
Single-Cell Measurements and Modeling and Computation of Decision-Making Errors in a Molecular Signaling System with Two Output Molecules

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Using the Likelihood Ratio to Compute the Optimal Decision Thresholds and the Decision Error Probabilities for the TNF—NFκB/ATF-2 System
2.1.1. Univariate Decision Analysis
2.1.2. Bivariate Decision Analysis
2.2. Using the Discriminant Function to Compute the Decision Error Probabilities for the TNF—NFκB/ATF-2 System
3. Results and Discussion
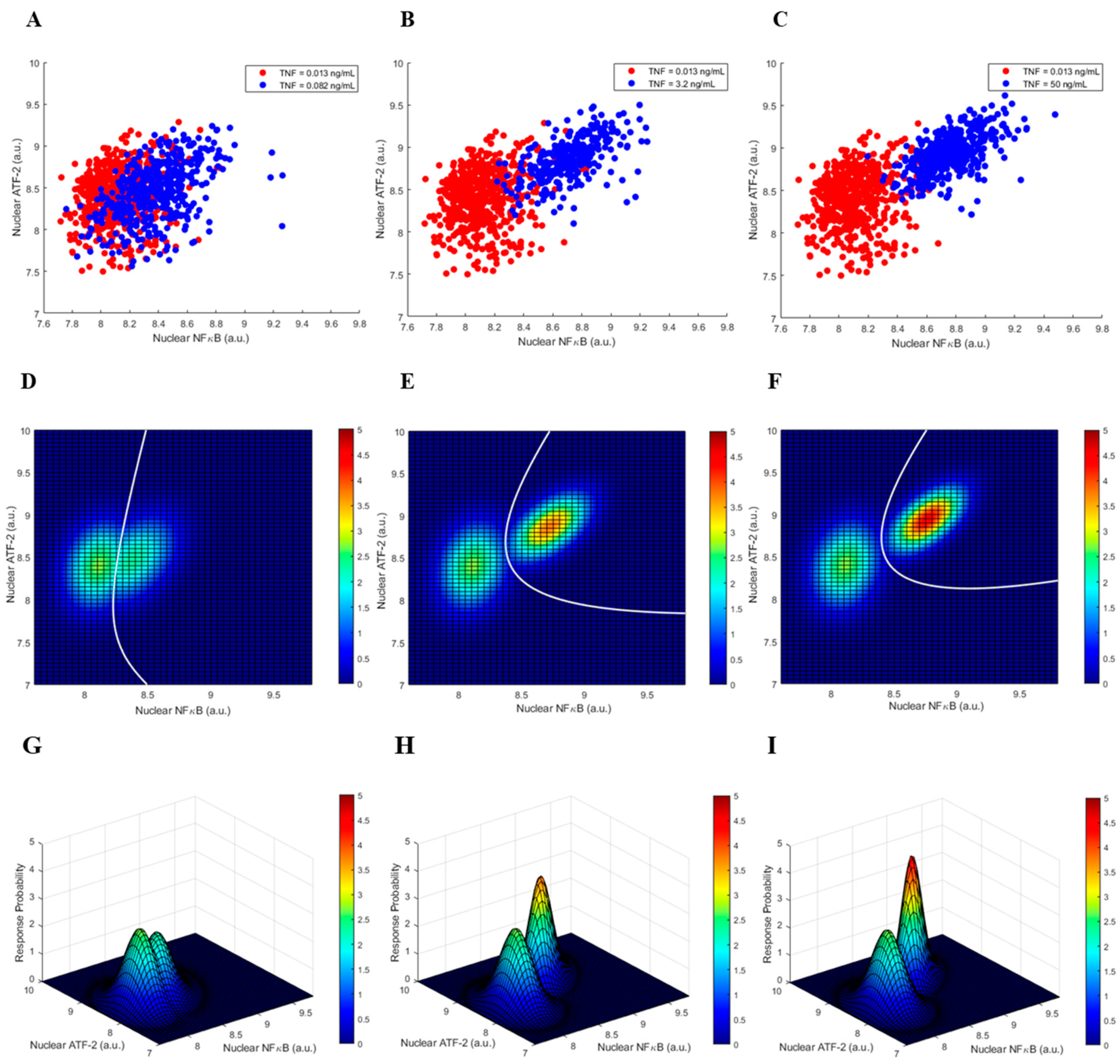
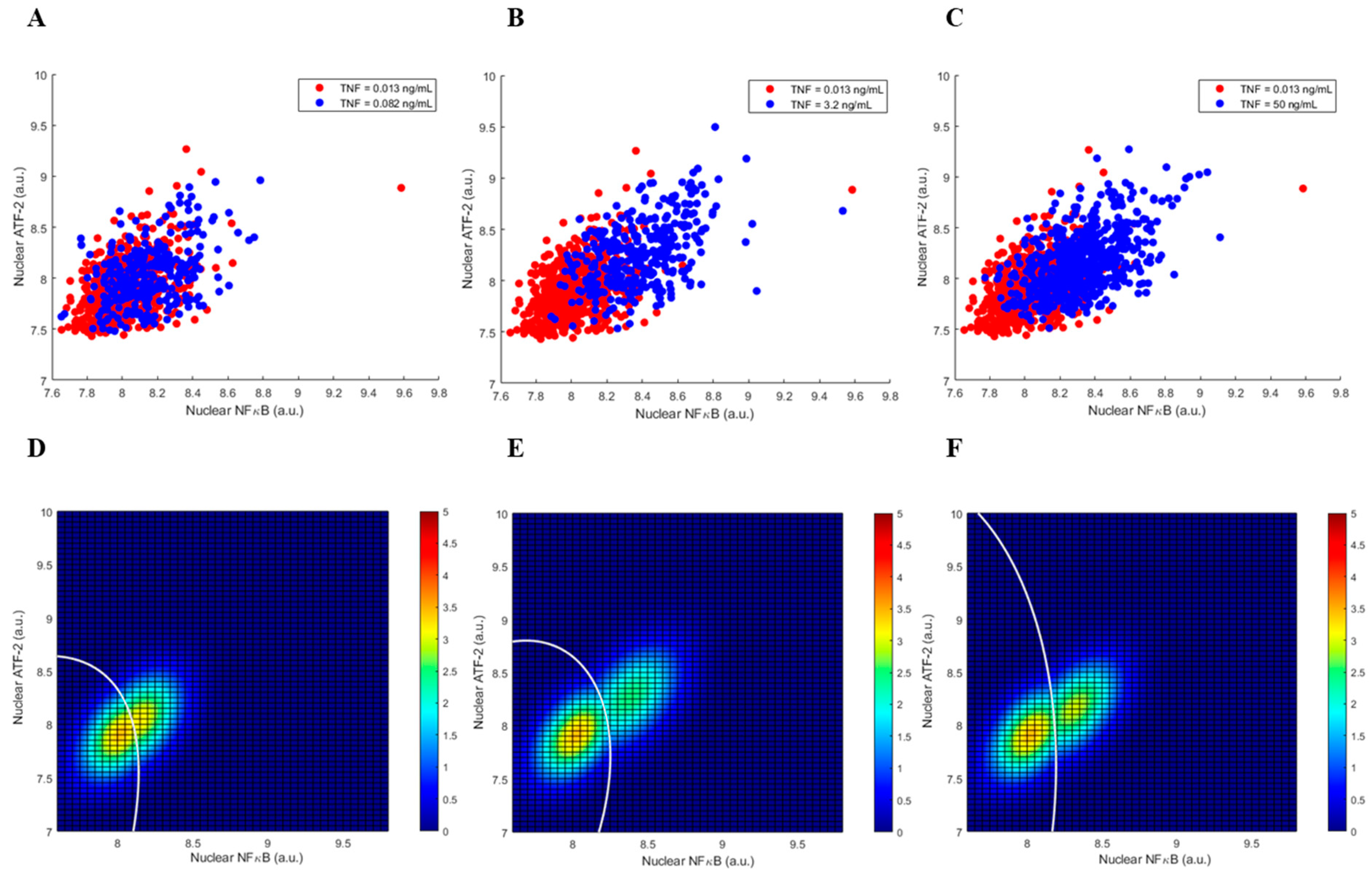
3.1. Single Cell Data of the Two-Output TNF—NFκB/ATF-2 System
3.2. Graphical Representation of the Two-Output System Data and the Decision Thresholds
3.3. Decision Error Probabilities of the Two-Output System
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kolitz, S.E.; Lauffenburger, D.A. Measurement and modeling of signaling at the single-cell level. Biochemistry 2012, 51, 7433–7443. [Google Scholar] [CrossRef] [PubMed]
- Tay, S.; Hughey, J.J.; Lee, T.K.; Lipniacki, T.; Quake, S.R.; Covert, M.W. Single-cell NF-κB dynamics reveal digital activation and analogue information processing. Nature 2010, 466, 267–271. [Google Scholar] [CrossRef]
- Maity, A.; Wollman, R. Information transmission from NFkB signaling dynamics to gene expression. PLoS Comput. Biol. 2020, 16, e1008011. [Google Scholar] [CrossRef] [PubMed]
- Talia, S.D.; Skotheim, J.M.; Bean, J.M.; Siggia, E.D.; Cross, F.R. The effects of molecular noise and size control on variability in the budding yeast cell cycle. Nature 2007, 448, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Cheong, R.; Rhee, A.; Wang, C.J.; Nemenman, I.; Levchenko, A. Information transduction capacity of noisy biochemical signaling networks. Science 2011, 334, 354–358. [Google Scholar] [CrossRef]
- Ladbury, J.E.; Arold, S.T. Noise in cellular signaling pathways: Causes and effects. Trends Biochem. Sci. 2012, 37, 173–178. [Google Scholar] [CrossRef]
