
Dysregulated Cyclic Nucleotide Metabolism in Alcohol-Associated Steatohepatitis: Implications for Novel Targeted Therapies
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Study
2.2. Animal Experiments
2.3. Measurement of Liver Injury Markers
2.4. Cytokine Measurement by MSD Platform
2.5. MPO Activity Assay
2.6. Measurements of cAMP and cGMP Levels
2.7. Liver Histopathology and Sirius Red Staining
2.8. Statistical Analysis
2.9. RNA Sequencing and Statistical Analysis
3. Results
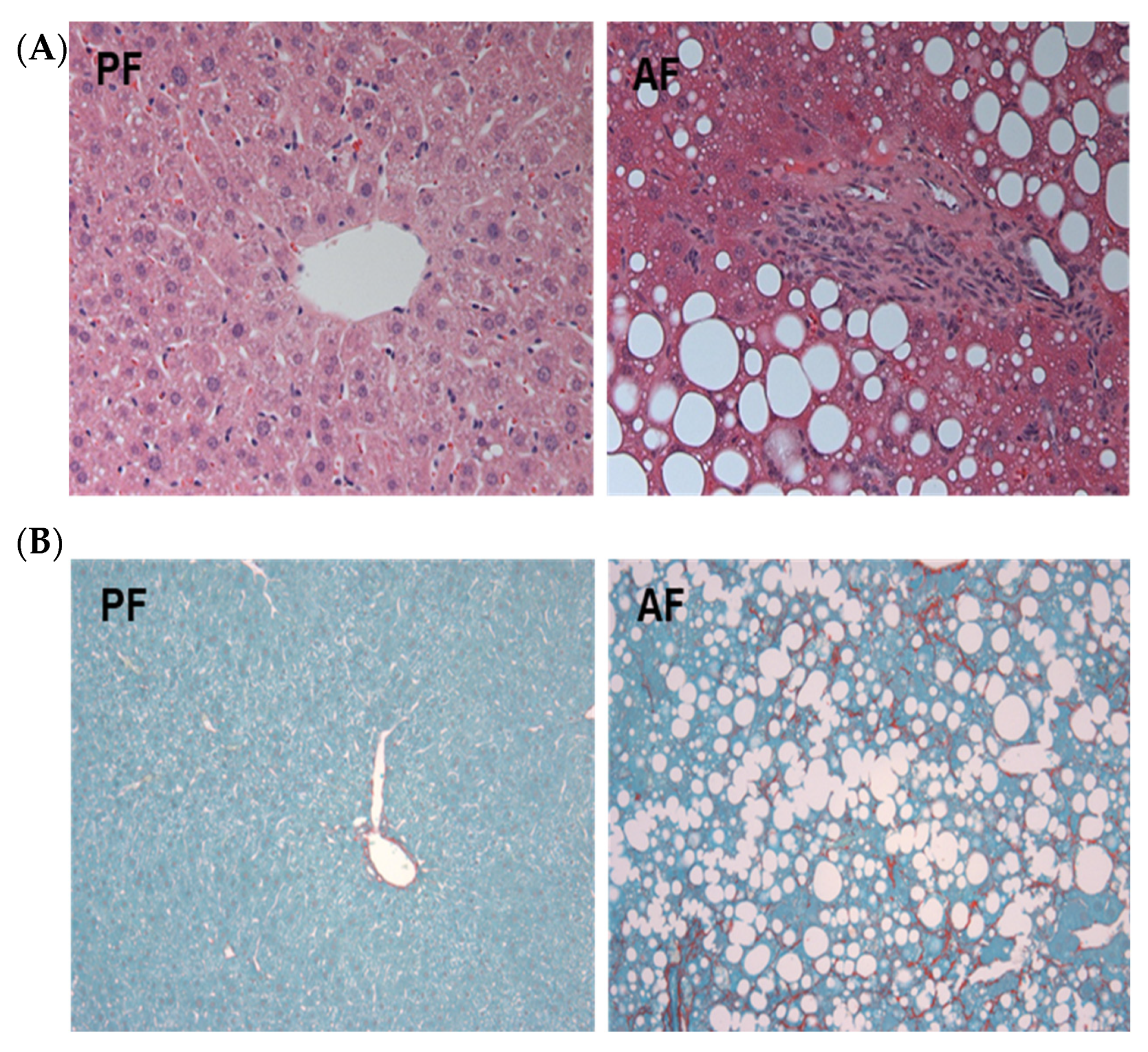
3.1. Mice Developed Severe Liver Injury, Inflammation, Steatosis, and Pericellular Fibrosis in the ASH Model
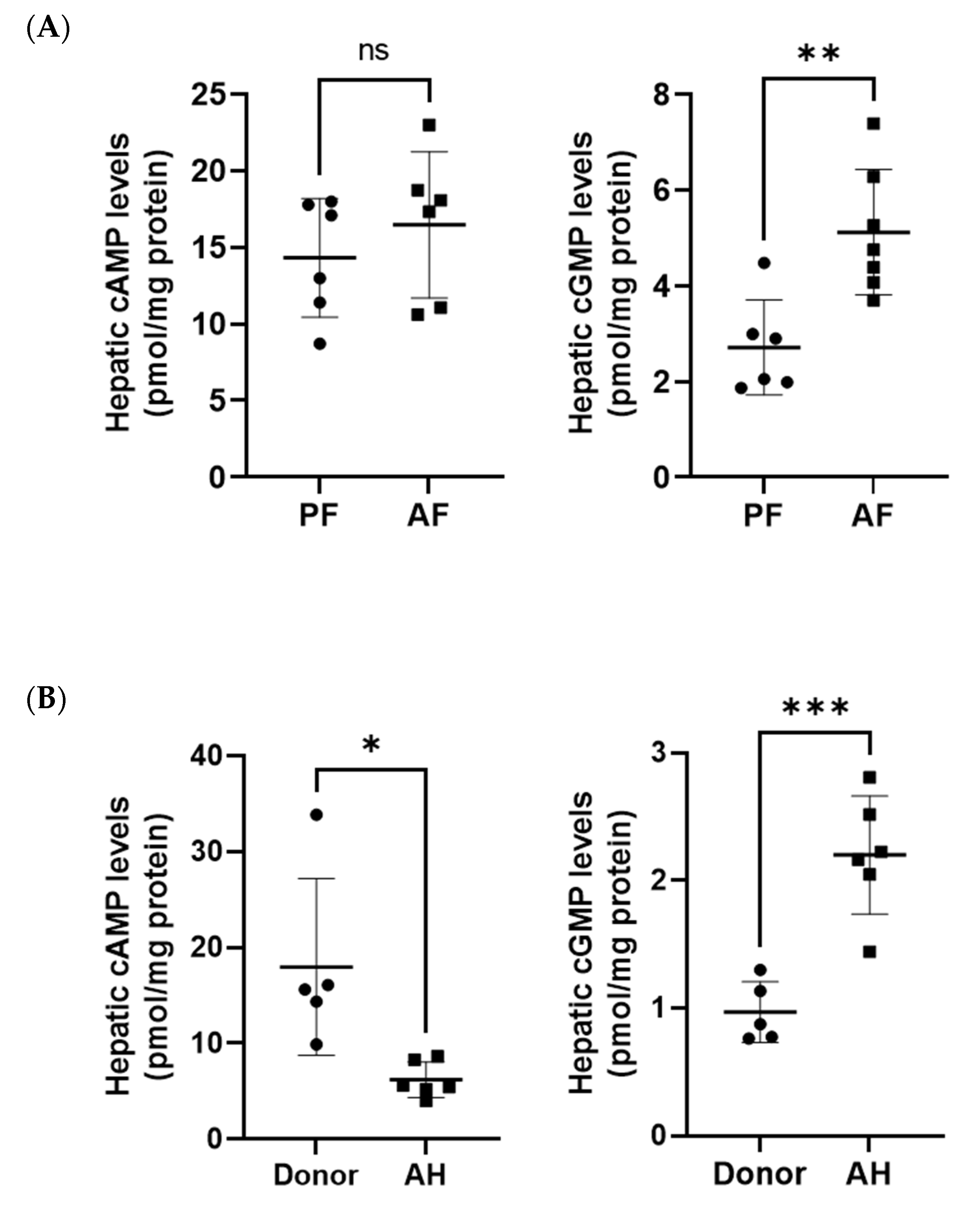
3.2. Hepatic Cyclic AMP and Cyclic GMP Levels in ASH Mice and Patients with AH
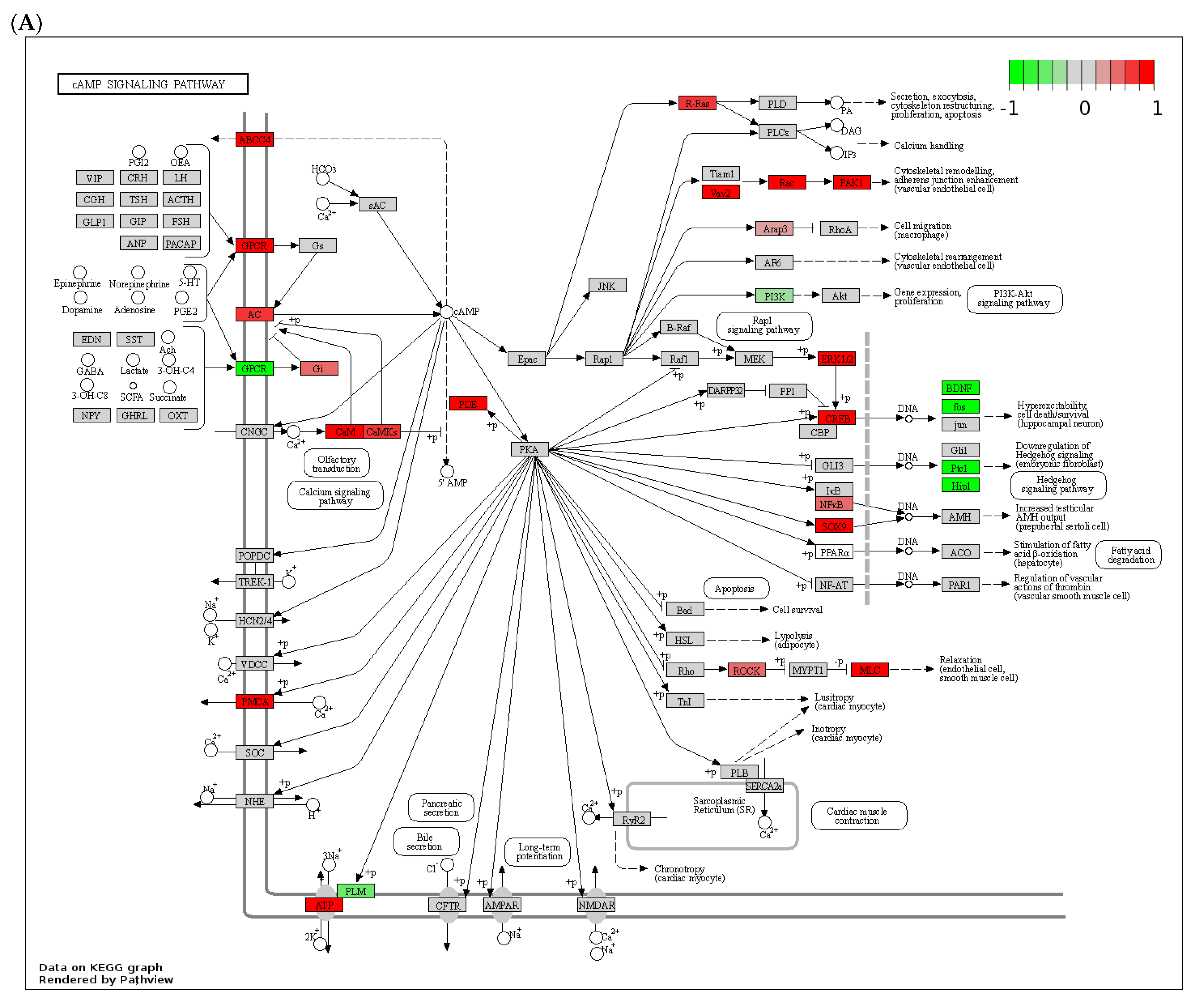
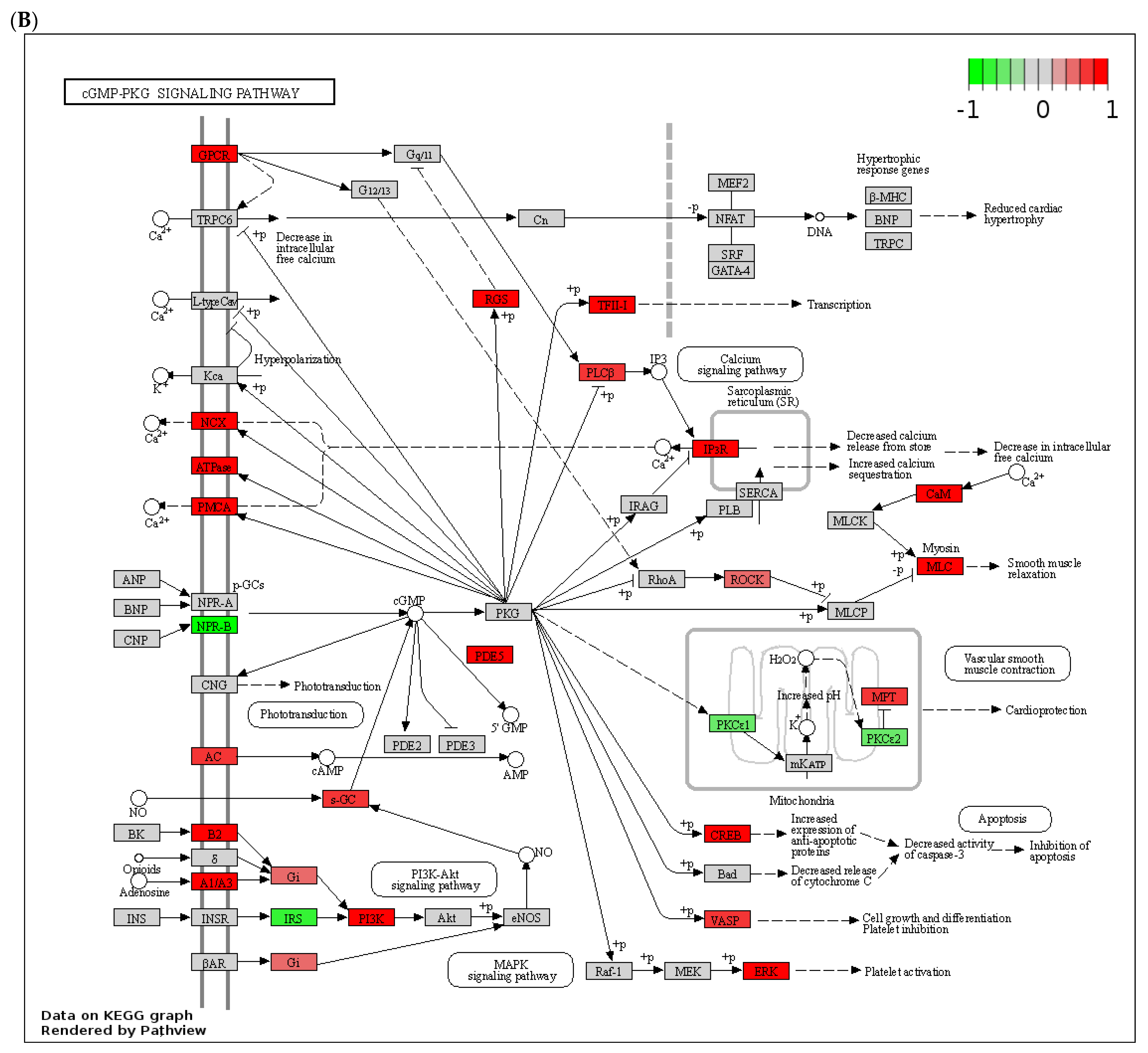
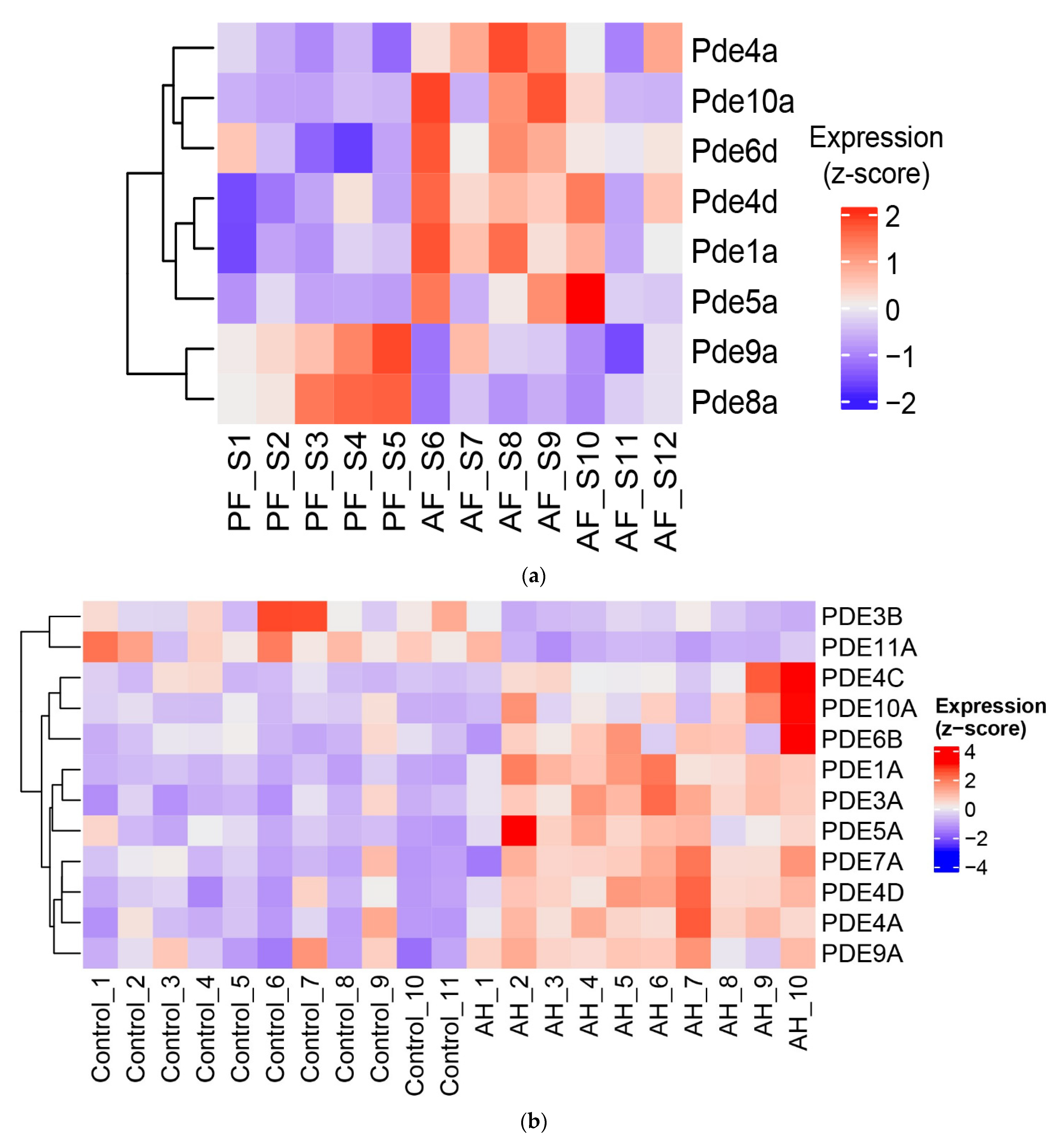
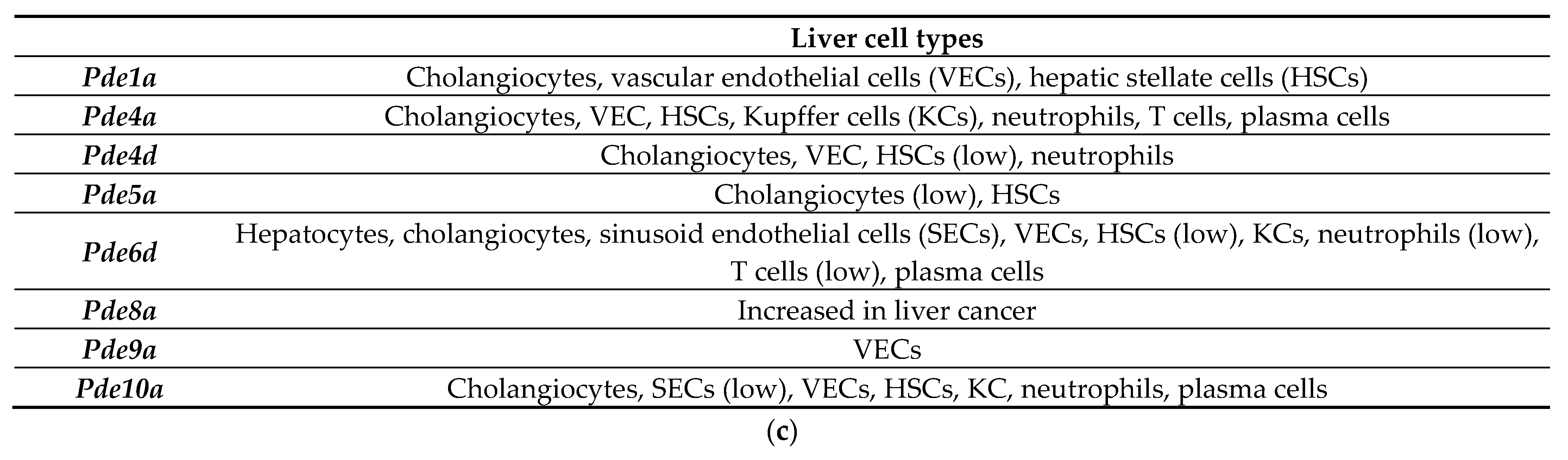
3.3. Dysregulated Expression of Several Phosphodiesterases by Alcohol in the Livers of ASH Mice and Patients with Alcohol-Associated Hepatitis
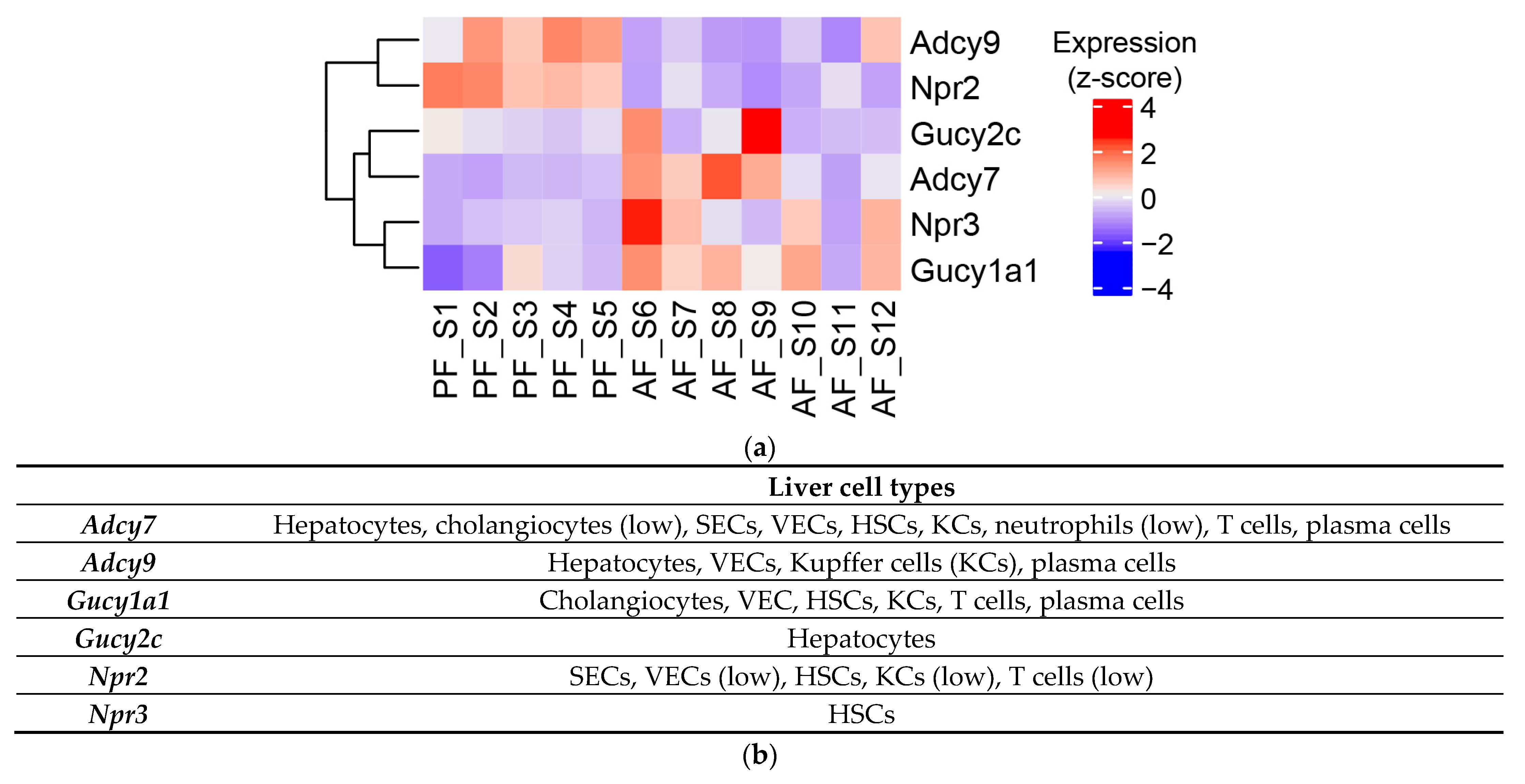
3.4. Effect of Chronic Alcohol Feeding on Adenylyl and Guanylyl Cyclases
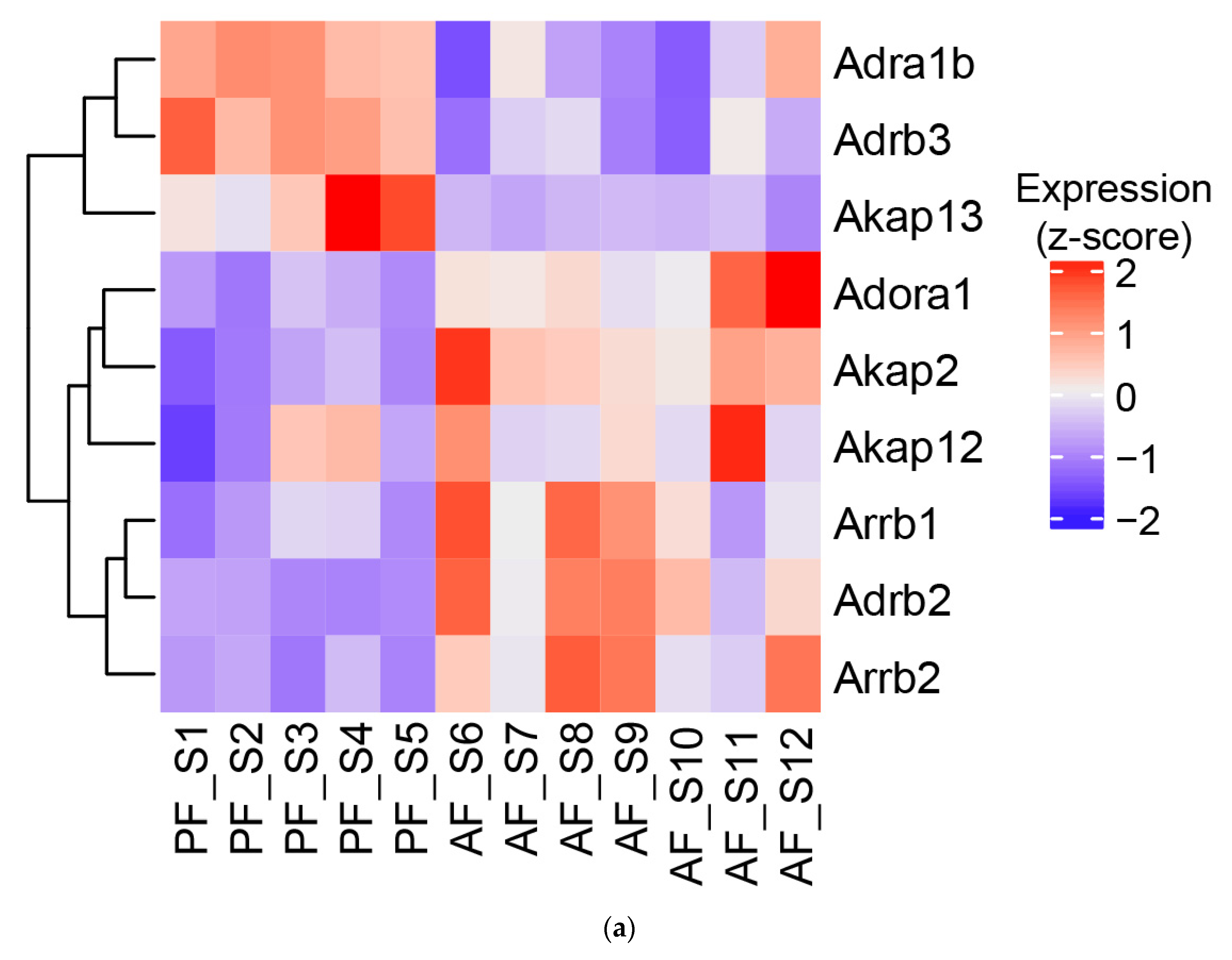
3.5. Effect of Alcohol on G-Protein-Coupled Receptor Expression, A-Kinase Anchoring Proteins (AKAPs), and β-Arrestins
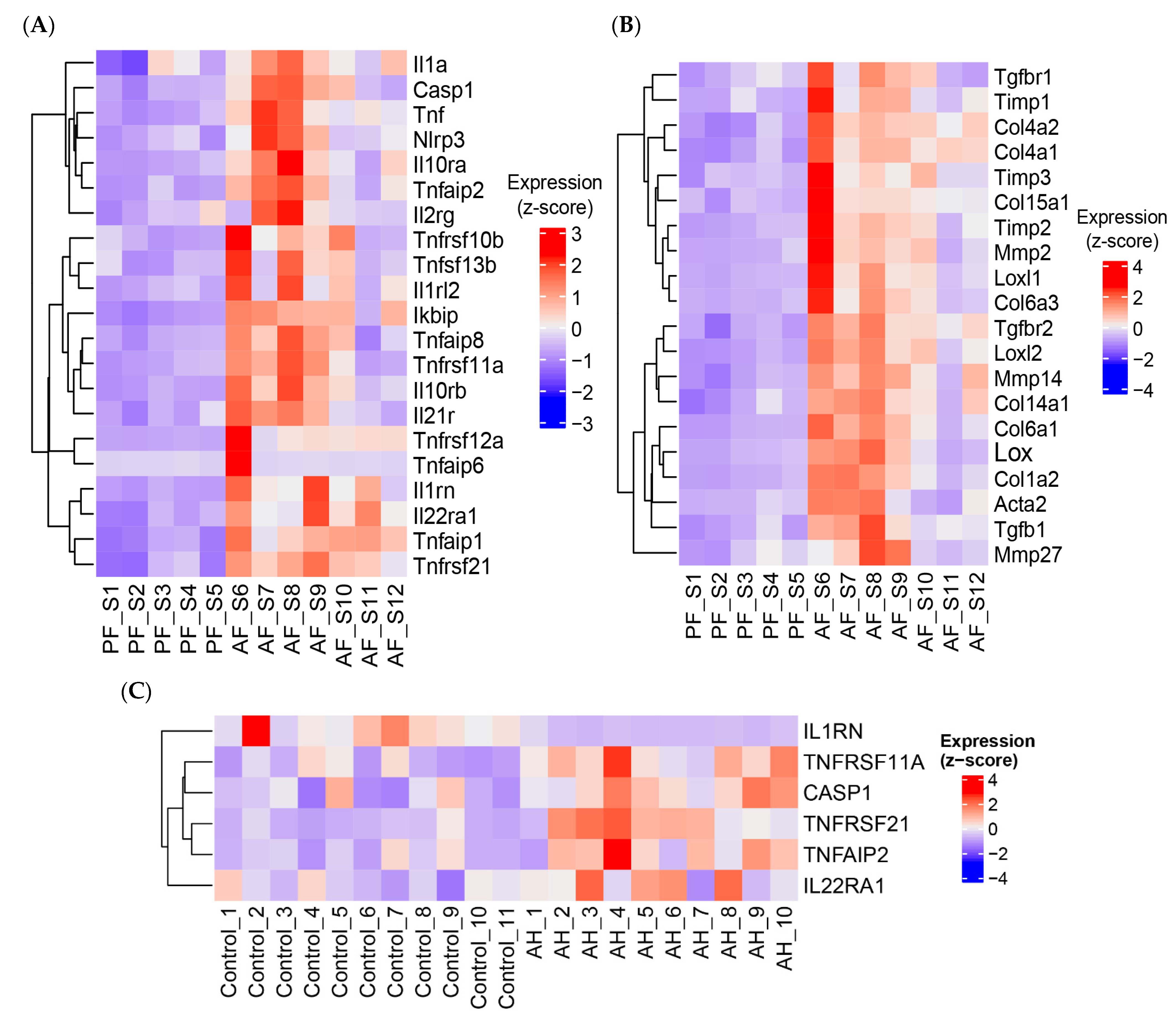
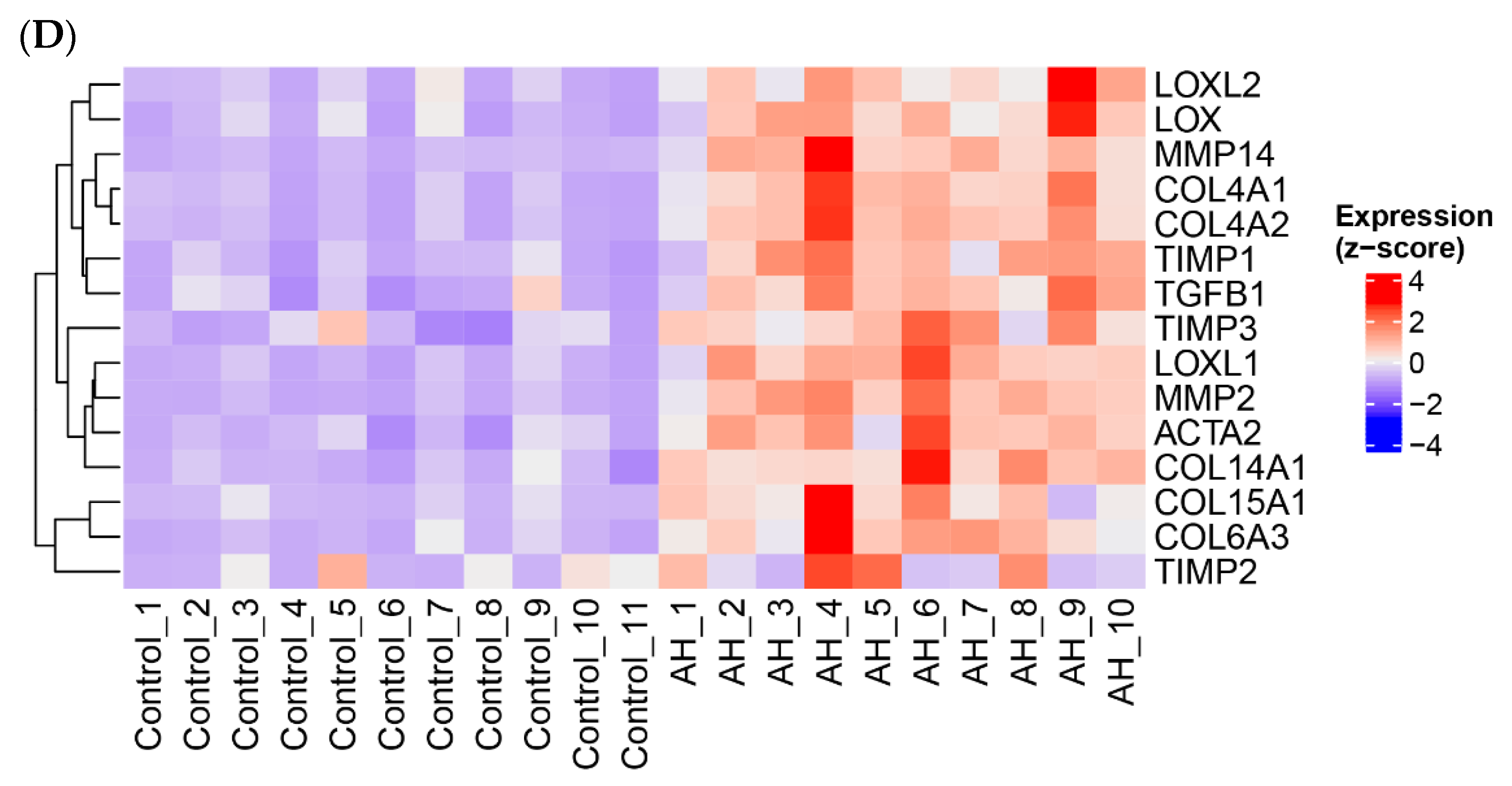
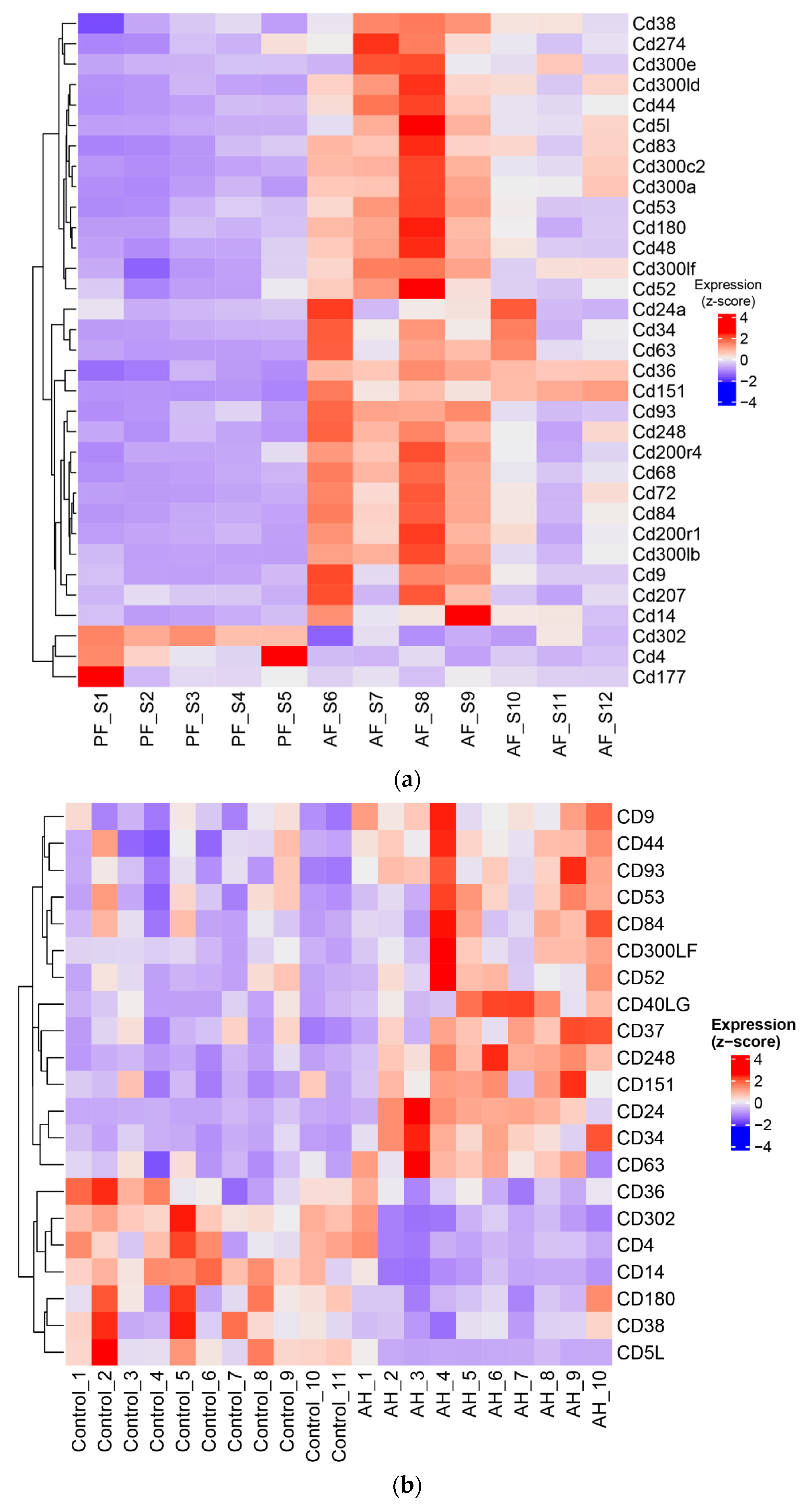
3.6. ASH/AH Pathogenesis Is Associated with Significant Changes in Cellular Markers Signifying the Changes in Cell Types and Stage of Differentiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deutsch-Link, S.; Jiang, Y.; Peery, A.F.; Barritt, A.S.; Bataller, R.; Moon, A.M. Alcohol-Associated Liver Disease Mortality Increased from 2017 to 2020 and Accelerated during the COVID-19 Pandemic. Clin. Gastroenterol. Hepatol. 2022, 20, 2142–2144.e2. [Google Scholar] [CrossRef]
- Bataller, R.; Arab, J.P.; Shah, V.H. Alcohol-Associated Hepatitis. N. Engl. J. Med. 2022, 387, 2436–2448. [Google Scholar] [CrossRef]
- Osna, N.A.; Rasineni, K.; Ganesan, M.; Donohue, T.M., Jr.; Kharbanda, K.K. Pathogenesis of Alcohol-Associated Liver Disease. J. Clin. Exp. Hepatol. 2022, 12, 1492–1513. [Google Scholar] [CrossRef]
- Singal, A.K.; Mathurin, P. Diagnosis and Treatment of Alcohol-Associated Liver Disease: A Review. JAMA 2021, 326, 165–176. [Google Scholar] [CrossRef]
- Zaccolo, M.; Zerio, A.; Lobo, M.J. Subcellular Organization of the cAMP Signaling Pathway. Pharmacol. Rev. 2021, 73, 278–309. [Google Scholar] [CrossRef]
- Friebe, A.; Sandner, P.; Schmidtko, A. cGMP: A unique 2nd messenger molecule—Recent developments in cGMP research and development. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 287–302. [Google Scholar] [CrossRef]
- Sadana, R.; Dessauer, C.W. Physiological roles for G protein-regulated adenylyl cyclase isoforms: Insights from knockout and overexpression studies. Neurosignals 2009, 17, 5–22. [Google Scholar] [CrossRef]
- Buglioni, A.; Burnett, J.C., Jr. New Pharmacological Strategies to Increase cGMP. Annu. Rev. Med. 2016, 67, 229–243. [Google Scholar] [CrossRef]
- Potter, L.R. Guanylyl cyclase structure, function and regulation. Cell. Signal. 2011, 23, 1921–1926. [Google Scholar] [CrossRef]
