

HN1 Is Enriched in the S-Phase, Phosphorylated in Mitosis, and Contributes to Cyclin B1 Degradation in Prostate Cancer Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mammalian Cell Culture
2.2. Chemicals and Antibodies
2.3. Cell Cycle Synchronizations
2.4. Plasmid DNA and siRNA Transfections
2.5. Stable Cell Line Generation
2.6. Protein Isolation, Immunoblotting, and Immunoprecipitations
2.7. Sub-Cellular Fractionations
2.8. Immunofluorescence (IF) Labeling and Microscopy
2.9. Flow Cytometry Analysis
2.10. Statistical Analyses
3. Results
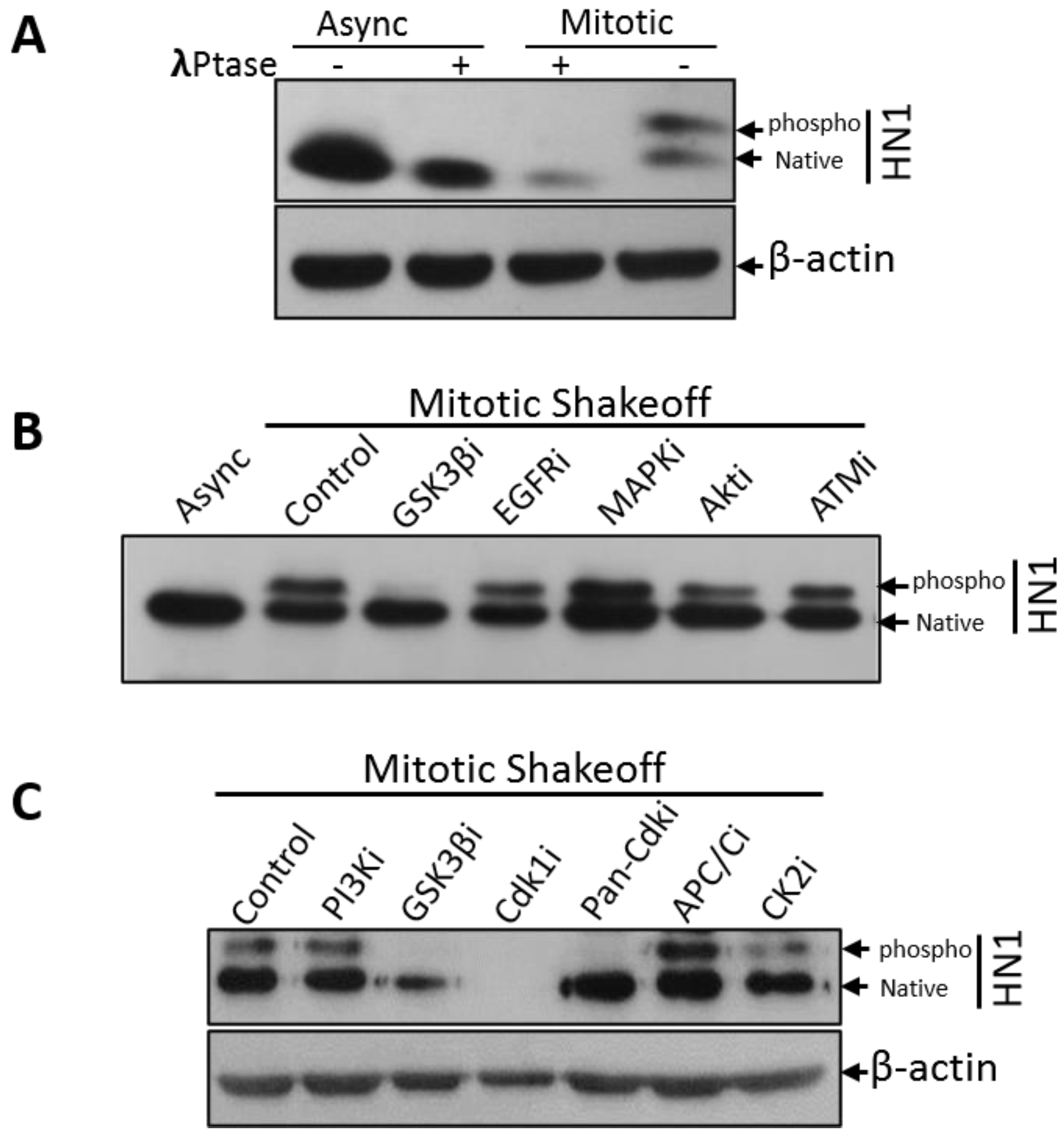
3.1. HN1 Protein Levels Fluctuate during the Cell Cycle with a Specific Phosphorylated Form in Mitosis
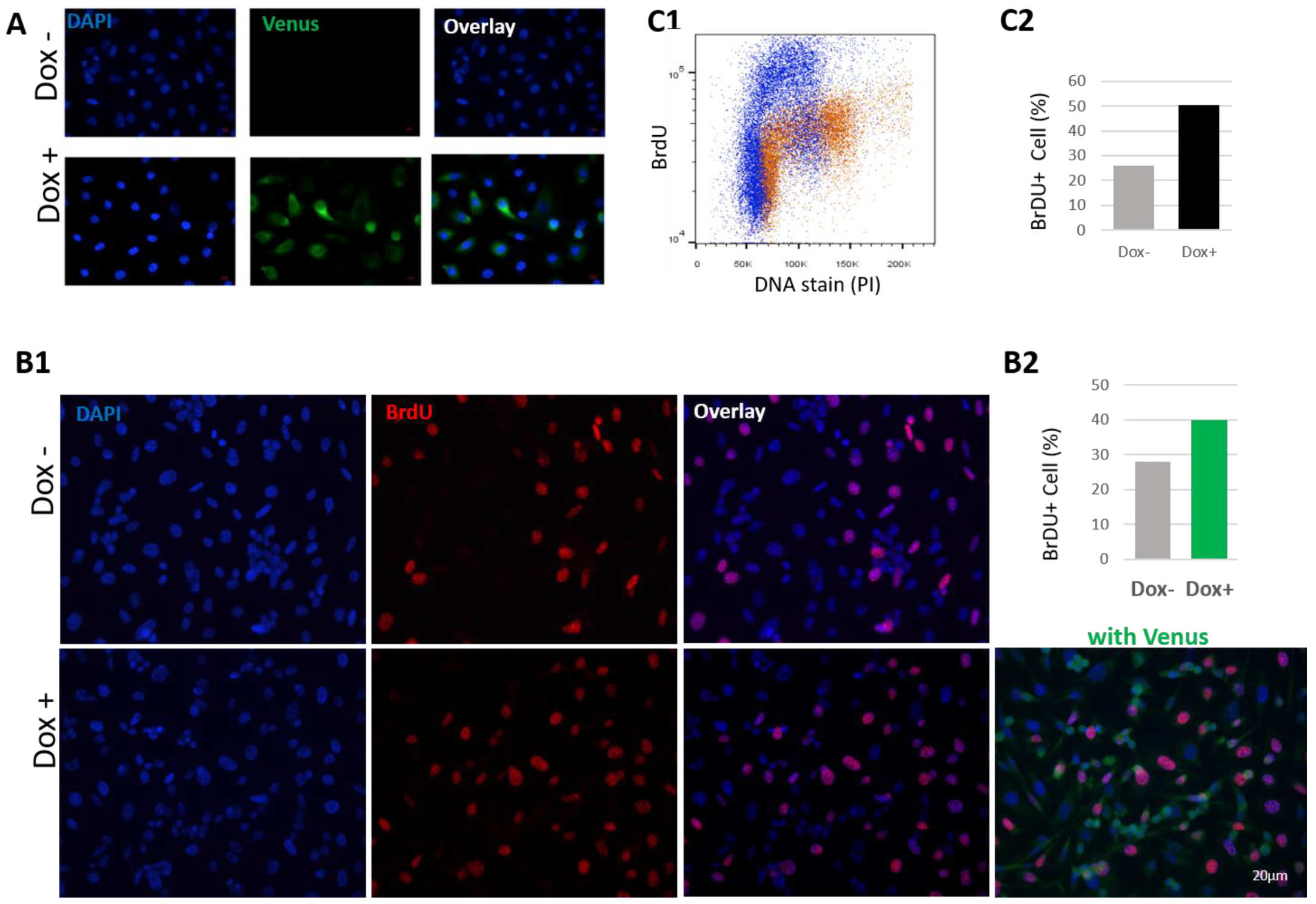
3.2. HN1 Overexpression Perturbs Cell Cycle Dynamics in Synchronized Cells
3.3. HN1 Associates with Cell Cycle Regulators and Modulates Cyclin B1 Levels
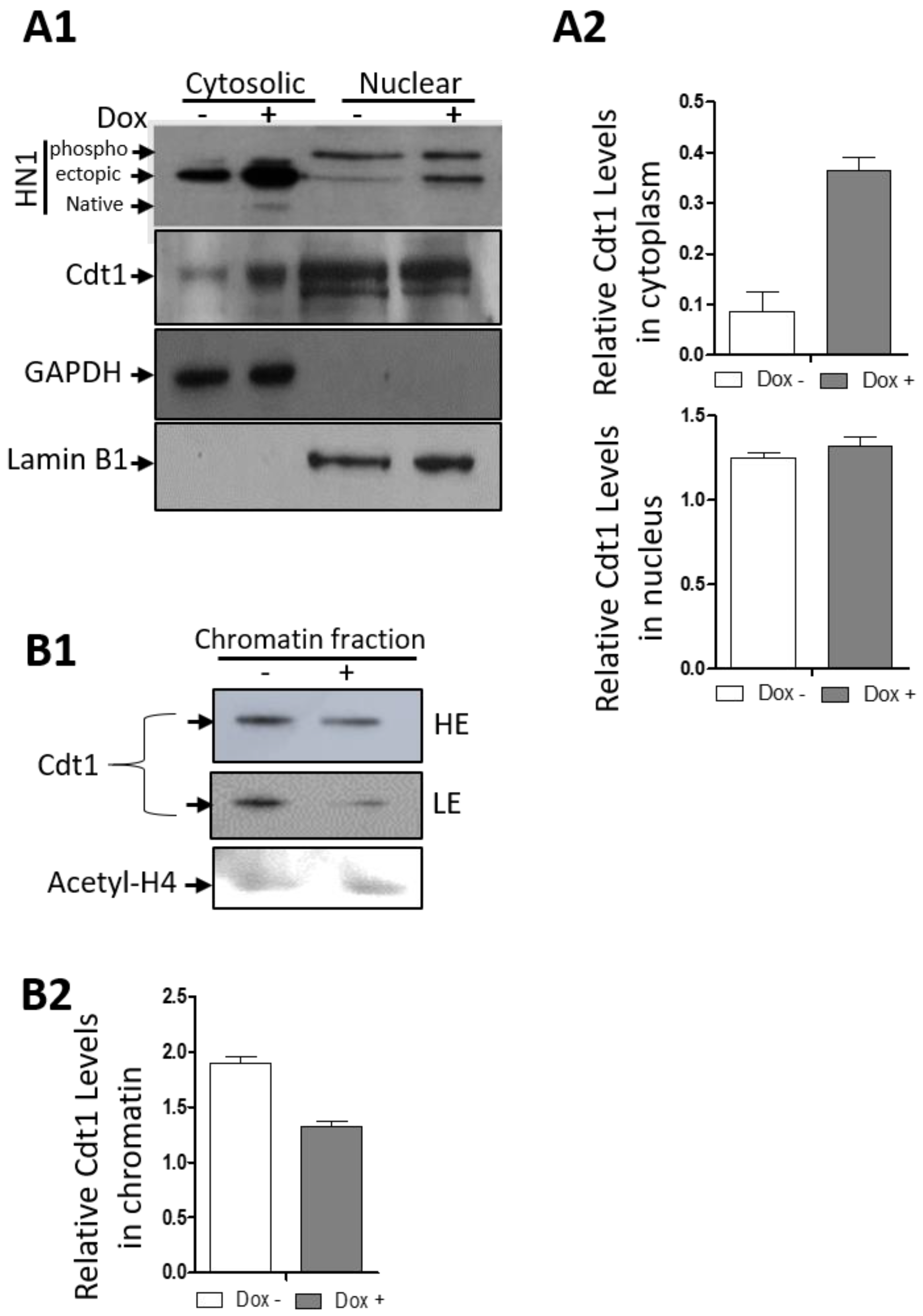
3.4. HN1 Is Directly Involved in The Regulation of S-Phase Dynamics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pernar, C.H.; Ebot, E.M.; Wilson, K.M.; Mucci, L.A. The Epidemiology of Prostate Cancer. Cold Spring Harb. Perspect. Med. 2018, 8, a030361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Huang, H.; Liu, A.; Li, J.; Liu, C.; Sun, B.; Chen, L.; Gao, Y.; Xu, D.; Su, C. Cell Division Cycle 20 (CDC20) Drives Prostate Cancer Progression via Stabilization of β-Catenin in Cancer Stem-like Cells. EBioMedicine 2019, 42, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Javed, A.; Malagraba, G.; Yarmohammadi, M.; Perelló-Reus, C.M.; Barceló, C.; Rubio-Tomás, T. Therapeutic Potential of Mitotic Kinases’ Inhibitors in Cancers of the Gastrointestinal System. Future Pharmacol. 2022, 2, 214–237. [Google Scholar] [CrossRef]
- Galbraith, M.D.; Bender, H.; Espinosa, J.M. Therapeutic Targeting of Transcriptional Cyclin-Dependent Kinases. Transcription 2019, 10, 118–136. [Google Scholar] [CrossRef] [PubMed]
