
Bioactive Components in Fruit Interact with Gut Microbes
Abstract
:Simple Summary
Abstract
1. Introduction
2. Effects of GM on Human Disease
2.1. GI Disorders
2.1.1. Inflammatory Bowel Disease (IBD)
2.1.2. Colorectal Cancer (CRC)
2.2. Metabolic Disorders
2.2.1. Obesity
2.2.2. Diabetes
2.3. Neurodegenerative Disease
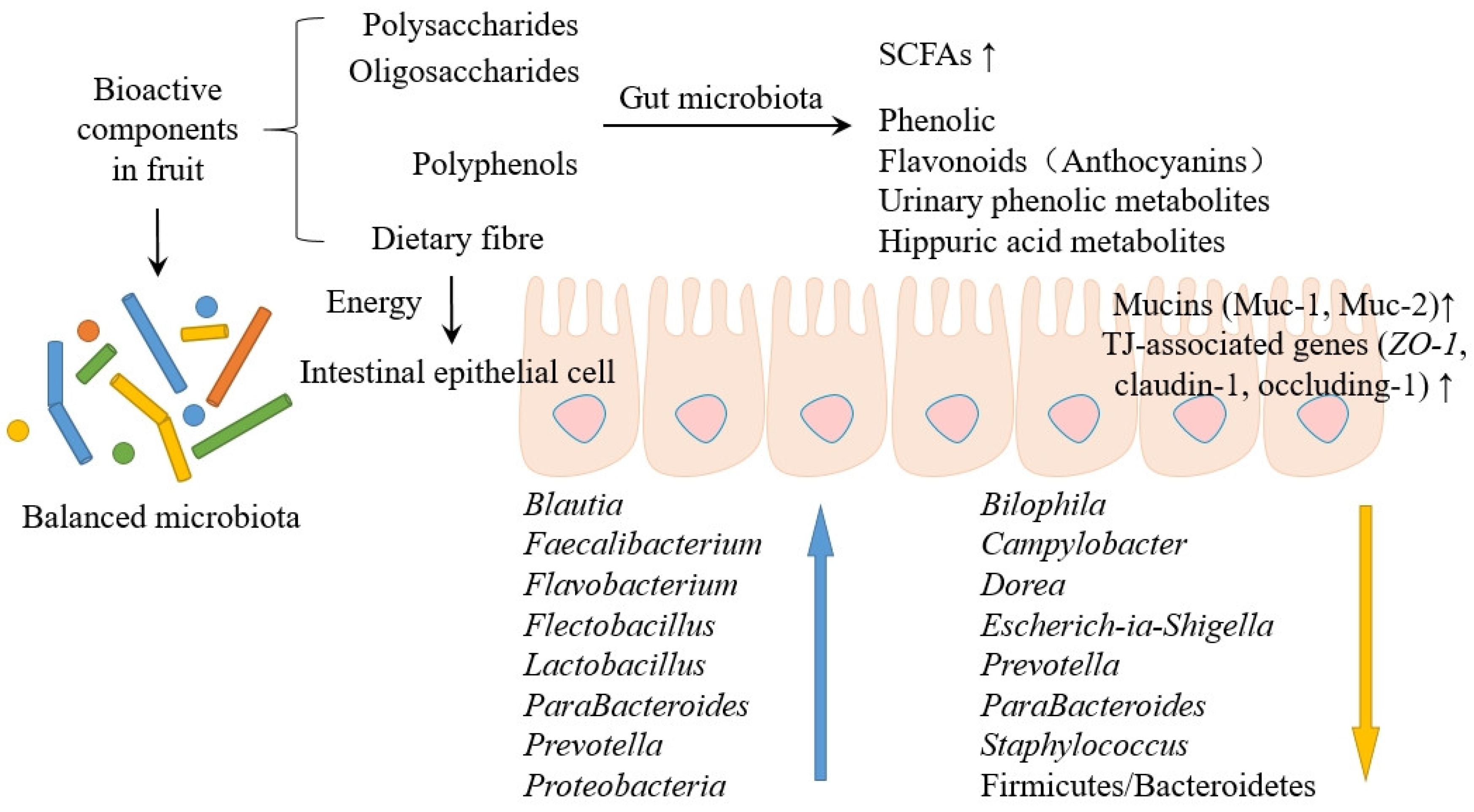
3. Fruit Bioactives’ Effect on GM and Possible Health Benefits
3.1. Fruit Bioactives’ Effect on GM
3.1.1. Polysaccharides
3.1.2. Oligosaccharides
3.1.3. DF
3.1.4. Polyphenols
3.1.5. Anthocyanins
3.1.6. Flavonoids
3.2. The Efficacy of Fruit-Derived Compounds In Vivo Experiments
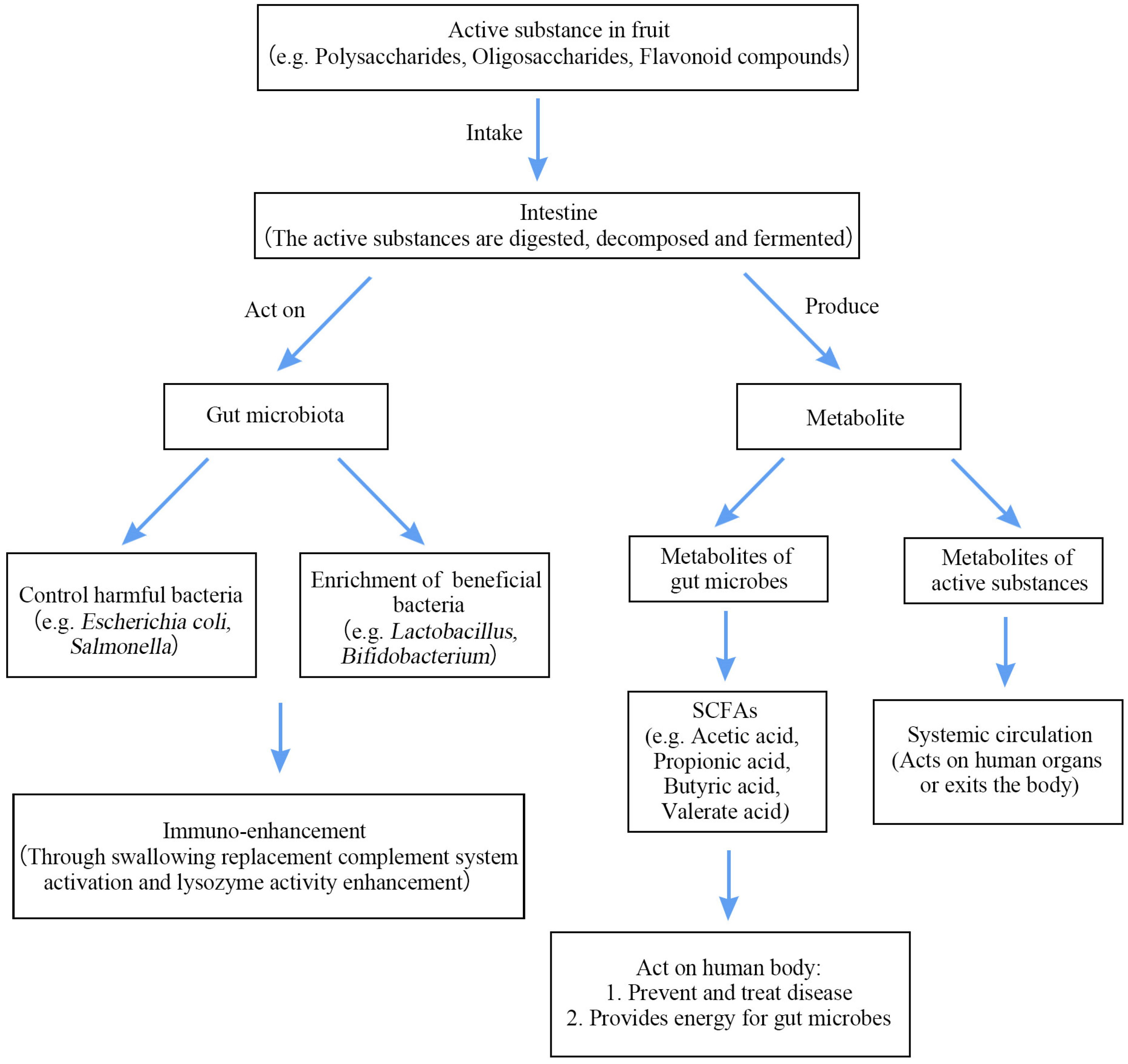
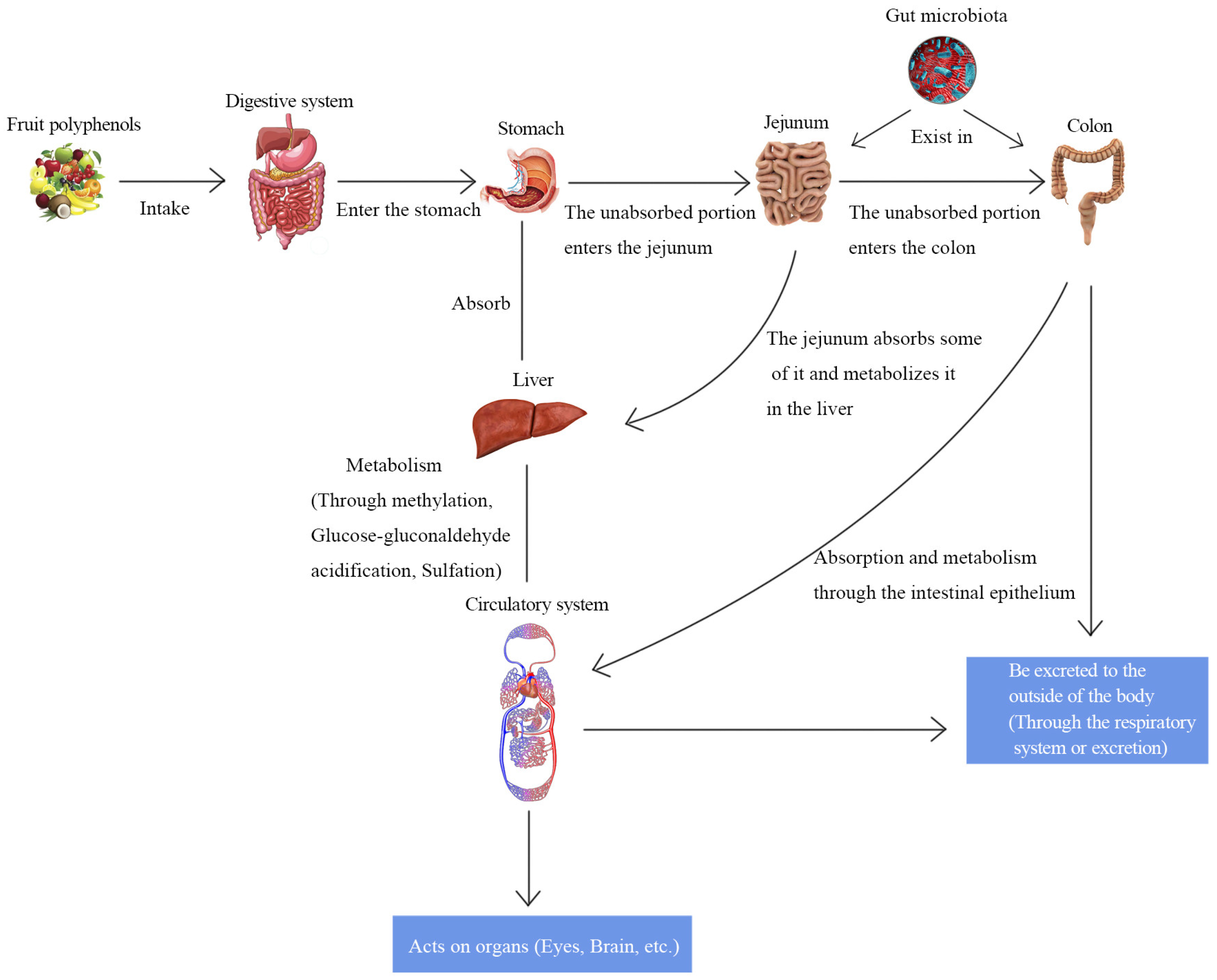
4. Interaction of Bioactive Ingredients with the GM
4.1. Absorption and Metabolism of Bioactive Ingredients in the Intestine
4.2. Mechanism of Interaction between Fruits and Gut Microorganisms
4.2.1. Inhibiting the Growth of Harmful Bacteria
4.2.2. Enrichment of Beneficial Bacteria
4.2.3. Production of Metabolites
4.3. Regulating the Immune System
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rakoff-Nahoum, S.; Coyne, M.J.; Comstock, L.E. An Ecological Network of Polysaccharide Utilization among Human Intestinal Symbionts. Curr. Biol. 2014, 24, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.L.; Wang, X.; Li, L.J. Human gut microbiome: The second genome of human body. Protein Cell 2010, 1, 718–725. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M.; de Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Larsbrink, J.; Zhu, Y.T.; Kharade, S.S.; Kwiatkowski, K.J.; Eijsink, V.G.H.; Koropatkin, N.M.; McBride, M.J.; Pope, P.B. A polysaccharide utilization locus from Flavobacterium johnsoniae enables conversion of recalcitrant chitin. Biotechnol. Biofuels 2016, 9, 260. [Google Scholar] [CrossRef] [PubMed]
- Oniszczuk, A.; Oniszczuk, T.; Gancarz, M.; Szymańska, J. Role of Gut Microbiota, Probiotics and Prebiotics in the Cardiovascular Diseases. Molecules 2021, 26, 1172. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Pérez, L.; Gosalbes, M.J.; Yuste, S.; Valls, R.M.; Pedret, A.; Llauradó, E.; Jimenez-Hernandez, N.; Artacho, A.; Pla-Pagà, L.; Companys, J.; et al. Gut metagenomic and short chain fatty acids signature in hypertension: A cross-sectional study. Sci. Rep. 2020, 10, 6436. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Shen, X.; Chu, Q.; Zheng, X. Pomegranate fruit pulp polyphenols reduce diet-induced obesity with modulation of gut microbiota in mice. J. Sci. Food Agric. 2022, 102, 1968–1977. [Google Scholar] [CrossRef]
- Alasalvar, C.; Chang, S.K.; Kris-Etherton, P.M.; Sullivan, V.K.; Petersen, K.S.; Guasch-Ferré, M.; Jenkins, D.J. Dried Fruits: Bioactives, Effects on Gut Microbiota, and Possible Health Benefits—An Update. Nutrients 2023, 15, 1611. [Google Scholar] [CrossRef]
- Borgonovi, T.F.; Virgolin, L.B.; Janzantti, N.S.; Casarotti, S.N.; Penna, A.L.B. Fruit bioactive compounds: Effect on lactic acid bacteria and on intestinal microbiota. Food Res. Int. 2022, 161, 111809. [Google Scholar] [CrossRef]
- Peerakietkhajorn, S.; Jeanmard, N.; Chuenpanitkit, P.; K-Da, S.; Khuituan, P. Effects of Plant Oligosaccharides Derived from Dragon Fruit on Gut Microbiota in Proximal and Distal Colon of Mice. Sains Malays. 2020, 49, 603–611. [Google Scholar] [CrossRef]
- Zahid, H.F.; Ranadheera, C.S.; Fang, Z.; Ajlouni, S. Utilization of Mango, Apple and Banana Fruit Peels as Prebiotics and Functional Ingredients. Agriculture 2021, 11, 584. [Google Scholar] [CrossRef]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst. Rev. 2015, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Cani, P.D. Gut microbiota and GLP-1. Rev. Endocr. Metab. Disord. 2014, 15, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Arthur, S.; Haynes, J.; Butts, M.R.; Nepal, N.; Sundaram, U. The Role of Gut Microbiota and Metabolites in Obesity-Associated Chronic Gastrointestinal Disorders. Nutrients 2022, 14, 624. [Google Scholar] [CrossRef]
- Lazar, V.; Ditu, L.-M.; Pircalabioru, G.G.; Picu, A.; Petcu, L.; Cucu, N.; Chifiriuc, M.C. Gut Microbiota, Host Organism, and Diet Trialogue in Diabetes and Obesity. Front. Nutr. 2019, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Westfall, S.; Lomis, N.; Kahouli, I.; Dia, S.Y.; Singh, S.P.; Prakash, S. Microbiome, probiotics and neurodegenerative diseases: Deciphering the gut brain axis. Cell. Mol. Life Sci. 2017, 74, 3769–3787. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Wu, X.; Li, M.; Li, X.; Huang, R.; Sun, Y.; Xu, Z. Noni (Morinda citrifolia L.) Fruit Polysaccharides Regulated IBD Mice Via Targeting Gut Microbiota: Association of JNK/ERK/NF-κB Signaling Pathways. J. Agric. Food Chem. 2021, 69, 10151–10162. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yue, B.; Wang, H.; Zhang, B.; Luo, X.; Yu, Z.; Zhang, J.; Ren, Y.; Mani, S.; Wang, Z. Acacetin ameliorates experimental colitis in mice via inhibiting macrophage inflammatory response and regulating the composition of gut microbiota. Front. Physiol. 2021, 11, 1858. [Google Scholar] [CrossRef] [PubMed]
- Cong, J.; Zhu, H.; Liu, D.; Li, T.; Zhang, C.; Zhu, J.; Lv, H.; Liu, K.; Hao, C.; Tian, Z.; et al. A Pilot Study: Changes of Gut Microbiota in Post-surgery Colorectal Cancer Patients. Front. Microbiol. 2018, 9, 2777. [Google Scholar] [CrossRef]
- Bai, Y.-f.; Wang, S.-w.; Wang, X.-x.; Weng, Y.-y.; Fan, X.-y.; Sheng, H.; Zhu, X.-t.; Lou, L.-j.; Zhang, F. The flavonoid-rich Quzhou Fructus Aurantii extract modulates gut microbiota and prevents obesity in high-fat diet-fed mice. Nutr. Diabetes 2019, 9, 30. [Google Scholar] [CrossRef]