- Hanoune, J.; Defer, N. Regulation and role of adenylyl cyclase isoforms. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 145–174. [Google Scholar] [CrossRef]
- Breckler, M.; Berthouze, M.; Laurent, A.C.; Crozatier, B.; Morel, E.; Lezoualc’h, F. Rap-linked cAMP signaling Epac proteins: Compartmentation, functioning and disease implications. Cell. Signal. 2011, 23, 1257–1266. [Google Scholar] [CrossRef]
- Brand, T.; Schindler, R. New kids on the block: The Popeye domain containing (POPDC) protein family acting as a novel class of cAMP effector proteins in striated muscle. Cell. Signal. 2017, 40, 156–165. [Google Scholar] [CrossRef]
- Kaupp, U.B.; Seifert, R. Cyclic nucleotide-gated ion channels. Physiol. Rev. 2002, 82, 769–824. [Google Scholar] [CrossRef]
- Franca, M.E.R.; Peixoto, C.A. cGMP signaling pathway in hepatic encephalopathy neuroinflammation and cognition. Int. Immunopharmacol. 2020, 79, 106082. [Google Scholar] [CrossRef]
- Francis, S.H.; Busch, J.L.; Corbin, J.D.; Sibley, D. cGMP-dependent protein kinases and cGMP phosphodiesterases in nitric oxide and cGMP action. Pharmacol. Rev. 2010, 62, 525–563. [Google Scholar] [CrossRef]
- Bolger, G.B. The PDE-Opathies: Diverse Phenotypes Produced by a Functionally Related Multigene Family. Trends Genet. 2021, 37, 669–681. [Google Scholar] [CrossRef]
- Elnagdy, M.; Barve, S.; McClain, C.; Gobejishvili, L. cAMP Signaling in Pathobiology of Alcohol Associated Liver Disease. Biomolecules 2020, 10, 1433. [Google Scholar] [CrossRef]
- Wahlang, B.; McClain, C.; Barve, S.; Gobejishvili, L. Role of cAMP and phosphodiesterase signaling in liver health and disease. Cell. Signal. 2018, 49, 105–115. [Google Scholar] [CrossRef]
- Avila, D.V.; Barker, D.F.; Zhang, J.; McClain, C.J.; Barve, S.; Gobejishvili, L. Dysregulation of hepatic cAMP levels via altered Pde4b expression plays a critical role in alcohol-induced steatosis. J. Pathol. 2016, 240, 96–107. [Google Scholar] [CrossRef]
- Rodriguez, W.E.; Wahlang, B.; Wang, Y.; Zhang, J.; Vadhanam, M.V.; Joshi-Barve, S.; Bauer, P.; Cannon, R.; Ahmadi, A.R.; Sun, Z.; et al. Phosphodiesterase 4 Inhibition as a Therapeutic Target for Alcoholic Liver Disease: From Bedside to Bench. Hepatology 2019, 70, 1958–1971. [Google Scholar] [CrossRef]
- Gobejishvili, L.; Barve, S.; Joshi-Barve, S.; McClain, C. Enhanced PDE4B expression augments LPS-inducible TNF expression in ethanol-primed monocytes: Relevance to alcoholic liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G718–G724. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, F.; Wu, X.; Lv, X.; Li, J. EPAC activation inhibits acetaldehyde-induced activation and proliferation of hepatic stellate cell via Rap1. Can. J. Physiol. Pharmacol. 2016, 94, 498–507. [Google Scholar] [CrossRef]
- Lazaro, R.; Wu, R.; Lee, S.; Zhu, N.L.; Chen, C.L.; French, S.W.; Xu, J.; Machida, K.; Tsukamoto, H. Osteopontin deficiency does not prevent but promotes alcoholic neutrophilic hepatitis in mice. Hepatology 2015, 61, 129–140. [Google Scholar] [CrossRef]