- Mahdessian, D.; Cesnik, A.J.; Gnann, C.; Danielsson, F.; Stenström, L.; Arif, M.; Zhang, C.; Le, T.; Johansson, F.; Shutten, R.; et al. Spatiotemporal Dissection of the Cell Cycle with Single-Cell Proteogenomics. Nature 2021, 590, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, K.M.; Luo, D.; Liu, C.; Shaw, G.; Warrington, K.H.; Qiu, J.; Yachnis, A.T.; Harrison, J.K. Hematopoietic- and Neurologic-Expressed Sequence 1 Expression in the Murine GL261 and High-Grade Human Gliomas. Pathol. Oncol. Res. 2009, 15, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Bateman, N.W.; Teng, P.N.; Hope, E.; Hood, B.L.; Oliver, J.; Ao, W.; Zhou, M.; Wang, G.; Tommarello, D.; Wilson, K.; et al. Jupiter Microtubule-Associated Homolog 1 (JPT1): A Predictive and Pharmacodynamic Biomarker of Metformin Response in Endometrial Cancers. Cancer Med. 2020, 9, 1092–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanuik, T.L.; Wang, G.; Morozova, O.; Delaney, A.; Marra, M.A.; Sadar, M.D. LNCaP Atlas: Gene Expression Associated with in Vivo Progression to Castration-Recurrent Prostate Cancer. BMC Med. Genom. 2010, 3, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varisli, L.; Gonen-Korkmaz, C.; Debelec-Butuner, B.; Erbaykent-Tepedelen, B.; Muhammed, H.S.; Bogurcu, N.; Saatcioglu, F.; Korkmaz, K.S. Ubiquitously Expressed Hematological and Neurological Expressed 1 Downregulates Akt-Mediated GSK3β Signaling, and Its Knockdown Results in Deregulated G2/M Transition in Prostate Cells. DNA Cell Biol. 2011, 30, 419–429. [Google Scholar] [CrossRef]
- Varisli, L.; Gonen-Korkmaz, C.; Syed, H.M.; Bogurcu, N.; Debelec-Butuner, B.; Erbaykent-Tepedelen, B.; Korkmaz, K.S. Androgen Regulated HN1 Leads Proteosomal Degradation of Androgen Receptor (AR) and Negatively Influences AR Mediated Transactivation in Prostate Cells. Mol. Cell. Endocrinol. 2012, 350, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Varisli, L.; Ozturk, B.E.; Akyuz, G.K.; Korkmaz, K.S. HN1 Negatively Influences the β-Catenin/E-Cadherin Interaction, and Contributes to Migration in Prostate Cells. J. Cell. Biochem. 2015, 116, 170–178. [Google Scholar] [CrossRef]