- Liu, P.; Zhou, W.; Xu, W.; Peng, Y.; Yan, Y.; Lu, L.; Mi, J.; Zeng, X.; Cao, Y. The Main Anthocyanin Monomer from Lycium ruthenicum Murray Fruit Mediates Obesity via Modulating the Gut Microbiota and Improving the Intestinal Barrier. Foods 2022, 11, 98. [Google Scholar] [CrossRef]
- Gaike, A.H.; Paul, D.; Bhute, S.; Dhotre, D.P.; Pande, P.; Upadhyaya, S.; Reddy, Y.; Sampath, R.; Ghosh, D.; Chandraprabha, D.; et al. The Gut Microbial Diversity of Newly Diagnosed Diabetics but Not of Prediabetics Is Significantly Different from That of Healthy Nondiabetics. mSystems 2020, 5, 519. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, A.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Functional Deficits in Gut Microbiome of Young and Middle-Aged Adults with Prediabetes Apparent in Metabolizing Bioactive (Poly)phenols. Nutrients 2020, 12, 3595. [Google Scholar] [CrossRef]
- Wang, S.-s.; Li, X.-h.; Liu, P.; Li, J.; Liu, L. The relationship between Alzheimer’s disease and intestinal microflora structure and inflammatory factors. Front. Aging Neurosci. 2022, 14, 972982. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Li, Y.; Xu, X.; Li, R.; Zhang, M.; Cui, Y.; Zhang, L.; Wei, Z.; Wang, S.; Tuo, H. Probiotics for constipation and gut microbiota in Parkinson’s disease. Park. Relat. Disord. 2022, 103, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Yi, X.; Zhang, X.; Wang, H.; Liu, H.; Mou, W.-W. Imbalance in the Gut Microbiota of Children With Autism Spectrum Disorders. Front. Cell. Infect. Microbiol. 2021, 11, 572752. [Google Scholar] [CrossRef]
- Guan, Q. A Comprehensive Review and Update on the Pathogenesis of Inflammatory Bowel Disease. J. Immunol. Res. 2019, 2019, 7247238. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Mah, C.; Koentgen, S.; Zhang, L.; Grimm, M.C.; El-Omar, E.; Hold, G.L. Inflammatory bowel disease and the gut microbiota. Proc. Nutr. Soc. 2021, 80, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Ishimoto, T.; Fu, L.; Zhang, J.; Zhang, Z.; Liu, Y. The Gut Microbiota in Inflammatory Bowel Disease. Front. Cell. Infect. Microbiol. 2022, 12, 733992. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Juillerat, P.; Øyås, O.; Ramon, C.; Bravo, F.D.; Franc, Y.; Fournier, N.; Michetti, P.; Mueller, C.; Macpherson, A.J.; et al. Microbial network disturbances in relapsing refractory Crohn’s disease. Nat. Med. 2019, 25, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Pu, D.; Zhang, Z.; Feng, B. Alterations and Potential Applications of Gut Microbiota in Biological Therapy for Inflammatory Bowel Diseases. Front. Pharmacol. 2022, 13, 906419. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell. Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Hou, X.; Bian, Z.; Jia, W.; Zhao, L. Gut microbiota and colorectal cancer metastasis. Cancer Lett. 2023, 555, 216039. [Google Scholar] [CrossRef]
- Si, H.; Yang, Q.; Hu, H.; Ding, C.; Wang, H.; Lin, X. Colorectal cancer occurrence and treatment based on changes in intestinal flora. Semin. Cancer Biol. 2021, 70, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Maisonneuve, C.; Irrazabal, T.; Martin, A.; Girardin, S.E.; Philpott, D.J. The Impact of the Gut Microbiome on Colorectal Cancer. Annu. Rev. Cancer Biol. 2018, 2, 229–249. [Google Scholar] [CrossRef]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Obesity and Obesity-Associated Metabolic Disorders: Current Evidence and Perspectives. Curr. Obes. Rep. 2019, 8, 317–332. [Google Scholar] [CrossRef]
- Pan, W.; Zhang, Y.; Zeng, C.; Xu, F.; Yan, J.; Weng, J. miR-192 is upregulated in T1DM, regulates pancreatic β-cell development and inhibits insulin secretion through suppressing GLP-1 expression. Exp. Ther. Med. 2018, 16, 2717–2724. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, X.; Sun, L.; Liu, Y.; Lv, Y.; Gang, X.; Wang, G. Gut Microbiota Profile in Patients with Type 1 Diabetes Based on 16S rRNA Gene Sequencing: A Systematic Review. Dis. Markers 2020, 8, 3936247. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chang, Y.; Zhang, K.; Chen, H.; Tao, S.; Zhang, Z. Implication of the gut microbiome composition of type 2 diabetic patients from northern China. Sci. Rep. 2020, 10, 5450. [Google Scholar] [CrossRef] [PubMed]
- Vuotto, C.; Battistini, L.; Caltagirone, C.; Borsellino, G. Gut Microbiota and Disorders of the Central Nervous System. Neuroscientist 2020, 26, 487–502. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, X.; Zheng, Y.; Liu, W.; Ding, C. Aronia melanocarpa polysaccharide ameliorates inflammation and aging in mice by modulating the AMPK/SIRT1/NF-κB signaling pathway and gut microbiota. Nat. Publ. Group 2021, 11, 20558. [Google Scholar] [CrossRef]
- Lee, B.-H.; Hsu, K.-T.; Chen, Y.-Z.; Tain, Y.-L.; Hou, C.-Y.; Lin, Y.-C.; Hsu, W.-H. Polysaccharide extracts derived from defloration waste of fruit Pitaya regulates gut microbiota in a mice model. Fermentation 2022, 8, 108. [Google Scholar] [CrossRef]
- Song, S.K.; Beck, B.R.; Kim, D.; Park, J.; Kim, J.; Kim, H.D.; Ringø, E. Prebiotics as immunostimulants in aquaculture: A review. Fish Shellfish Immunol. 2014, 40, 40–48. [Google Scholar] [CrossRef]
- Li, E.; Long, X.; Liao, S.; Pang, D.; Li, Q.; Zou, Y. Effect of mulberry galacto-oligosaccharide isolated from mulberry on glucose metabolism and gut microbiota in a type 2 diabetic mice. J. Funct. Foods 2021, 87, 104836. [Google Scholar] [CrossRef]