- Massey, V.; Parrish, A.; Argemi, J.; Moreno, M.; Mello, A.; García-Rocha, M.; Altamirano, J.; Odena, G.; Dubuquoy, L.; Louvet, A.; et al. Integrated Multiomics Reveals Glucose Use Reprogramming and Identifies a Novel Hexokinase in Alcoholic Hepatitis. Gastroenterology 2021, 160, 1725–1740.e2. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 February 2019).
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Luo, W.; Pant, G.; Bhavnasi, Y.K.; Blanchard Jr, S.G.; Brouwer, C. Pathview Web: User friendly pathway visualization and data integration. Nucleic Acids Res. 2017, 45, W501–W508. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Azevedo, M.F.; Faucz, F.R.; Bimpaki, E.; Horvath, A.; Levy, I.; de Alexandre, R.B.; Ahmad, F.; Manganiello, V.; Stratakis, C.A. Clinical and molecular genetics of the phosphodiesterases (PDEs). Endocr. Rev. 2014, 35, 195–233. [Google Scholar] [CrossRef]
- Peng, Z.; Borea, P.A.; Varani, K.; Wilder, T.; Yee, H.; Chiriboga, L.; Blackburn, M.R.; Azzena, G.; Resta, G.; Cronstein, B.N. Adenosine signaling contributes to ethanol-induced fatty liver in mice. J. Clin. Investig. 2009, 119, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Tiwari, N.; Madar, J.; Mehta, P.; Qiao, L.Y. Beta 2-adrenergic receptor mediates noradrenergic action to induce cyclic adenosine monophosphate response element-binding protein phosphorylation in satellite glial cells of dorsal root ganglia to regulate visceral hypersensitivity. Pain 2022, 163, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.P.; Hohl, C.; Altschuld, R.; Jones, L.; Livingston, B.; Ziman, B.; Tantini, B.; Lakatta, E.G. Beta 2-adrenergic receptor-stimulated increase in cAMP in rat heart cells is not coupled to changes in Ca2+ dynamics, contractility, or phospholamban phosphorylation. J. Biol. Chem. 1994, 269, 19151–19156. [Google Scholar] [CrossRef]
- Flacco, N.; Segura, V.; Perez-Aso, M.; Estrada, S.; Seller, J.F.; Jimenez-Altayo, F.; Noguera, M.A.; D’Ocon, P.; Vila, E.; Ivorra, M.D. Different beta-adrenoceptor subtypes coupling to cAMP or NO/cGMP pathways: Implications in the relaxant response of rat conductance and resistance vessels. Br. J. Pharmacol. 2013, 169, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Osawa, Y.; Kawai, H.; Tsunoda, T.; Komatsu, H.; Okawara, M.; Tsutsui, Y.; Yoshida, Y.; Yoshikawa, S.; Mori, T.; Yamazoe, T.; et al. Cluster of Differentiation 44 Promotes Liver Fibrosis and Serves as a Biomarker in Congestive Hepatopathy. Hepatol. Commun. 2021, 5, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Hochst, B.; Schildberg, F.A.; Sauerborn, P.; Gabel, Y.A.; Gevensleben, H.; Goltz, D.; Heukamp, L.C.; Turler, A.; Ballmaier, M.; Gieseke, F.; et al. Activated human hepatic stellate cells induce myeloid derived suppressor cells from peripheral blood monocytes in a CD44-dependent fashion. J. Hepatol. 2013, 59, 528–535. [Google Scholar] [CrossRef]
- Reinecke, B.; Frericks, N.; Lauber, C.; Dinkelborg, K.; Matthaei, A.; Vondran, F.W.R.; Behrendt, P.; Haid, S.; Brown, R.J.P.; Pietschmann, T. The Human Liver-Expressed Lectin CD302 Restricts Hepatitis C Virus Infection. J. Virol. 2022, 96, e0199521. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, W.; Xing, W.; You, T.; Xu, J.; Qin, X.; Peng, Z. The protective role of CD59 and pathogenic role of complement in hepatic ischemia and reperfusion injury. Am. J. Pathol. 2011, 179, 2876–2884. [Google Scholar] [CrossRef]