- Testa, U.; Castelli, G.; Pelosi, E. Cellular and Molecular Mechanisms Underlying Prostate Cancer Development: Therapeutic Implications. Medicines 2019, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Varisli, L.; Javed, A.; Ozturk, B.E.; Akyuz, G.K.; Takir, G.; Roumelioti, F.-M.; Gagos, S.; Yorukoglu, K.; Korkmaz, K.S. HN1 Interacts with γ-Tubulin to Regulate Centrosomes in Advanced Prostate Cancer Cells. Cell Cycle 2021, 20, 1723–1744. [Google Scholar] [CrossRef]
- Schrock, M.S.; Stromberg, B.R.; Scarberry, L.; Summers, M.K. APC/C Ubiquitin Ligase: Functions and Mechanisms in Tumorigenesis. Semin. Cancer Biol. 2020, 67, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Clute, P.; Pines, J. Temporal and Spatial Control of Cyclin B1 Destruction in Metaphase. Nat. Cell Biol. 1999, 1, 82–87. [Google Scholar] [CrossRef]
- Van Zon, W.; Ogink, J.; Ter Riet, B.; Medema, R.H.; Te Riele, H.; Wolthuis, R.M.F. The APC/C Recruits Cyclin B1-Cdk1-Cks in Prometaphase before D Box Recognition to Control Mitotic Exit. J. Cell Biol. 2010, 190, 587–602. [Google Scholar] [CrossRef]
- Haneke, K.; Schott, J.; Lindner, D.; Hollensen, A.K.; Damgaard, C.K.; Mongis, C.; Knop, M.; Palm, W.; Ruggieri, A.; Stoecklin, G. CDK1 Couples Proliferation with Protein Synthesis. J. Cell Biol. 2020, 219, 201906147. [Google Scholar] [CrossRef] [Green Version]
- Kalous, J.; Jansová, D.; Šušor, A. Role of Cyclin-Dependent Kinase 1 in Translational Regulation in the M-Phase. Cells 2020, 9, 1568. [Google Scholar] [CrossRef] [PubMed]
- Varisli, L. Decreased Expression of HN1 Sensitizes Prostate Cancer Cells to Apoptosis Induced by Docetaxel and 2-Methoxyestradiol. Ann. Clin. Lab. Sci. 2022, 52, 196–201. [Google Scholar]
- Murray, A.W. Recycling the Cell Cycle: Cyclins Revisited. Cell 2004, 116, 221–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara-Gonzalez, P.; Moyle, M.W.; Budrewicz, J.; Mendoza-Lopez, J.; Oegema, K.; Desai, A. The G2-to-M Transition Is Ensured by a Dual Mechanism That Protects Cyclin B from Degradation by Cdc20-Activated APC/C. Dev. Cell 2019, 51, 313–325.e10. [Google Scholar] [CrossRef]