- Dou, Z.; Chen, C.; Qiang, H.; Fu, X. In vitro digestion of the whole blackberry fruit: Bioaccessibility, bioactive variation of active ingredients and impacts on human gut microbiota. Food Chem. 2021, 370, 131001. [Google Scholar] [CrossRef]
- Song, H.Z.; Shen, X.C.; Deng, R.; Zhang, Y.; Zheng, X.D. Dietary anthocyanin-rich extract of açai protects from diet-induced obesity, liver steatosis, and insulin resistance with modulation of gut microbiota in mice. Nutrition 2021, 86, 111176. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.Y.; Hao, Y.F.; Hao, Z.X.; Liu, Q.; Zhang, L.; Jiang, C.P.; Jiang, J.G. Flavonoids from Rosa davurica Pall. fruits prevent high-fat diet-induced obesity and liver injury via modulation of the gut microbiota in mice. Food Funct. 2021, 12, 10097–10106. [Google Scholar] [CrossRef] [PubMed]
- Han, X.P.; Song, Y.; Huang, R.M.; Zhu, M.Q.; Li, M.Y.; Requena, T.; Wang, H. Anti-Inflammatory and Gut Microbiota Modulation Potentials of Flavonoids Extracted from Passiflora foetida Fruits. Foods 2023, 12, 2889. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, S.; Zhang, J.; Che, G.; Zhou, M.; Liu, M.; Zhang, C.; Xu, N.; Lin, L.; Liu, Y.; et al. The antihyperlipidemic activities of enzymatic and acidic intracellular polysaccharides by Termitomyces albuminosus. Carbohydr. Polym. 2016, 151, 1227–1234. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.-M.; Qin, G.-Y. Structure characterization and antioxidant activity of polysaccharides from Chinese quince seed meal. Food Chem. 2017, 234, 314–322. [Google Scholar] [CrossRef]
- Li, X.; Guo, R.; Wu, X.; Liu, X.; Ai, L.; Sheng, Y.; Song, Z.; Wu, Y. Dynamic digestion of tamarind seed polysaccharide: Indigestibility in gastrointestinal simulations and gut microbiota changes in vitro. Carbohydr. Polym. 2020, 239, 116194. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Ke, Y.; Li, C.; Zhang, Z.; Wu, Y.; Hu, B.; Liu, A.; Luo, Q.; Wu, W. In vitro saliva-gastrointestinal digestion and fecal fermentation of Oudemansiella radicata polysaccharides reveal its digestion profile and effect on the modulation of the gut microbiota. Carbohydr. Polym. 2021, 251, 117041. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xu, Y.; Su, J.; Zhu, B.; Wang, S.; Liu, K.; Wang, H.; Shi, S.; Zhang, Q.; Qin, L.; et al. Roles of gut microbiota and metabolites in a homogalacturonan-type pectic polysaccharide from Ficus pumila Linn. fruits mediated amelioration of obesity. Carbohydr. Polym. 2020, 248, 116780. [Google Scholar] [CrossRef]
- Lieu, L.N. Functional compounds in dragon fruit peels and their potential health benefits: A review. Int. J. Food Sci. Technol. 2021, 57, 2571–2580. [Google Scholar]
- Liu, H.; Zhang, Z.; Li, J.P.; Liu, W.; Warda, M.; Cui, B.; El-Aty, A.M.A. Oligosaccharides derived from Lycium barbarum ameliorate glycolipid metabolism and modulate the gut microbiota community and the faecal metabolites in a type 2 diabetes mouse model: Metabolomic bioinformatic analysis. Food Funct. 2022, 13, 5416–5429. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.Y.; Zhou, L.N.; Fu, Z.L.; Zhang, B.; Li, Q.C.; Pan, L.; Zhou, C.; Zhao, Q.; Shang, Q.; Yu, G.L. Enterotype-Specific Effect of Human Gut Microbiota on the Fermentation of Marine Algae Oligosaccharides: A Preliminary Proof-of-Concept In Vitro Study. Polymers 2022, 14, 770. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.T.; Cheong, K. Recent advances in marine algae oligosaccharides: Structure, analysis, and potential prebiotic activities. Crit. Rev. Food Sci. Nutr. 2021, 62, 7703–7717. [Google Scholar] [CrossRef] [PubMed]
- Sangkuanun, T.; Wichienchot, S.; Kato, Y.; Watanabe, H.; Peerakietkhajorn, S. Oligosaccharides derived from dragon fruit modulate gut microbiota, reduce oxidative stress and stimulate toll-pathway related gene expression in freshwater crustacean Daphnia magna—ScienceDirect. Fish Shellfish Immunol. 2020, 103, 126–134. [Google Scholar] [CrossRef]
- Hijová, E.; Bertková, I.; Štofilová, J. Dietary fibre as prebiotics in nutrition. Cent. Eur. J. Public Health 2019, 27, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Barber, T.M.; Kabisch, S.; Pfeiffer, A.F.H.; Weickert, M.O. The Health Benefits of Dietary Fibre. Nutrients 2020, 12, 3209. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Nguyen, L.H.; Song, M.; Jovani, M.; Chan, A.T. Intake of Dietary Fiber, Fruits, and Vegetables and Risk of Diverticulitis. Am. J. Gastroenterol. 2019, 114, 1. [Google Scholar] [CrossRef] [PubMed]
- Prado, S.B.R.D.; Minguzzi, B.T.; Hoffmann, C.; Fabi, J.P. Modulation of Human Gut Microbiota by Dietary Fibers from Unripe and Ripe Papayas: Distinct Polysaccharide Degradation using a Colonic In Vitro Fermentation Model. Food Chem. 2021, 348, 129071. [Google Scholar] [CrossRef]
- Hernández-Maldonado, L.M.; Blancas-Benítez, F.J.; Zamora-Gasga, V.M.; Cárdenas-Castro, A.P.; Sáyago-Ayerdi, S.G. In Vitro Gastrointestinal Digestion and Colonic Fermentation of High Dietary Fiber and Antioxidant-Rich Mango (Mangifera indica L.) “Ataulfo”-Based Fruit Bars. Nutrients 2019, 11, 1564. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef]
- Parkar, S.G.; Stevenson, D.E.; Skinner, M.A. The potential influence of fruit polyphenols on colonic microflora and human gut health. Int. J. Food Microbiol. 2008, 124, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, K.T.R.; Moreira, L.d.S.G.; Borges, N.A.; Brum, I.; Paiva, B.R.d.; Alvarenga, L.; Nakao, L.S.; Leal, V.d.O.; Carraro-Eduardo, J.C.; Rodrigues, S.D.; et al. Effect of cranberry supplementation on toxins produced by the gut microbiota in chronic kidney disease patients: A pilot randomized placebo-controlled trial. Clin. Nutr. ESPEN 2022, 47, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morató, J.; Matthan, N.R.; Liu, J.; Torre, R.d.l.; Chen, C.-Y.O. Cranberries attenuate animal-based diet-induced changes in microbiota composition and functionality: A randomized crossover controlled feeding trial. J. Nutr. Biochem. 2018, 62, 76–86. [Google Scholar] [CrossRef]