- Balazsi, G.; Van Oudenaarden, A.; Collins, J.J. Cellular decision making and biological noise: From microbes to mammals. Cell 2011, 144, 910–925. [Google Scholar] [CrossRef]
- Weitz, J.S.; Mileyko, Y.; Joh, R.I.; Voit, E.O. Collective decision making in bacterial viruses. Biophys. J. 2008, 95, 2673–2680. [Google Scholar] [CrossRef]
- Zeng, L.; Skinner, S.O.; Zong, C.; Sippy, J.; Feiss, M.; Golding, I. Decision making at a subcellular level determines the outcome of bacteriophage infection. Cell 2010, 141, 682–691. [Google Scholar] [CrossRef]
- Moris, N.; Pina, C.; Arias, A.M. Transition states and cell fate decisions in epigenetic landscapes. Nat. Rev. Genet. 2016, 17, 693–703. [Google Scholar] [CrossRef]
- Matson, J.P.; Cook, J.G. Cell cycle proliferation decisions: The impact of single cell analyses. FEBS J. 2017, 284, 362–375. [Google Scholar] [CrossRef]
- Rodrigo, G. Insights about collective decision-making at the genetic level. Biophys. Rev. 2020, 12, 19–24. [Google Scholar] [CrossRef]
- Habibi, I.; Cheong, R.; Lipniacki, T.; Levchenko, A.; Emamian, E.S.; Abdi, A. Computation and measurement of cell decision making errors using single cell data. PLoS Comput. Biol. 2017, 13, e1005436. [Google Scholar] [CrossRef] [PubMed]
- Levchenko, A. Genetic diseases: How the noise fits in. Curr. Biol. 2023, 33, 228–230. [Google Scholar] [CrossRef]
- Emadi, A.; Ozen, M.; Abdi, A. A hybrid model to study how late long-term potentiation is affected by faulty molecules in an intraneuronal signaling network regulating transcription factor CREB. Integr. Biol. 2022, 14, 111–125. [Google Scholar] [CrossRef]
- Micheau, O.; Tschopp, J. Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes. Cell 2003, 114, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Oliver Metzig, M.; Tang, Y.; Mitchell, S.; Taylor, B.; Foreman, R.; Wollman, R.; Hoffmann, A. An incoherent feedforward loop interprets NFκB/RelA dynamics to determine TNF-induced necroptosis decisions. Mol. Syst. Biol. 2020, 16, e9677. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.A. Viruses and the TNF-related cytokines, an evolving battle. Cytokine Growth Factor Rev. 2003, 14, 349–357. [Google Scholar] [CrossRef]