- McCloy, R.A.; Rogers, S.; Caldon, C.E.; Lorca, T.; Castro, A.; Burgess, A. Partial Inhibition of Cdk1 in G2 Phase Overrides the SAC and Decouples Mitotic Events. Cell Cycle 2014, 13, 1400–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Deng, X. Cell Synchronization by Double Thymidine Block. BIO-PROTOCOL 2018, 8, e2994. [Google Scholar] [CrossRef] [PubMed]
- Zieve, G.W.; Turnbull, D.; Mullins, J.M.; McIntosh, J.R. Production of Large Numbers of Mitotic Mammalian Cells by Use of the Reversible Microtubule Inhibitor Nocodazole. Nocodazole Accumulated Mitotic Cells. Exp. Cell Res. 1980, 126, 397–405. [Google Scholar] [CrossRef]
- Langan, T.J.; Chou, R.C. Synchronization of Mammalian Cell Cultures by Serum Deprivation. Methods Mol. Biol. 2011, 761, 75–83. [Google Scholar] [CrossRef]
- Schorl, C.; Sedivy, J.M. Analysis of Cell Cycle Phases and Progression in Cultured Mammalian Cells. Methods 2007, 41, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; King, R.W. An APC/C Inhibitor Stabilizes Cyclin B1 by Prematurely Terminating Ubiquitination. Nat. Chem. Biol. 2012, 8, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.; Hsiao, J.Y.; Davey, N.E.; van Voorhis, V.A.; Foster, S.A.; Tang, C.; Morgan, D.O. Multiple Mechanisms Determine the Order of APC/C Substrate Degradation in Mitosis. J. Cell Biol. 2014, 207, 23–39. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhang, P. The Function of APC/CCdh1 in Cell Cycle and Beyond. Cell Div. 2009, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Dimova, N.V.; Hathaway, N.A.; Lee, B.H.; Kirkpatrick, D.S.; Berkowitz, M.L.; Gygi, S.P.; Finley, D.; King, R.W. APC/C-Mediated Multiple Monoubiquitylation Provides an Alternative Degradation Signal for Cyclin B1. Nat. Cell Biol. 2012, 14, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Su, T.T.; Sprenger, F.; DiGregorio, P.J.; Campbell, S.D.; O’Farrell, P.H. Exit from Mitosis in Drosophila Syncytial Embryos Requires Proteolysis and Cychn Degradation, and Is Associated with Localized Dephosphorylation. Genes Dev. 1998, 12, 1495–1503. [Google Scholar] [CrossRef] [Green Version]