- Ma, H.; Wang, C.; Liang, S.; Yu, X.; Yuan, Y.; Lv, Z.; Zhang, J.; Jin, C.; Zhu, J.; Wang, C.; et al. ROCK inhibition enhanced hepatocyte liver engraftment by retaining membrane CD59 and attenuating complement activation. Mol. Ther. 2023, 31, 1846–1856. [Google Scholar] [CrossRef]
- Diestelhorst, J.; Junge, N.; Schlue, J.; Falk, C.S.; Manns, M.P.; Baumann, U.; Jaeckel, E.; Taubert, R. Pediatric autoimmune hepatitis shows a disproportionate decline of regulatory T cells in the liver and of IL-2 in the blood of patients undergoing therapy. PLoS ONE 2017, 12, e0181107. [Google Scholar] [CrossRef]
- Fuentes-Panana, E.M.; Bannish, G.; Karnell, F.G.; Treml, J.F.; Monroe, J.G. Analysis of the individual contributions of Igalpha (CD79a)- and Igbeta (CD79b)-mediated tonic signaling for bone marrow B cell development and peripheral B cell maturation. J. Immunol. 2006, 177, 7913–7922. [Google Scholar] [CrossRef] [PubMed]
- Calvente, C.J.; Tameda, M.; Johnson, C.D.; Del Pilar, H.; Lin, Y.C.; Adronikou, N.; De Mollerat Du Jeu, X.; Llorente, C.; Boyer, J.; Feldstein, A.E. Neutrophils contribute to spontaneous resolution of liver inflammation and fibrosis via microRNA-223. J. Clin. Investig. 2019, 129, 4091–4109. [Google Scholar] [CrossRef] [PubMed]
- Engering, A.; Pieters, J. Association of distinct tetraspanins with MHC class II molecules at different subcellular locations in human immature dendritic cells. Int. Immunol. 2001, 13, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.C.; van Spriel, A.B.; Gartlan, K.H.; Sofi, M.; Apostolopoulos, V.; Ashman, L.; Wright, M.D. Tetraspanins CD37 and CD151 differentially regulate Ag presentation and T-cell co-stimulation by DC. Eur. J. Immunol. 2009, 39, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.G.; Tran, J.L.; Erion, D.M.; Vera, N.B.; Febbraio, M.; Weiss, E.J. Hepatocyte-Specific Disruption of CD36 Attenuates Fatty Liver and Improves Insulin Sensitivity in HFD-Fed Mice. Endocrinology 2016, 157, 570–585. [Google Scholar] [CrossRef]
- Maehara, N.; Arai, S.; Mori, M.; Iwamura, Y.; Kurokawa, J.; Kai, T.; Kusunoki, S.; Taniguchi, K.; Ikeda, K.; Ohara, O.; et al. Circulating AIM prevents hepatocellular carcinoma through complement activation. Cell Rep. 2014, 9, 61–74. [Google Scholar] [CrossRef]
- Barlas, N.; Mutchnick, M.G.; Grant, G.J.; Trainin, N. The effect of thymic humoral factor on intracellular lymphocyte cyclic AMP in alcoholic liver disease. Thymus 1983, 5, 433–437. [Google Scholar]
- Diamond, I.; Wrubel, B.; Estrin, W.; Gordon, A. Basal and adenosine receptor-stimulated levels of cAMP are reduced in lymphocytes from alcoholic patients. Proc. Natl. Acad. Sci. USA 1987, 84, 1413–1416. [Google Scholar] [CrossRef]
- Mochly-Rosen, D.; Chang, F.H.; Cheever, L.; Kim, M.; Diamond, I.; Gordon, A.S. Chronic ethanol causes heterologous desensitization of receptors by reducing alpha s messenger RNA. Nature 1988, 333, 848–850. [Google Scholar] [CrossRef]
- Nagy, L.E.; DeSilva, S.E. Ethanol increases receptor-dependent cyclic AMP production in cultured hepatocytes by decreasing G(i)-mediated inhibition. Biochem. J. 1992, 286 Pt 3, 681–686. [Google Scholar] [CrossRef]