- Kumari, M.; Kozyrskyj, A.L. Gut microbial metabolism defines host metabolism: An emerging perspective in obesity and allergic inflammation. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2017, 18, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, A.; Lamiae, B.; Samira, B.; Ramchoun, M.; Abdelouahed, K.; Tamas, F.; Hicham, B. Pomegranate (Punica granatum L.) Attenuates Neuroinflammation Involved in Neurodegenerative Diseases. Foods 2022, 11, 2570. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mejía, E.; Roriz, C.L.; Heleno, S.A.; Calhelha, R.; Dias, M.I.; Pinela, J.; Rosales-Conrado, N.; León-González, M.E.; Ferreira, I.C.F.R.; Barros, L. Valorisation of black mulberry and grape seeds: Chemical characterization and bioactive potential. Food Chem. 2021, 337, 127998. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Gan, T.; Huang, Y.; Bao, L.; Liu, S.; Cui, X.; Wang, H.; Jiao, F.; Zhang, M.; Su, C.; et al. Anti-Inflammatory Activity of Mulberry Leaf Flavonoids In Vitro and In Vivo. Int. J. Mol. Sci. 2022, 23, 7694. [Google Scholar] [CrossRef] [PubMed]
- Tomas, M.; Toydemir, G.; Boyacioglu, D.; Hall, R.D.; Beekwilder, J.; Capanoglu, E. Processing black mulberry into jam: Effects on antioxidant potential and in vitro bioaccessibility. J. Sci. Food Agric. 2017, 97, 3106–3113. [Google Scholar] [CrossRef]
- Wu, T.; Yin, J.; Zhang, G.; Long, H.R.; Zheng, X. Mulberry and cherry anthocyanin consumption prevents oxidative stress and inflammation in diet-induced obese mice. Mol. Nutr. Food Res. 2016, 60, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Y.; Xiang, J.; Wang, C.; Johnson, J.B.; Beta, T. Diverse polyphenol components contribute to antioxidant activity and hypoglycemic potential of mulberry varieties. LWT-Food Sci. Technol. 2023, 173, 114308. [Google Scholar] [CrossRef]
- Liu, J.; Hao, W.; He, Z.; Kwek, E.; Zhu, H.; Ma, N.; Ma, K.Y.; Chen, Z.Y. Blueberry and cranberry anthocyanin extracts reduce bodyweight and modulate gut microbiota in C57BL/6J mice fed with a high-fat diet. Eur. J. Nutr. 2021, 60, 2735–2746. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Deng, R.; Chu, Q.; Zheng, X. Pomegranate peel anthocyanins prevent diet-induced obesity and insulin resistance in association with modulation of the gut microbiota in mice. Eur. J. Nutr. 2022, 61, 1837–1847. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Ni, J.; Zhang, M.; Xu, Y.; Li, Y.; Karim, N.; Chen, W. Mulberry Anthocyanins Ameliorate DSS-Induced Ulcerative Colitis by Improving Intestinal Barrier Function and Modulating Gut Microbiota. Antioxidants 2022, 11, 1647. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Li, X.; He, S.; Gao, C.; Wang, C.; Shao, Y. Effects of Natural Flavonoid-Isoorientin on Growth Performance and Gut Microbiota of Mice. J. Agric. Food Chem. 2018, 66, 9777–9784. [Google Scholar] [CrossRef] [PubMed]
- Pierre, J.F.; Heneghan, A.F.; Feliciano, R.P.; Shanmuganayagam, D.; Krueger, C.G.; Reed, J.D.; Kudsk, K.A. Cranberry Proanthocyanidins Improve Intestinal sIgA During Elemental Enteral Nutrition. J. Parenter. Enter. Nutr. 2014, 38, 107–114. [Google Scholar] [CrossRef]
- Sudheeran, P.K.; Ovadia, R.; Galsarker, O.; Maoz, I.; Sela, N.; Maurer, D.; Feygenberg, O.; Shamir, M.O.; Alkan, N. Glycosylated flavonoids: Fruit’s concealed antifungal arsenal. New Phytol. 2020, 225, 1788–1798. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.-X.; Gao, A.X.; Zhu, Y.; Dong, T.T.-X.; Tsim, K.W.-K. Flavonoids from Seabuckthorn (Hippophae rhamnoides L.) restore CUMS-induced depressive disorder and regulate the gut microbiota in mice. Food Funct. 2023, 14, 7426–7438. [Google Scholar] [CrossRef]
- Meng, W.; Zhao, Z.; Chen, L.; Lin, S.; Zhang, Y.; He, J.; Ouyang, K.; Wang, W. Total Flavonoids from Chimonanthus nitens Oliv. Leaves Ameliorate HFD-Induced NAFLD by Regulating the Gut–Liver Axis in Mice. Foods 2022, 11, 2169. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, B.; Zhang, X.; Shen, X.; Ma, Y.; Jing, L. Lycium barbarum polysaccharide antagonizes cardiomyocyte apoptosis by inhibiting the upregulation of GRK2 induced by I/R injury, and salvage mitochondrial fission/fusion imbalance and AKT/eNOS signaling. Cell. Signal. 2022, 92, 110252. [Google Scholar] [CrossRef] [PubMed]
- Pansai, N.; Detarun, P.; Chinnaworn, A.; Sangsupawanich, P.; Wichienchot, S. Effects of dragon fruit oligosaccharides on immunity, gut microbiome, and their metabolites in healthy adults—A randomized double-blind placebo controlled study. Food Res. Int. 2023, 167, 112657. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Aparicio, I.; De La Peña Armada, R.; Pérez-Cózar, M.L.; Rupérez, P.; Redondo-Cuenca, A.; Villanueva-Suárez, M.J. Apple by-product dietary fibre exhibits potential prebiotic and hypolipidemic effectsin high-fat fed Wistar rats. Bioact. Carbohydr. Diet. Fibre 2020, 23, 100219. [Google Scholar] [CrossRef]
- Cladis, D.P.; Simpson, A.M.R.; Cooper, K.J.; Nakatsu, C.H.; Ferruzzi, M.G.; Weaver, C.M. Blueberry polyphenols alter gut microbiota phenolic metabolism in rats. Food Funct. 2021, 12, 2442–2456. [Google Scholar] [CrossRef]