- Cappell, S.D.; Mark, K.G.; Garbett, D.; Pack, L.R.; Rape, M.; Meyer, T. EMI1 Switches from Being a Substrate to an Inhibitor of APC/CCDH1 to Start the Cell Cycle. Nature 2018, 558, 313–317. [Google Scholar] [CrossRef]
- Tang, W.; Lai, Y.H.; Han, X.D.; Wong, P.M.C.; Peters, L.L.; Chui, D.H.K. Murine Hn1 on Chromosome 11 Is Expressed in Hemopoietic and Brain Tissues. Mamm. Genome 1997, 8, 695–696. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Wang, J.; Zhang, Y.; Zhong, C.; Ni, J.; Wang, L.; Guo, J.; Zhang, K.; Yu, L.; Zhao, S. Cloning, Expression and Subcellular Localization of HN1 and HN1L Genes, as Well as Characterization of Their Orthologs, Defining an Evolutionarily Conserved Gene Family. Gene 2004, 331, 115–123. [Google Scholar] [CrossRef]
- Laughlin, K.M.; Luo, D.; Liu, C.; Shaw, G.; Warrington, K.H.; Law, B.K.; Harrison, J.K. Hematopoietic- and Neurologic-Expressed Sequence 1 (Hn1) Depletion in B16.F10 Melanoma Cells Promotes a Differentiated Phenotype That Includes Increased Melanogenesis and Cell Cycle Arrest. Differentiation 2009, 78, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.J.; Sun, X.; Mao, Q.Q.; Jiang, X.Y.; Zhao, X.G.; Xu, W.J.; Zhong, L. Increased Expression of Hematological and Neurological Expressed 1 (HN1) Is Associated with a Poor Prognosis of Hepatocellular Carcinoma and Its Knockdown Inhibits Cell Growth and Migration Partly by down-Regulation of c-Met. Kaohsiung J. Med. Sci. 2020, 36, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Liu, Y.; Fang, T.; Zhu, J.; Wang, G.; Li, J. Hematological and Neurological Expressed 1 (HN1) Activates C-Myc Signaling by Inhibiting Ubiquitin-mediated Proteasomal Degradation of C-Myc in Hepatocellular Carcinoma. Cell Biol. Int. 2022. [Google Scholar] [CrossRef]
- Wenzel, E.S.; Singh, A.T.K. Cell-Cycle Checkpoints and Aneuploidy on the Path to Cancer. In Vivo 2018, 32, 1–5. [Google Scholar]