- Brenner, D.; Blaser, H.; Mak, T.W. Regulation of tumour necrosis factor signalling: Live or let die. Nat. Rev. Immunol. 2015, 15, 362–374. [Google Scholar] [CrossRef]
- Oyler-Yaniv, J.; Oyler-Yaniv, A.; Maltz, E.; Wollman, R. TNF controls a speed-accuracy tradeoff in the cell death decision to restrict viral spread. Nat. Commun. 2021, 12, 2992. [Google Scholar] [CrossRef]
- Hayden, M.S.; West, A.P.; Ghosh, S. NF-κB and the immune response. Oncogene 2006, 25, 6758–6780. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Natoli, G.; Gosh, G. Transcriptional regulation via the NF-κB signaling module. Oncogene 2006, 25, 6706–6716. [Google Scholar] [CrossRef]
- Hoffmann, A.; Baltimore, D. Circuitry of nuclear factor κB signaling. Immunol. Rev. 2006, 210, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Covert, M.W. High-throughput, single-cell NF-κB dynamics. Curr. Opin. Genet. Dev. 2010, 20, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Barnabei, L.; Laplantine, E.; Mbongo, W.; Rieux-Laucat, F.; Weil, R. NF-κB: At the Borders of Autoimmunity and Inflammation. Front. Immunol. 2021, 12, 716469. [Google Scholar] [CrossRef]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef]
- O’Dea, E.; Hoffmann, A. NF-κB signaling. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 107–115. [Google Scholar] [CrossRef]
- Hymowitz, S.G.; Wertz, I.E. A20: From ubiquitin editing to tumour suppression. Nat. Rev. Cancer 2010, 10, 332–341. [Google Scholar] [CrossRef]
- Chen, M.; Liu, Y.; Yang, Y.; Qiu, Y.; Wang, Z.; Li, X.; Zhang, W. Emerging roles of activating transcription factor (ATF) family members in tumourigenesis and immunity: Implications in cancer immunotherapy. Genes Dis. 2021, 9, 981–999. [Google Scholar] [CrossRef]