- Gobejishvili, L.; Barve, S.; Joshi-Barve, S.; Uriarte, S.; Song, Z.; McClain, C. Chronic ethanol-mediated decrease in cAMP primes macrophages to enhanced LPS-inducible NF-kappaB activity and TNF expression: Relevance to alcoholic liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G681–G688. [Google Scholar] [CrossRef] [PubMed]
- Gouillon, Z.Q.; Miyamoto, K.; Donohue, T.M.; Wan, Y.J.; French, B.A.; Nagao, Y.; Fu, P.; Reitz, R.C.; Hagbjork, A.; Yap, C.; et al. Role of CYP2E1 in the pathogenesis of alcoholic liver disease: Modifications by cAMP and ubiquitin-proteasome pathway. Front. Biosci. 1999, 4, A16–A25. [Google Scholar] [CrossRef] [PubMed]
- Diehl, A.M.; Yang, S.Q.; Cote, P.; Wand, G.S. Chronic ethanol consumption disturbs G-protein expression and inhibits cyclic AMP-dependent signaling in regenerating rat liver. Hepatology 1992, 16, 1212–1219. [Google Scholar]
- Tabakoff, B.; Hoffman, P.L. The role of the type 7 adenylyl cyclase isoform in alcohol use disorder and depression. Front. Pharmacol. 2022, 13, 1012013. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Pohli, S.; Durany, N.; Ozawa, H.; Saito, T.; Boissl, K.W.; Zochling, R.; Riederer, P.; Boning, J.; Gotz, M.E. Increased levels of calcium-sensitive adenylyl cyclase subtypes in the limbic system of alcoholics: Evidence for a specific role of cAMP signaling in the human addictive brain. Brain Res. 2001, 895, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Gobejishvili, L.; Barve, S.; Breitkopf-Heinlein, K.; Li, Y.; Zhang, J.; Avila, D.V.; Dooley, S.; McClain, C.J. Rolipram attenuates bile duct ligation-induced liver injury in rats: A potential pathogenic role of PDE4. J. Pharmacol. Exp. Ther. 2013, 347, 80–90. [Google Scholar] [CrossRef]
- Elnagdy, M.; Wang, Y.; Rodriguez, W.; Zhang, J.; Bauer, P.; Wilkey, D.W.; Merchant, M.; Pan, J.; Farooqui, Z.; Cannon, R.; et al. Increased expression of phosphodiesterase 4 in activated hepatic stellate cells promotes cytoskeleton remodeling and cell migration. J. Pathol. 2023, 261, 361–371. [Google Scholar] [CrossRef]
- Mukherjee, D.; Previs, R.A.; Haines, C.; Al Abo, M.; Juras, P.K.; Strickland, K.C.; Chakraborty, B.; Artham, S.; Whitaker, R.S.; Hebert, K.; et al. Targeting CaMKK2 inhibits actin cytoskeletal assembly to suppress cancer metastasis. Cancer Res. 2023, 83, 2889–2907. [Google Scholar] [CrossRef]
- Tao, X.; He, H.; Peng, J.; Xu, R.; Fu, J.; Hu, Y.; Li, L.; Yang, X.; Feng, X.; Zhang, C.; et al. Overexpression of PDE4D in mouse liver is sufficient to trigger NAFLD and hypertension in a CD36-TGF-β1 pathway: Therapeutic role of roflumilast. Pharmacol. Res. 2022, 175, 106004. [Google Scholar] [CrossRef]
- Mishra, S.; Sadagopan, N.; Dunkerly-Eyring, B.; Rodriguez, S.; Sarver, D.C.; Ceddia, R.P.; Murphy, S.A.; Knutsdottir, H.; Jani, V.P.; Ashok, D.; et al. Inhibition of phosphodiesterase type 9 reduces obesity and cardiometabolic syndrome in mice. J. Clin. Investig. 2021, 131, e148798. [Google Scholar] [CrossRef]
