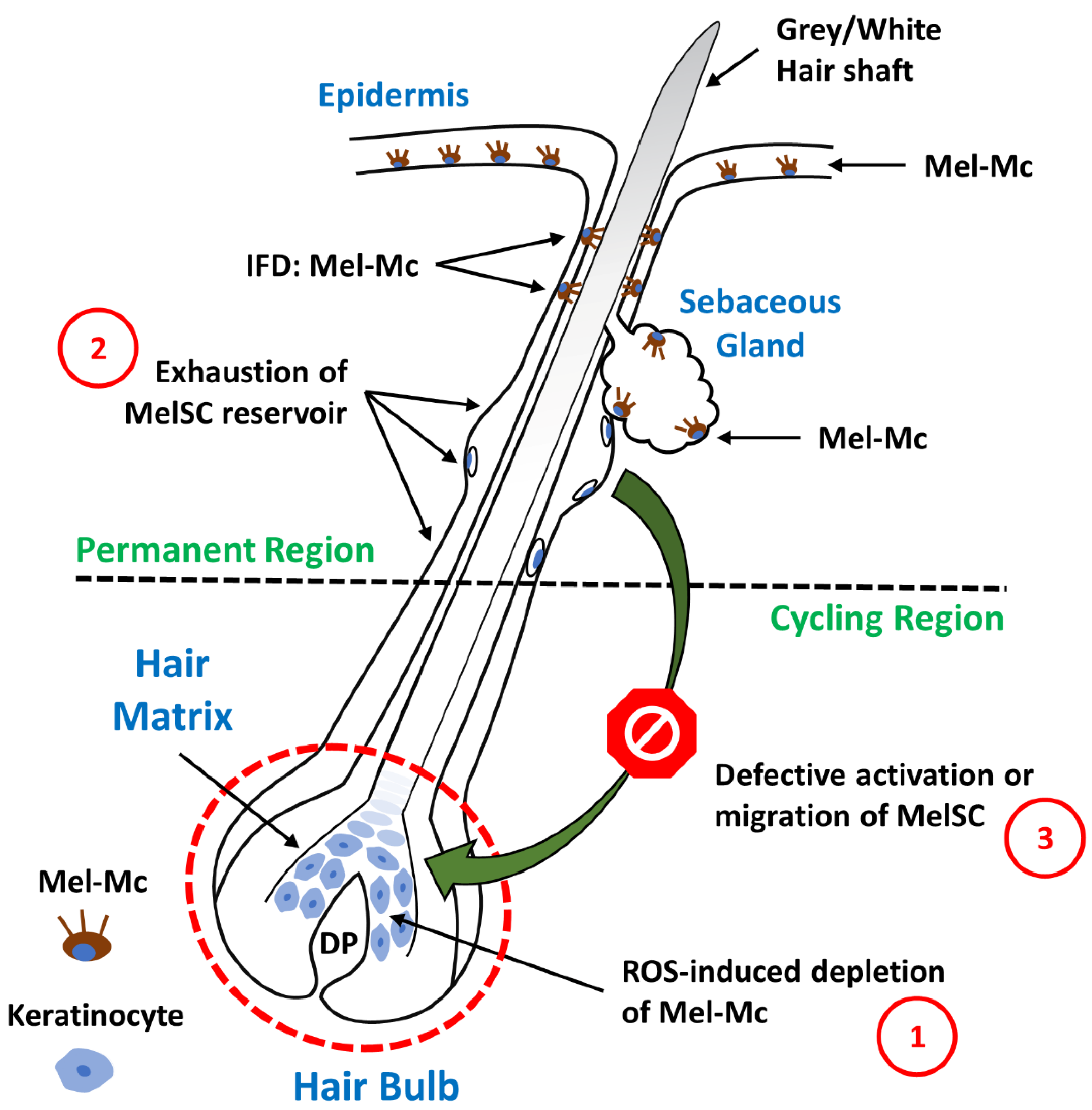{kind=link}
| Symbol | Gene Name | Protein Function | Reference(s) |
|---|---|---|---|
| Melanosomes Biogenesis | |||
| PMEL | Premelanosome protein | Amyloid fibril essential for melanosome biogenesis | [69] |
| MLANA | Melan-a | Regulation of PMEL expression | [69] |
| GPR143 | G protein-coupled receptor 143 | Regulation of melanosome maturation | [70] |
| Biochemical Synthesis of Melanin | |||
| TYR | Tyrosinase | Hydroxylation of L-tyrosine to L-DOPAOxidative polymerization of DHI | [71,72,73,74] |
| TYRP1 | Tyrosinase related protein 1 | Stabilization of tyrosinase | [73,75] |
| DCT | Dopachrome tautomerase | Tautomerization of dopachrome | [75] |
| Regulation of Melanosomal pH | |||
| SLC24A5 | Solute carrier family 24 member 5 | Na+/H+ exchanger | [76] |
| SLC45A2 | Solute carrier family 45 member 2 | K+-dependent Na+/Ca2+ exchanger | [77] |
| Melanosome Transport | |||
| RAB27A | RAB27A, member RAS oncogene family | Small GTPase able to bind to a variety of effector proteins; regulates melanosome trafficking in melanocytes | [78] |
| Regulation of Melanogenesis | |||
| MC1R | Melanocortin 1 receptor | Mediates melanocortin signalling | [79] |
| KIT | KIT proto-oncogene, receptor tyrosine kinase | Mediates stem cell factor signalling | [76] |
| EDNRB | Endothelin receptor type B | Mediates endothelin-1 signalling | [80] |
| Drug | Reason for Use | Hair Change | Reference(s) |
|---|---|---|---|
| α-Interferon | Melanoma | Lightening | [487] |
| Acitretin | Psoriasis | Repigmentation | [488,489] |
| Atezolizumab | Lung cancer | Repigmentation | [490] |
| Chloroquine | Malaria prophylaxis | Lightening | [491] |
| Dermatomyositis | [492] | ||
| Cisplatin | Metastatic germ cell neoplasm of the testis | Darkening | [493] |
| Lightening | |||
| Cyclosporin | Psoriasis | Darkening | [494] |
| Dabrafenib | Metastatic melanoma | Repigmentation | [495] |
| Dasatinib | Chronic myeloid leukaemia | Lightening | [496] |
| Defibrotide | Deep venous thrombosis | Darkening | [497] |
| Dupilumab | Atopic dermatitis | Repigmentation | [498] |
| Erlotinib | Lung adenocarcinoma | Repigmentation | [499] |
| Etretinate | Psoriasis | Lightening | [500] |