- Song, H.; Zhang, Y.; Huang, Q.; Deng, R.; Zheng, X. Averrhoa carambola L. fruit polyphenols ameliorate hyperlipidemia, hepatic steatosis, and hyperglycemia by modulating lipid and glucose metabolism in mice with obesity. J. Sci. Food Agric. 2023, 103, 6531–6539. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, S.; Tang, L.; Zhang, M.; Wang, P.; Sun, X.; Shang, L.; Wang, Q.; Zhao, Y.; Meng, D.; et al. Analysis of the effects of Rosa roxburghii Tratt fruit polyphenols on immune function in mice through gut microbiota and metabolomics: An in vivo preclinical trial study. J. Funct. Foods 2023, 102, 105464. [Google Scholar] [CrossRef]
- Chen, S.; Hu, N.; Wang, H.; Wu, Y.; Li, G. Bioactivity-guided isolation of the major anthocyanin from Lycium ruthenicum Murr. fruit and its antioxidant activity and neuroprotective effects in vitro and in vivo. Food Funct. 2022, 13, 3247–3257. [Google Scholar] [CrossRef]
- Villa-Jaimes, G.S.; Moshage, H.; Avelar-González, F.J.; González-Ponce, H.A.; Buist-Homan, M.; Guevara-Lara, F.; Sánchez-Alemán, E.; Martínez-Hernández, S.L.; Ventura-Juárez, J.; Muñoz-Ortega, M.H.; et al. Molecular and Antioxidant Characterization of Opuntia robusta Fruit Extract and Its Protective Effect against Diclofenac-Induced Acute Liver Injury in an In Vivo Rat Model. Antioxidants 2023, 12, 113. [Google Scholar] [CrossRef]
- Zhan, M.; Liang, X.; Chen, J.; Yang, X.; Han, Y.; Zhao, C.; Xiao, J.; Cao, Y.; Xiao, H.; Song, M. Dietary 5-demethylnobiletin prevents antibiotic-associated dysbiosis of gut microbiota and damage to the colonic barrier. Food Funct. 2023, 14, 4414–4429. [Google Scholar] [CrossRef]
- Dobranowski, P.A.; Stintzi, A. Resistant starch, microbiome, and precision modulation. Gut Microbes 2021, 13, 1926842. [Google Scholar] [CrossRef]
- Li, S.; Li, T.; Wang, B.; Wen, C.; Li, M.; Ding, K. A structure defined pectin SA02B from Semiaquilegia adoxoides is metabolized by human gut microbes. Int. J. Biol. Macromol. 2023, 234, 123673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.L.; Mao, Y.Q.; Zhang, Z.Y.; Li, Z.M.; Wang, L.S. Pectin supplement significantly enhanced the anti-PD-1 efficacy in tumor-bearing mice humanized with gut microbiota from patients with colorectal cancer. Theranostics 2021, 11, 4155–4170. [Google Scholar] [CrossRef]
- Oku, T.; Nakamura, S. Fructooligosaccharide: Metabolism through Gut Microbiota and Prebiotic Effect. Food Nutr. J. 2017, 2, 128. [Google Scholar] [CrossRef]
- Tanno, H.; Fujii, T.; Hirano, K.; Maeno, S.; Endo, A. Characterization of fructooligosaccharide metabolism and fructooligosaccharide-degrading enzymes in human commensal butyrate producers. Gut Microbes 2021, 13, 869503. [Google Scholar] [CrossRef]
- Zhang, X.; Sandhu, A.; Edirisinghe, I.; Burton-Freeman, B. An exploratory study of red raspberry (Rubus idaeus L.) (poly)phenols/metabolites in human biological samples. Food Funct. 2018, 9, 806–818. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Mol. Nutr. Food Res. 2017, 61, 1500901. [Google Scholar] [CrossRef] [PubMed]
- Milala, J.; Kosmala, M.; Karlińska, E.b.; Juskiewicz, J.; Zdunczyk, Z.; Fotschki, B. Ellagitannins from Strawberry with Different Degree of Polymerization Showed Different Metabolism Through Gastrointestinal Tract of Rats. J. Agric. Food Chem. 2017, 65, 10738–10748. [Google Scholar] [CrossRef] [PubMed]
- Komarnytsky, S.; Wagner, C.; Gutierrez, J.; Shaw, O.M. Berries in Microbiome-Mediated Gastrointestinal, Metabolic, and Immune Health. Curr. Nutr. Rep. 2023, 12, 151–166. [Google Scholar] [CrossRef]
- Esteban-Torres, M.; Santamaría, L.; Cabrera-Rubio, R.; Plaza-Vinuesa, L.; Crispie, F.; Rivas, B.d.L.; Cotter, P.; Muñoz, R. A Diverse Range of Human Gut Bacteria Have the Potential To Metabolize the Dietary Component Gallic Acid. Am. Soc. Microbiol. 2018, 84, 1558. [Google Scholar] [CrossRef]
- Tomas-Barberan, F.; García-Villalba, R.; Quartieri, A.; Raimondi, S.; Amaretti, A.; Leonardi, A.; Rossi, M. In vitro transformation of chlorogenic acid by human gut microbiota. Mol. Nutr. Food Res. 2014, 58, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Peng, X.; Li, S.; Zhang, N.; Wang, Y.; Wei, H. Isolation and Identification of Quercetin Degrading Bacteria from Human Fecal Microbes. PLoS ONE 2014, 9, 90531. [Google Scholar] [CrossRef]
- Erlund, I. Review of the flavonoids quercetin, hesperetin, and naringenin. Dietary sources, bioactivities, bioavailability, and epidemiology. Nutr. Res. 2004, 24, 851–874. [Google Scholar] [CrossRef]
- Yang, J.; Qian, D.; Jiang, S.; Shang, E.-X.; Guo, J.; Duan, J.-A. Identification of rutin deglycosylated metabolites produced by human intestinal bacteria using UPLC–Q-TOF/MS. J. Chromatogr. B 2012, 898, 95–100. [Google Scholar] [CrossRef]
- Nectoux, A.M.; Abe, C.; Huang, S.-W.; Ohno, N.; Tabata, J.; Miyata, Y.; Tanaka, K.; Tanaka, T.; Yamamura, H.; Matsui, T. Absorption and Metabolic Behavior of Hesperidin (Rutinosylated Hesperetin) after Single Oral Administration to Sprague-Dawley Rats. J. Agric. Food Chem. 2019, 67, 9812–9819. [Google Scholar] [CrossRef]
- Wu, F.; Lei, H.; Chen, G. Multiomics Analyses Reveal That Long-Term Intake of Hesperetin-7-O-glucoside Modulates the Gut Microbiota and Bile Acid Metabolism in Mice. J. Agric. Food Chem. 2022, 70, 14831–14840. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Raskine, L.; Simoneau, G.; Vicaut, E.; Neut, C.; Flourié, B.; Brouns, F.; Bornet, F.R. The capacity of nondigestible carbohydrates to stimulate fecal bifidobacteria in healthy humans: A double-blind, randomized, placebo-controlled, parallel-group, dose-response relation study. Am. J. Clin. Nutr. 2004, 80, 1658–1664. [Google Scholar] [CrossRef]