- Ceddia, R.P.; Liu, D.; Shi, F.; Crowder, M.K.; Mishra, S.; Kass, D.A.; Collins, S. Increased Energy Expenditure and Protection From Diet-Induced Obesity in Mice Lacking the cGMP-Specific Phosphodiesterase PDE9. Diabetes 2021, 70, 2823–2836. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Q.; Jiang, M.Y.; Huang, Y.Y.; Zhu, Z.; Han, C.; Tian, Y.J.; Zhang, B.; Luo, H.B. Discovery of Potent Phosphodiesterase-9 Inhibitors for the Treatment of Hepatic Fibrosis. J. Med. Chem. 2021, 64, 9537–9549. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Shi, J.R.; Li, S.C.; Wang, L.M.; Dhar, R.; Li, N.; Cao, X.W.; Li, Z.G.; Tang, H.F. Phosphodiesterase type 10A inhibitor attenuates lung fibrosis by targeting myofibroblast activation. iScience 2023, 26, 106586. [Google Scholar] [CrossRef] [PubMed]
- Kisseleva, T.; Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Wehbi, V.L.; Tasken, K. Molecular Mechanisms for cAMP-Mediated Immunoregulation in T cells—Role of Anchored Protein Kinase A Signaling Units. Front. Immunol. 2016, 7, 222. [Google Scholar] [CrossRef]
- Negreiros-Lima, G.L.; Lima, K.M.; Moreira, I.Z.; Jardim, B.L.O.; Vago, J.P.; Galvao, I.; Teixeira, L.C.R.; Pinho, V.; Teixeira, M.M.; Sugimoto, M.A.; et al. Cyclic AMP Regulates Key Features of Macrophages via PKA: Recruitment, Reprogramming and Efferocytosis. Cells 2020, 9, 128. [Google Scholar] [CrossRef]
- Sullivan, G.W.; Rieger, J.M.; Scheld, W.M.; Macdonald, T.L.; Linden, J. Cyclic AMP-dependent inhibition of human neutrophil oxidative activity by substituted 2-propynylcyclohexyl adenosine A(2A) receptor agonists. Br. J. Pharmacol. 2001, 132, 1017–1026. [Google Scholar] [CrossRef]
- Schafer, P.H.; Truzzi, F.; Parton, A.; Wu, L.; Kosek, J.; Zhang, L.H.; Horan, G.; Saltari, A.; Quadri, M.; Lotti, R.; et al. Phosphodiesterase 4 in inflammatory diseases: Effects of apremilast in psoriatic blood and in dermal myofibroblasts through the PDE4/CD271 complex. Cell. Signal. 2016, 28, 753–763. [Google Scholar] [CrossRef]
- Ariga, M.; Neitzert, B.; Nakae, S.; Mottin, G.; Bertrand, C.; Pruniaux, M.P.; Jin, S.L.; Conti, M. Nonredundant function of phosphodiesterases 4D and 4B in neutrophil recruitment to the site of inflammation. J. Immunol. 2004, 173, 7531–7538. [Google Scholar] [CrossRef]
- Jin, S.L.; Goya, S.; Nakae, S.; Wang, D.; Bruss, M.; Hou, C.; Umetsu, D.; Conti, M. Phosphodiesterase 4B is essential for T(H)2-cell function and development of airway hyperresponsiveness in allergic asthma. J. Allergy Clin. Immunol. 2010, 126, 1252–1259.e12. [Google Scholar] [CrossRef]
- Peter, D.; Jin, S.L.; Conti, M.; Hatzelmann, A.; Zitt, C. Differential expression and function of phosphodiesterase 4 (PDE4) subtypes in human primary CD4+ T cells: Predominant role of PDE4D. J. Immunol. 2007, 178, 4820–4831. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.L.; Conti, M. Induction of the cyclic nucleotide phosphodiesterase PDE4B is essential for LPS-activated TNF-α responses. Proc. Natl. Acad. Sci. USA 2002, 99, 7628–7633. [Google Scholar] [CrossRef]
- Jin, S.L.; Lan, L.; Zoudilova, M.; Conti, M. Specific role of phosphodiesterase 4B in lipopolysaccharide-induced signaling in mouse macrophages. J. Immunol. 2005, 175, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Gobejishvili, L.; Avila, D.V.; Barker, D.F.; Ghare, S.; Henderson, D.; Brock, G.N.; Kirpich, I.A.; Joshi-Barve, S.; Mokshagundam, S.P.; McClain, C.J.; et al. S-adenosylmethionine decreases lipopolysaccharide-induced phosphodiesterase 4B2 and attenuates tumor necrosis factor expression via cAMP/protein kinase A pathway. J. Pharmacol. Exp. Ther. 2011, 337, 433–443. [Google Scholar] [CrossRef]