- Malumbres, M.; Barbacid, M. Cell Cycle, CDKs and Cancer: A Changing Paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef]
- Choi, H.J.; Fukui, M.; Zhu, B.T. Role of Cyclin B1/Cdc2 up-Regulation in the Development of Mitotic Prometaphase Arrest in Human Breast Cancer Cells Treated with Nocodazole. PLoS ONE 2011, 6, e24312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, L.A.; Camacho Reis, M.; Ciossani, G.; Huis in ‘t Veld, P.J.; Wohlgemuth, S.; Kops, G.J.; Musacchio, A.; Saurin, A.T. Cyclin B1 Scaffolds MAD 1 at the Kinetochore Corona to Activate the Mitotic Checkpoint. EMBO J. 2020, 39, e103180. [Google Scholar] [CrossRef]
- Martínez-Alonso, D.; Malumbres, M. Mammalian Cell Cycle Cyclins. Semin. Cell Dev. Biol. 2020, 107, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Müllers, E.; Cascales, H.S.; Jaiswal, H.; Saurin, A.T.; Lindqvist, A. Nuclear Translocation of Cyclin B1 Marks the Restriction Point for Terminal Cell Cycle Exit in G2 Phase. Cell Cycle 2014, 13, 2733–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, D.V.; Sramkoski, R.M.; Lam, M.; Stefan, T.; Jacobberger, J.W. Cyclin B1 Is Rate Limiting but Not Essential for Mitotic Entry and Progression in Mammalian Somatic Cells. Cell Cycle 2008, 7, 1285–1300. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wei, K.; Li, X.; Hu, R.; Qiu, J.; Zhang, Y.; Yao, W.; Zhang, C.; Zhu, C. Upregulation of Cdh1 Signaling in the Hippocampus Attenuates Brain Damage after Transient Global Cerebral Ischemia in Rats. Neurochem. Int. 2018, 112, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Bolaños, J.P.; Moreno, S. Cdh1/Hct1-APC Is Essential for the Survival of Postmitotic Neurons. J. Neurosci. 2005, 25, 8115–8121. [Google Scholar] [CrossRef] [Green Version]
- Shang, Z.F.; Tan, W.; Liu, X.D.; Yu, L.; Li, B.; Li, M.; Song, M.; Wang, Y.; Xiao, B.B.; Zhong, C.G.; et al. DNA-PKcs Negatively Regulates Cyclin B1 Protein Stability through Facilitating Its Ubiquitination Mediated by Cdh1-APC/C Pathway. Int. J. Biol. Sci. 2015, 11, 1026–1035. [Google Scholar] [CrossRef] [Green Version]