| Repigmentation | [501] | ||
| Pityriasis rubra pilaris | [502] | ||
| Hydroxychloroquine | Discoid lupus erythematosus | Lightening | [503] |
| Imatinib mesylate | Gastrointestinal stromal tumour | Lightening | [504] |
| Chronic myeloid leukaemia | [505] | ||
| L-Thyroxine | Hypothyroidism | Repigmentation | [506] |
| Latanoprost | Open-angle glaucoma | Repigmentation | [507] |
| Lenalidomide | Multiple myeloma | Repigmentation | [508] |
| Levodopa | Parkinson’s disease | Repigmentation | [509] |
| Nivolumab | Lung cancer | Repigmentation | [490] |
| Para-aminobenzoic acid | Lymphoblastoma cutis | Darkening | [510] |
| Dermatomyositis | |||
| Dermatitis herpetiformis | |||
| Scaly erythroderma | |||
| Scleroderma | |||
| Pazopanib | Hürthle cell carcinoma | Lightening | [511] |
| Myofibroblastic sarcoma | [512] | ||
| Pembrolizumab | Lung cancer | Repigmentation | [490] |
| Prednisone | Bullous pemphigoid | Repigmentation | [513] |
| Selumetinib | Plexiform neurofibroma | Lightening | [514] |
| Sunitinib malate | Gastrointestinal stromal tumour | Lightening | [515] |
| Tamoxifen | Breast cancer | Repigmentation | [516] |
| Thalidomide | Multiple myeloma | Repigmentation | [517] |
| Triptorelin | Precocious puberty | Greying | [518] |
| Ustekinumab | Psoriasis vulgaris | Repigmentation | [519] |
| Valproic acid | Seizures | Lightening | [520] |
| Vemurafenib | Metastatic melanoma | Repigmentation | [495] |
| Verapamil | Hypertension | Repigmentation | [521] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, B.; Cavaco-Paulo, A.; Matamá, T. A Comprehensive Review of Mammalian Pigmentation: Paving the Way for Innovative Hair Colour-Changing Cosmetics. Biology 2023, 12, 290. https://doi.org/10.3390/biology12020290
Fernandes B, Cavaco-Paulo A, Matamá T. A Comprehensive Review of Mammalian Pigmentation: Paving the Way for Innovative Hair Colour-Changing Cosmetics. Biology. 2023; 12(2):290. https://doi.org/10.3390/biology12020290
Chicago/Turabian StyleFernandes, Bruno, Artur Cavaco-Paulo, and Teresa Matamá. 2023. "A Comprehensive Review of Mammalian Pigmentation: Paving the Way for Innovative Hair Colour-Changing Cosmetics" Biology 12, no. 2: 290. https://doi.org/10.3390/biology12020290