- Foley, M.H.; Cockburn, D.W.; Koropatkin, N.M. The Sus operon: A model system for starch uptake by the human gut Bacteroidetes. Cell. Mol. Life Sci. 2016, 73, 2603–2617. [Google Scholar] [CrossRef]
- Heinken, A.; Khan, M.T.; Paglia, G.; Rodionov, D.A.; Harmsen, H.J.M.; Thiele, I. Functional Metabolic Map of Faecalibacterium prausnitzii, a Beneficial Human Gut Microbe. J. Bacteriol. 2014, 196, 3289–3302. [Google Scholar] [CrossRef]
- Cummings, J.H.; Stephen, A.M. Carbohydrate terminology and classification. Eur. J. Clin. Nutr. 2007, 61, S5–S18. [Google Scholar] [CrossRef] [PubMed]
- Bach Knudsen, K.E. Microbial Degradation of Whole-Grain Complex Carbohydrates and Impact on Short-Chain Fatty Acids and Health. Adv. Nutr. 2015, 6, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Bach Knudsen, K.E.; Nørskov, N.P.; Bolvig, A.K.; Hedemann, M.S.; Laerke, H.N. Dietary fibers and associated phytochemicals in cereals. Mol. Nutr. Food Res. 2017, 61, 1600518. [Google Scholar] [CrossRef] [PubMed]
- Pei, R.; Liu, X.; Bolling, B. Flavonoids and gut health. Curr. Opin. Biotechnol. 2020, 61, 153–159. [Google Scholar] [CrossRef]
- Feng, X.; Li, Y.; Brobbey Oppong, M.; Qiu, F. Insights into the intestinal bacterial metabolism of flavonoids and the bioactivities of their microbe-derived ring cleavage metabolites. Drug Metab. Rev. 2018, 50, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.F.; Maier, C.S. The Chemistry of Gut Microbial Metabolism of Polyphenols. Phytochem. Rev. 2016, 15, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Faria, A.; de Freitas, V.; Calhau, C.; Mateus, N. Multiple-approach studies to assess anthocyanin bioavailability. Phytochem. Rev. Proc. Phytochem. Soc. Eur. 2015, 14, 899–919. [Google Scholar] [CrossRef]
- Victoria-Campos, C.I.; Ornelas-Paz, J.d.J.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; Failla, M.L.; Pérez-Martínez, J.D.; Rios-Velasco, C.; Ibarra-Junquera, V. Gastrointestinal metabolism and bioaccessibility of selected anthocyanins isolated from commonly consumed fruits. Food Chem. 2022, 383, 132451. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Costa, E.M.; Mendes, M.; Morais, R.M.; Calhau, C.; Pintado, M.M. Antimicrobial, antiadhesive and antibiofilm activity of an ethanolic, anthocyanin-rich blueberry extract purified by solid phase extraction. J. Appl. Microbiol. 2016, 121, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Polewski, M.A.; Esquivel-Alvarado, D.; Wedde, N.S.; Kruger, C.G.; Reed, J.D. Isolation and Characterization of Blueberry Polyphenolic Components and Their Effects on Gut Barrier Dysfunction. J. Agric. Food Chem. 2020, 68, 2940–2947. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Zhu, J.; Tong, Y.; Kong, Y.; Tan, C.; Wang, M.; Wan, M.; Meng, X. Antibacterial characteristics and mechanisms of action of Aronia melanocarpa anthocyanins against Escherichia coli. LWT-Food Sci. Technol. 2021, 150, 112018. [Google Scholar] [CrossRef]
- Cai, M.; Zhang, G.; Li, C.; Chen, X.; Cui, H.; Lin, L. Pleurotus eryngii polysaccharide nanofiber containing pomegranate peel polyphenol/chitosan nanoparticles for control of E. coli O157:H7. Int. J. Biol. Macromol. 2021, 192, 939–949. [Google Scholar] [CrossRef]
- Pang, J.; Wang, S.; Wang, Z.; Wu, Y.; Zhang, X.; Pi, Y.; Han, D.; Zhang, S.; Wang, J. Xylo-oligosaccharide alleviates Salmonella induced inflammation by stimulating Bifidobacterium animalis and inhibiting Salmonella colonization. FASEB J. 2021, 35, 21977. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Lazarte, A.; Moreno, J.F.; Cueva, C.; Gil-Sánchez, I.; Villamiel, M. Behaviour of citrus pectin during its gastrointestinal digestion and fermentation in a dynamic simulator (simgi®). Carbohydr. Polym. 2019, 207, 382–390. [Google Scholar] [CrossRef]
- Schlotz, N.; Sørensen, J.G.; Martin-Creuzburg, D. The potential of dietary polyunsaturated fatty acids to modulate eicosanoid synthesis and reproduction in Daphnia magna: A gene expression approach. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 162, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, A.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Red raspberry and fructo-oligosaccharide supplementation, metabolic biomarkers, and the gut microbiota in adults with prediabetes: A randomized crossover clinical trial. J. Nutr. 2022, 152, 1438–1449. [Google Scholar] [CrossRef] [PubMed]
- Nogal, A.; Valdes, A.M.; Menni, C. The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut Microbes 2021, 13, 1897212. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The role of short-chain fatty acids in immunity, inflammation and metabolism. Crit. Rev. Food Sci. Nutr. 2022, 61, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, Z.; Ma, N.; Chen, Z.Y. Beneficial Effects of Dietary Polyphenols on High-Fat Diet-Induced Obesity Linking with Modulation of Gut Microbiota. J. Agric. Food Chem. 2020, 68, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.Q. Strategies to promote abundance of Akkermansia muciniphila, an emerging probiotics in the gut, evidence from dietary intervention studies. J. Funct. Foods 2017, 33, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Ze, X.; Duncan, S.H.; Louis, P.; Flint, H.J. Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J. Multidiscip. J. Microb. Ecol. 2012, 6, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Han, Z.; Kong, Q.; Wang, Y.; Mou, H.; Duan, X. Clostridium butyricum Prevents Dysbiosis and the Rise in Blood Pressure in Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2023, 24, 4955. [Google Scholar] [CrossRef]