- Grant, G.D.; Kedziora, K.M.; Limas, J.C.; Cook, J.G.; Purvis, J.E. Accurate Delineation of Cell Cycle Phase Transitions in Living Cells with PIP-FUCCI. Cell Cycle 2018, 17, 2496–2516. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Hellwarth, P.B.; Randolph, L.N.; Sun, Y.; Xing, Y.; Zhu, W.; Lian, X.L.; Bao, X. Fluorescent Indicators for Continuous and Lineage-Specific Reporting of Cell-Cycle Phases in Human Pluripotent Stem Cells. Biotechnol. Bioeng. 2020, 117, 2177–2186. [Google Scholar] [CrossRef]
- Drpic, D.; Almeida, A.C.; Aguiar, P.; Renda, F.; Damas, J.; Lewin, H.A.; Larkin, D.M.; Khodjakov, A.; Maiato, H. Chromosome Segregation Is Biased by Kinetochore Size. Curr. Biol. 2018, 28, 1344–1356.e5. [Google Scholar] [CrossRef] [Green Version]
- Cairo, G.; Lacefield, S. Establishing Correct Kinetochore-Microtubule Attachments in Mitosis and Meiosis. Essays Biochem. 2020, 64, 277–287. [Google Scholar]
- Pu, J.; Wang, J.; Li, W.; Lu, Y.; Wu, X.; Long, X.; Luo, C.; Wei, H. Hsa_circ_0000092 Promotes Hepatocellular Carcinoma Progression through Up-Regulating HN1 Expression by Binding to MicroRNA-338-3p. J. Cell. Mol. Med. 2020, 22, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Fu, Y.; Yao, M.; Cui, X.; Zhao, Y.; Lu, X.; Li, Y.; Lin, Y.; He, S. The HN1/HMGB1 Axis Promotes the Proliferation and Metastasis of Hepatocellular Carcinoma and Attenuates the Chemosensitivity to Oxaliplatin. FEBS J. 2022, 289, 6400–6419. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Fang, Q.; Li, L.; Zhang, Y.; Xu, T.; Liu, Y.; Zheng, X.; Tan, Z.; Huang, P.; Ge, M. HN1 Promotes Tumor Growth and Metastasis of Anaplastic Thyroid Carcinoma by Interacting with STMN1. Cancer Lett. 2021, 501, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qiu, J.; Li, F.; Jiang, X.; Sun, X.; Zheng, L.; Zhang, W.; Li, H.; Wu, H.; Ouyang, Y.; et al. HN1 Promotes Tumor Associated Lymphangiogenesis and Lymph Node Metastasis via NF-ΚB Signaling Activation in Cervical Carcinoma. Biochem. Biophys. Res. Commun. 2020, 530, 87–94. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, B.; Lu, S.; Zhao, Y.; Liu, P. HN1 Contributes to Migration, Invasion, and Tumorigenesis of Breast Cancer by Enhancing MYC Activity. Mol. Cancer 2017, 16, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Wang, S.; Cen, C.; Peng, S.; Chen, Y.; Li, X.; Diao, N.; Li, Q.; Ma, L.; Han, P. Identification of Differentially Expressed Genes in Pancreatic Ductal Adenocarcinoma and Normal Pancreatic Tissues Based on Microarray Datasets. Mol. Med. Rep. 2019, 20, 1901–1914. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Chen, W.X.; Wu, Y.H.; Liang, H.F.; Zhang, B.X. MiR-132 Prohibits Proliferation, Invasion, Migration, and Metastasis in Breast Cancer by Targeting HN1. Biochem. Biophys. Res. Commun. 2014, 454, 109–114. [Google Scholar] [CrossRef]
- Xouri, G.; Squire, A.; Dimaki, M.; Geverts, B.; Verveer, P.J.; Taraviras, S.; Nishitani, H.; Houtsmuller, A.B.; Bastiaens, P.I.H.; Lygerou, Z. Cdt1 Associates Dynamically with Chromatin throughout G1 and Recruits Geminin onto Chromatin. EMBO J. 2007, 26, 1303–1314. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H. Regulation of Dna Replication Licensing and Re-Replication by Cdt1. Int. J. Mol. Sci. 2021, 22, 5195. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javed, A.; Özduman, G.; Varışlı, L.; Öztürk, B.E.; Korkmaz, K.S. HN1 Is Enriched in the S-Phase, Phosphorylated in Mitosis, and Contributes to Cyclin B1 Degradation in Prostate Cancer Cells. Biology 2023, 12, 189. https://doi.org/10.3390/biology12020189
Javed A, Özduman G, Varışlı L, Öztürk BE, Korkmaz KS. HN1 Is Enriched in the S-Phase, Phosphorylated in Mitosis, and Contributes to Cyclin B1 Degradation in Prostate Cancer Cells. Biology. 2023; 12(2):189. https://doi.org/10.3390/biology12020189
Chicago/Turabian StyleJaved, Aadil, Gülseren Özduman, Lokman Varışlı, Bilge Esin Öztürk, and Kemal Sami Korkmaz. 2023. "HN1 Is Enriched in the S-Phase, Phosphorylated in Mitosis, and Contributes to Cyclin B1 Degradation in Prostate Cancer Cells" Biology 12, no. 2: 189. https://doi.org/10.3390/biology12020189