- Bao, F.; Fleming, J.C.; Golshani, R.; Pearse, D.D.; Kasabov, L.; Brown, A.; Weaver, L.C. A selective phosphodiesterase-4 inhibitor reduces leukocyte infiltration, oxidative processes, and tissue damage after spinal cord injury. J. Neurotrauma 2011, 28, 1035–1049. [Google Scholar] [CrossRef]
- Epstein, P.M.; Basole, C.; Brocke, S. The Role of PDE8 in T Cell Recruitment and Function in Inflammation. Front. Cell Dev. Biol. 2021, 9, 636778. [Google Scholar] [CrossRef]
- Kelkar, S.; Dong, Q.; Xiao, Y.; Joshi-Barve, S.; McClain, C.J.; Barve, S.S. Ethanol enhances activation-induced caspase-3 dependent cell death in T lymphocytes. Alcohol Clin. Exp. Res. 2002, 26, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Hote, P.T.; Sahoo, R.; Jani, T.S.; Ghare, S.S.; Chen, T.; Joshi-Barve, S.; McClain, C.J.; Barve, S.S. Ethanol inhibits methionine adenosyltransferase II activity and S-adenosylmethionine biosynthesis and enhances caspase-3-dependent cell death in T lymphocytes: Relevance to alcohol-induced immunosuppression. J. Nutr. Biochem. 2008, 19, 384–391. [Google Scholar] [CrossRef]
- Ghare, S.; Patil, M.; Hote, P.; Suttles, J.; McClain, C.; Barve, S.; Joshi-Barve, S. Ethanol inhibits lipid raft-mediated TCR signaling and IL-2 expression: Potential mechanism of alcohol-induced immune suppression. Alcohol Clin. Exp. Res. 2011, 35, 1435–1444. [Google Scholar] [CrossRef]
- Netherton, S.J.; Sutton, J.A.; Wilson, L.S.; Carter, R.L.; Maurice, D.H. Both protein kinase A and exchange protein activated by cAMP coordinate adhesion of human vascular endothelial cells. Circ. Res. 2007, 101, 768–776. [Google Scholar] [CrossRef]
- Houslay, M.D.; Baillie, G.S.; Maurice, D.H. cAMP-Specific phosphodiesterase-4 enzymes in the cardiovascular system: A molecular toolbox for generating compartmentalized cAMP signaling. Circ. Res. 2007, 100, 950–966. [Google Scholar] [CrossRef] [PubMed]
- Kreisel, W.; Lazaro, A.; Trebicka, J.; Grosse Perdekamp, M.; Schmitt-Graeff, A.; Deibert, P. Cyclic GMP in Liver Cirrhosis-Role in Pathophysiology of Portal Hypertension and Therapeutic Implications. Int. J. Mol. Sci. 2021, 22, 10372. [Google Scholar] [CrossRef] [PubMed]
- Brusilovskaya, K.; Konigshofer, P.; Lampach, D.; Szodl, A.; Supper, P.; Bauer, D.; Beer, A.; Stift, J.; Timelthaler, G.; Oberhuber, G.; et al. Soluble guanylyl cyclase stimulation and phosphodiesterase-5 inhibition improve portal hypertension and reduce liver fibrosis in bile duct-ligated rats. United Eur. Gastroenterol. J. 2020, 8, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- Knorr, A.; Hirth-Dietrich, C.; Alonso-Alija, C.; Harter, M.; Hahn, M.; Keim, Y.; Wunder, F.; Stasch, J.P. Nitric oxide-independent activation of soluble guanylate cyclase by BAY 60-2770 in experimental liver fibrosis. Arzneimittelforschung 2008, 58, 71–80. [Google Scholar] [CrossRef]
- Choi, S.M.; Shin, J.H.; Kim, J.M.; Lee, C.H.; Kang, K.K.; Ahn, B.O.; Yoo, M. Effect of udenafil on portal venous pressure and hepatic fibrosis in rats. A novel therapeutic option for portal hypertension. Arzneimittelforschung 2009, 59, 641–646. [Google Scholar]