- Pérez-Reytor, D.; Puebla, C.; Karahanian, E.; García, K. Use of Short-Chain Fatty Acids for the Recovery of the Intestinal Epithelial Barrier Affected by Bacterial Toxins. Front. Physiol. 2021, 12, 650313. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wu, S.; Guan, Y.; Ma, Y.; Huang, Y.; Liu, X.; Quan, D.; Zhang, J.; Lv, L.; Zhang, G. Changes in gut microbiota linked to a prevention of cardiac remodeling induced by hypertension in spontaneously hypertensive rats fed a pawpaw fruit diet. Heliyon 2023, 9, 15576. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Li, Q.; Yi, H.; Kuang, T.; Tang, Y.; Fan, G. Gut microbiota-derived metabolites as key actors in type 2 diabetes mellitus. Biomed. Pharmacother. 2022, 149, 112839. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.M.; Rangel, R.B. Gut microbiota-derived metabolites are novel targets for improving insulin resistance. World J. Diabetes Engl. Ed. (Electron. Version) 2022, 13, 65–69. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Tan, Y.; Yu, D.; Qiu, S.; Bai, Y.; He, J.; Cao, H.; Che, Q.; Guo, J.; Su, Z. The therapeutic effect of SCFA-mediated regulation of the intestinal environment on obesity. Front. Nutr. 2022, 9, 886902. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Heng, X.; Guo, L.; Lessing, D.J.; Chu, W. SCFAs improve disease resistance via modulate gut microbiota, enhance immune response and increase antioxidative capacity in the host. Fish Shellfish Immunol. 2022, 120, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.-L.; Hou, C.-Y.; Chang-Chien, G.-P.; Lin, S.-F.; Hsu, C.-N. Perinatal Propionate Supplementation Protects Adult Male Offspring from Maternal Chronic Kidney Disease-Induced Hypertension. Nutrients 2022, 14, 3435. [Google Scholar] [CrossRef] [PubMed]
- Ashique, S.; De Rubis, G.; Sirohi, E.; Mishra, N.; Rihan, M.; Garg, A.; Reyes, R.-J.; Manandhar, B.; Bhatt, S.; Jha, N.K.; et al. Short Chain Fatty Acids: Fundamental mediators of the gut-lung axis and their involvement in pulmonary diseases. Chem.-Biol. Interact. 2022, 368, 110231. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.T.; Macia, L.; Galvão, I.; Martins, F.S.; Canesso, M.C.C.; Amaral, F.A.; Garcia, C.C.; Maslowski, K.M.; De Leon, E.; Shim, D.; et al. A Role for Gut Microbiota and the Metabolite-Sensing Receptor GPR43 in a Murine Model of Gout. Arthritis Rheumatol. 2015, 67, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.Y.; Zheng, Y.Q.; Zhang, M.; Fei, W.D.; Sun, D.L.; Zhao, M.D.; Ye, Y.Q.; Zheng, C.H. Short-chain fatty acids regulate B cells differentiation via FFAR2 to alleviate rheumatoid arthritis. Br. J. Pharmacol. 2022, 179, 4315–4329. [Google Scholar] [CrossRef] [PubMed]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Yu, F.B.; Topf, M.; Gonzalez, C.; Robinson, J.L.; Elias, J.E. Gut Microbiota-Targeted Diets Modulate Human Immune Status. Cold Spring Harb. Lab. 2021, 184, 4137–4153. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Yan, Y.; Peng, Y.; Chen, D.; Mi, J.; Lu, L.; Luo, Q.; Li, X.; Zeng, X.; Cao, Y. In vitro digestion under simulated saliva, gastric and small intestinal conditions and fermentation by human gut microbiota of polysaccharides from the fruits of Lycium barbarum. Int. J. Biol. Macromol. 2019, 125, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, L.; Zhuang, H.; Yang, Z.; Jiang, G.; Liu, Z. Promoting intestinal IgA production in mice by oral administration with anthocyanins. Front. Immunol. 2022, 13, 826597. [Google Scholar] [CrossRef] [PubMed]
- Medina-Larqué, A.-S.; Rodríguez-Daza, M.-C.; Roquim, M.; Dudonné, S.; Pilon, G.; Levy, É.; Marette, A.; Roy, D.; Jacques, H.; Desjardins, Y. Cranberry polyphenols and agave agavins impact gut immune response and microbiota composition while improving gut barrier function, inflammation, and glucose metabolism in mice fed an obesogenic diet. Front. Immunol. 2022, 13, 871080. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
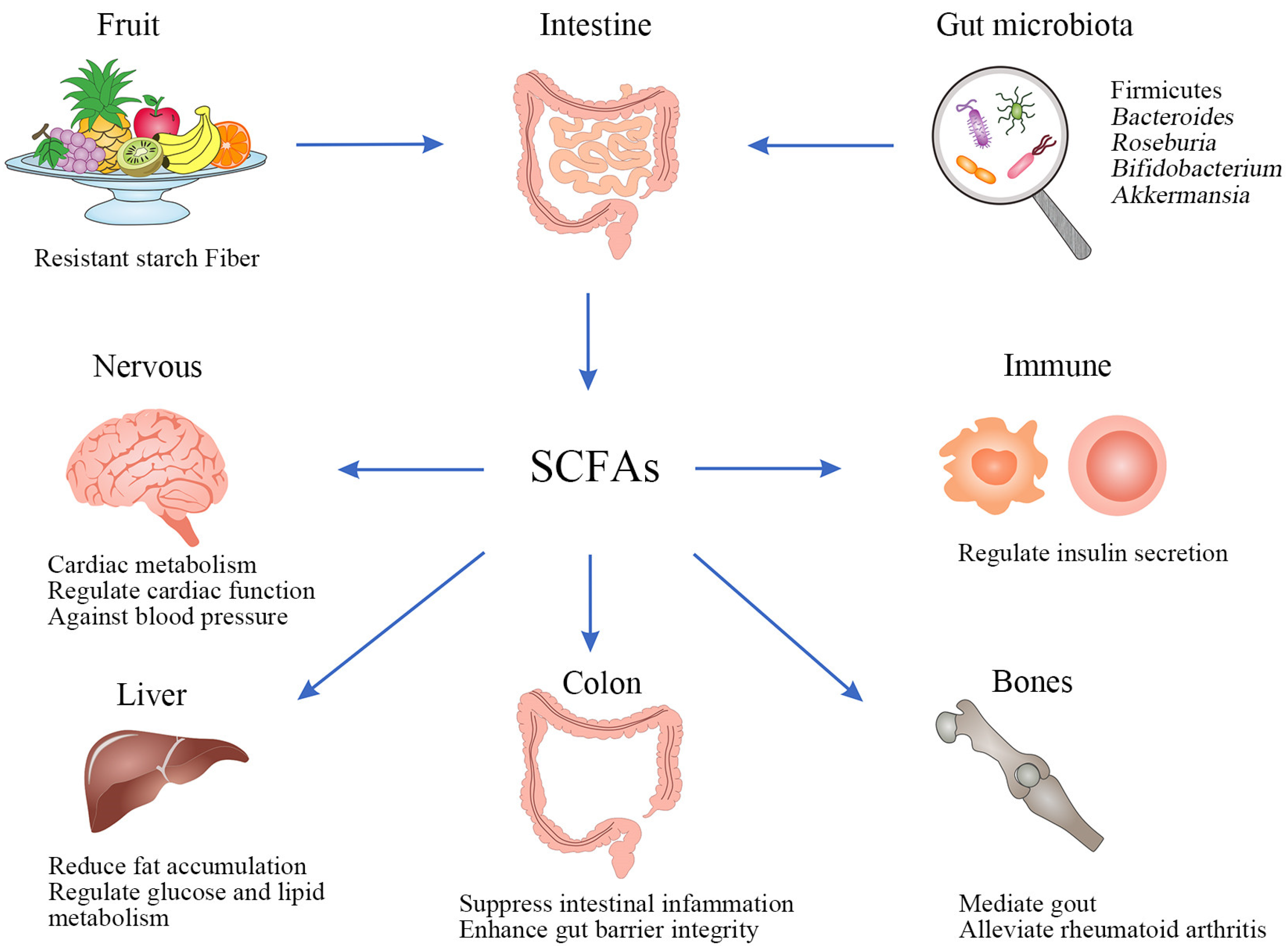{kind=link}
| Disease Types | Fruit Bioactives | Microbiota Influence | Others’ Influence | Conclusions | Reference |
|---|---|---|---|---|---|
| IBD | Noni (Morinda citrifolia L.) Fruit Polysaccharides | Relative abundance of Gram-positive bacteria ↓ Proteus and Spirospiral spp. ↓ | Acetic acid, propionic acid, and butyric acid ↑ JNK, ERK, and NF-κB phosphorylation ↓ LPS, TNF-α, and IL-17 ↓ | Gut microecological balance to inhibit inflammatory signaling pathways. | [19] |
| IBD | Acacetin, a Natural Dietary Flavonoid | Proteus and Shigella spp. ↓ | Alleviates body weight loss, diarrhea, colon shortening, inflammatory infiltration, and histological injury TNF-α, IL-1β, IL-6, COX-2, and iONS ↓ | Inhibit the inflammatory response and regulate the GM. | [20] |
| CRC | / | Genus Klebsiella ↑ Abundances of Klebsiella ↑ Abundance of phylum Proteobacteria ↓ | / | There are differences in intestinal microflora between patients with colorectal cancer and normal people after treatment, and the microbial diversity is reduced, which makes them more sensitive. | [21] |
| Obesity | Total Flavonoids of Quzhou Fructus Aurantii Extract | Genera Akkermansia, Alistipes ↑ Firmicutes to Bacteroidetes ratio; Genera Dubosiella, Faecalibaculum, and Lactobacillus ↓ | Reduced obesity, inflammation, and liver steatosis TC, TG, and OTGG ↓ Phospho-P65, phospho-IKKα/β, TNF-α, and COX-2 ↓ | Utilizing prebiotics as dietary supplements to regulate the GM. | [22] |