- Van Trees, H.L.; Bell, K.L.; Tian, Z. Detection, Estimation and Modulation Theory, Part I: Detection, Estimation, and Filtering Theory, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Kay, S.M. Fundamentals of Statistical Signal Processing: Detection Theory; PTR Prentice-Hall: Hoboken, NJ, USA, 1998. [Google Scholar]
- Papoulis, A. Probability, Random Variables, and Stochastic Processes, 3rd ed.; McGraw-Hill: New York, NY, USA, 1991. [Google Scholar]
- Duda, O.R.; Hart, P.E.; Stork, D.G. Pattern Classification; John Wiley & Sons: New York, NY, USA, 2001. [Google Scholar]
- Fukunaga, K. Introduction to Statistical Pattern Recognition, 2nd ed.; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Ding, C.; Peng, H. Minimum redundancy feature selection from microarray gene expression data. J. Bioinform. Comput. Biol. 2005, 3, 185–205. [Google Scholar] [CrossRef]
- MathWorks, fscmrmr Documentation. Available online: https://www.mathworks.com/help/stats/fscmrmr.html (accessed on 8 August 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | High TNF Level (ng/mL) | Importance Score | |
|---|---|---|---|
| NFκB | ATF-2 | ||
| 30 min | 0.082 | 0.18 | 0 |
| 3.2 | 0.49 | 0.15 | |
| 50 | 0.57 | 0.2 | |
| 4 h | 0.082 | 0.05 | 0.01 |
| 3.2 | 0.3 | 0.1 | |
| 50 | 0.24 | 0.07 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emadi, A.; Lipniacki, T.; Levchenko, A.; Abdi, A. Single-Cell Measurements and Modeling and Computation of Decision-Making Errors in a Molecular Signaling System with Two Output Molecules. Biology 2023, 12, 1461. https://doi.org/10.3390/biology12121461
Emadi A, Lipniacki T, Levchenko A, Abdi A. Single-Cell Measurements and Modeling and Computation of Decision-Making Errors in a Molecular Signaling System with Two Output Molecules. Biology. 2023; 12(12):1461. https://doi.org/10.3390/biology12121461
Chicago/Turabian StyleEmadi, Ali, Tomasz Lipniacki, Andre Levchenko, and Ali Abdi. 2023. "Single-Cell Measurements and Modeling and Computation of Decision-Making Errors in a Molecular Signaling System with Two Output Molecules" Biology 12, no. 12: 1461. https://doi.org/10.3390/biology12121461