| Obesity | Anthocyanin Monomer from Lycium ruthenicum Murray. Fruit | Abundances of Bifidobacteriaceae, Helicobacteraceae, and Deferribacteraceae ↑ Abundance of Firmicutes, Lactobacillaceae, Streptococcaceae, Ruminococcaceae, and Erysipelotrichaceae ↓ | Control the increase in fat and weight. ALT, AST, TG, and LDL-C ↓ LPS, IL-6, and IL-1β ↓ | Anthocyanins can maintain the integrity of intestinal barrier and regulate GM. | [23] |
| Diagnosed Diabetics | / | Lactobacillus ↑ Abundance of Megasphaera, Escherichia, and Acidaminococcus ↑ Akkermansia, Blautia, and Ruminococcus ↓ Abundance of Sutterella ↓ | LPS and IL-6 ↑ | The intestinal flora of diabetic patients at different stages tends to recover to normal people after treatment. | [24] |
| Diagnosed Diabetics | Blautia obeum and Blautia wexlerae ↑ Bacteroides dorei, Coprococcus eutactus, Akkermansia muciniphila, and Bacteroides spp. ↓ | 3,8-dihydroxy-urolithin (urolithin A), phenyl-γ-valerolactones, and various phenolic acid concentrations ↓ | The change of microbial composition is closely related to diabetes control. | [25] | |
| AD | / | Escherichia and Enterococcus ↑ Lactobacillus, Bifidobacterium, and Ruminococcus ↓ | TNF-α and IL-6 ↑ | The abundance of pro-inflammatory bacteria increases, thus releasing inflammatory factors. | [26] |
| PD | Probiotic Supplement | Christensenella spp. and Marseille-P2437 ↑ g_Eubacterium_oxidoreducens_group, g_Eubacterium_hallii_group, and s_Odoribacter_sp._N54.MGS-14 ↓ | / | Probiotics treatment can effectively improve the constipation symptoms of PD patients and positively affect the GM. | [27] |
| Autism Spectrum Disorders | / | Firmicutes, Actinobacteria, Bacteroidetes, and Proteobacteria ↑ Faecalibacterium, Bacteroides, Prevotella_9, Blautia, and Subdoligranulum ↑ | / | Gastrointestinal symptoms are positively correlated with autism symptoms, among which constipation is the most common. | [28] |
| Bioactives | Fruit | Microbiota Influenced | Effect on Health | Reference |
|---|---|---|---|---|
| Polysaccharide | Aronia melanocarpa | Bacteroides phylum ↑ Firmicutes ↓ | Alleviated inflammation and oxidative stress injury in aging brain tissue | [44] |
| Polysaccharides | Noni (Morinda citrifolia L.) | Enterobacteriales, BetaProteobacteriales, Verrucomicrobiales ↑ Gram-positive bacteria ↓ | Promote gut mucosal barrier; improve the expression of gut SCFAs | [19] |
| Polysaccharide | Pitaya flower buds | Lachnospiraceae, Ruminococcaceae ↑ Muribaculaceae, Lactobacillaceae ↑ | Alleviate obesity; prevent gut atrophy | [45] |
| Oligosaccharides | Dragon fruit | Flavobacterium, Flectobacillus, Acidovorax ↑ Leptolyngbya, Pseudomonas ↓ | Improve immunity; reduce lipid peroxidation and oxidative stress | [46] |
| Oligosaccharide | Mulberry | Prevotellaceae, Lactobacillus ↑ Lachnospiraceae ↓ | Decrease the blood glucose | [47] |
| Polyphenols | Pomegranate fruit | Bacteroidetes, A. muciniphila, ParaBacteroides distasonis, Bacteroides acidifaciens, Mucispirillum schaedleri, Lachnospiraceae ↑ Firmicutes, Proteobacteria ↓ | Alleviated obesity; insulin resistance and hepatic steatosis | [9] |
| Polyphenol | Blackberry | Firmicutes/Bacteroidetes ↓ | Anti-obese | [48] |
| Anthocyanin | Açaí (Euterpe oleracea Mart.) fruit | A. muciniphila ↑ | Anti-obesity; alleviate liver steatosis; insulin resistance | [49] |
| Flavonoids | Rosa davurica Pall. fruits | Erysipelotrichaceae ↓ | Anti-obesity; restoring liver function; reverse gut dysbiosis | [50] |
| Flavonoids | Passiflora foetida fruits | Bifidobacterium ↑ Enterococcus, Lactobacillus, Roseburia ↑ Alistipes, Bilophia, Enterobacteriaceae ↓ | Anti-inflammatory and GM modulation | [51] |
| Active Ingredients | Category | Representative Compound | Gut Microbes Involved in Metabolism | Metabolite Transformation | Bioavailability | Reference |
|---|---|---|---|---|---|---|
| Polysaccharide | Malus pumila polysaccharide | Starch | Bifidobacteria adolescentis, Ruminococcus bromii | SCFAs (mainly butyrate) | / | [95] |
| Pectin | Bacteroides, Lactobacillus | SCFAs (mainly acetic acid, propionic acid, butyrate) | / | [96,97] | ||
| Oligosaccharide | Fructooligosaccharide | / | Lactobacillus, Clostridium | SCFAs, carbon dioxide, methane, hydrogen | 60% | [98,99] |
| Polyphenol | Ellagitannins | Sanguiin H6 | Coriobacterium, Lactobacillus | Urolithins | / | [100,101] |
| Agrimoniin | Eggerthellaceae, Bacillus gordonae sp. | Urolithins | / | [102,103] | ||
| Phenolic acid | Gallic acid | Proteobacteria, Firmicutes | Pyrogallic acid | / | [104] | |
| Chlorogenic acid | Bifidobacterium, Bacillus | Coumaric acid, benzoic acid, phenylpropionic acid | 30% | [105] | ||
| Flavonoid | Flavonol | Quercetin | Streptococcus lutei, Lactobacillus acidophilus, Clostridium, Bacteroides fragilis, Clostridium | Quercetin 3-glucoside, quercetin 7-glucoside | 36–53% | [106,107] |
| Rutin | Bacteroides, Clostridium, Bacillus | 3, 4-dihydroxyphenylacetic acid, rapesin, quercetin-3-O-glucoside, phloroglucinol | 22% | [108] | ||
| Flavanone | Hesperetin | Bacteroides, Clostridium, Bifidobacterium | 7-O-glucuronic acid, hesperidin-7-O-hesperidin sulfate | / | [109,110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Y.; Chen, L.; Yu, Y.; Hussain, M.; Zhong, H. Bioactive Components in Fruit Interact with Gut Microbes. Biology 2023, 12, 1333. https://doi.org/10.3390/biology12101333
Jin Y, Chen L, Yu Y, Hussain M, Zhong H. Bioactive Components in Fruit Interact with Gut Microbes. Biology. 2023; 12(10):1333. https://doi.org/10.3390/biology12101333
Chicago/Turabian StyleJin, Yuanyuan, Ling Chen, Yufen Yu, Muhammad Hussain, and Hao Zhong. 2023. "Bioactive Components in Fruit Interact with Gut Microbes" Biology 12, no. 10: 1333. https://doi.org/10.3390/biology12101333