- Siqueira, C.; de Moura, M.C.; Pedro, A.J.; Rocha, P. Elevated nitric oxide and 3′,5′ cyclic guanosine monophosphate levels in patients with alcoholic cirrhosis. World J. Gastroenterol. 2008, 14, 236–242. [Google Scholar] [CrossRef]
- Kirstetter, P.; Moreau, R.; Vachiery, F.; Gadano, A.; Soupison, T.; Pilette, C.; Pussard, E.; Cailmail, S.; Takahashi, H.; Lebrec, D. Plasma concentrations of cyclic 3′, 5′-guanosine monophosphate in patients with cirrhosis: Relationship with atrial natriuretic peptide and haemodynamics. J. Gastroenterol. Hepatol. 1997, 12, 233–236. [Google Scholar] [CrossRef]
- Montoliu, C.; Kosenko, E.; Del Olmo, J.A.; Serra, M.A.; Rodrigo, J.M.; Felipo, V. Correlation of nitric oxide and atrial natriuretic peptide changes with altered cGMP homeostasis in liver cirrhosis. Liver Int. 2005, 25, 787–795. [Google Scholar] [CrossRef]
- Wang, J.; Kazmi, M.M.; Huxley, V.H. Microvascular Sex- and Age- Dependent Phosphodiesterase Expression. Front. Aging 2021, 2, 719698. [Google Scholar] [CrossRef]
- Stangherlin, A.; Zaccolo, M. Local termination of 3′-5′-cyclic adenosine monophosphate signals: The role of A kinase anchoring protein-tethered phosphodiesterases. J. Cardiovasc. Pharmacol. 2011, 58, 345–353. [Google Scholar] [CrossRef]
- Collins, K.B.; Scott, J.D. Phosphorylation, compartmentalization, and cardiac function. IUBMB Life 2023, 75, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Pidoux, G.; Tasken, K. Specificity and spatial dynamics of protein kinase A signaling organized by A-kinase-anchoring proteins. J. Mol. Endocrinol. 2010, 44, 271–284. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, H.; Melick, C.H.; Jeong, M.H.; Curukovic, A.; Tiwary, S.; Lama-Sherpa, T.D.; Meng, D.; Servage, K.A.; James, N.G.; et al. AKAP13 couples GPCR signaling to mTORC1 inhibition. PLoS Genet. 2021, 17, e1009832. [Google Scholar] [CrossRef]
- Chen, H.; Shen, F.; Sherban, A.; Nocon, A.; Li, Y.; Wang, H.; Xu, M.J.; Rui, X.; Han, J.; Jiang, B.; et al. DEP domain-containing mTOR-interacting protein suppresses lipogenesis and ameliorates hepatic steatosis and acute-on-chronic liver injury in alcoholic liver disease. Hepatology 2018, 68, 496–514. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Zhong, W.; Hao, L.; Sun, X.; Zhou, Z. Activation of mTORC1 by Free Fatty Acids Suppresses LAMP2 and Autophagy Function via ER Stress in Alcohol-Related Liver Disease. Cells 2021, 10, 2730. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Choi, J.; Son, T.; Wee, H.J.; Bae, S.J.; Seo, J.H.; Park, J.H.; Ryu, S.H.; Lee, D.; Jang, M.K.; et al. Altered AKAP12 expression in portal fibroblasts and liver sinusoids mediates transition from hepatic fibrogenesis to fibrosis resolution. Exp. Mol. Med. 2018, 50, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Luo, Y.; Wang, S.; Li, Y.; Bao, M.; Shang, Y.; Chen, L.; Liu, W. AKAP12 ameliorates liver injury via targeting PI3K/AKT/PCSK6 pathway. Redox Biol. 2022, 53, 102328. [Google Scholar] [CrossRef] [PubMed]
- Ramani, K.; Mavila, N.; Abeynayake, A.; Tomasi, M.L.; Wang, J.; Matsuda, M.; Seki, E. Targeting A-kinase anchoring protein 12 phosphorylation in hepatic stellate cells regulates liver injury and fibrosis in mouse models. Elife 2022, 11, e78430. [Google Scholar] [CrossRef]
- Li, H. Physiologic and pathophysiologic roles of AKAP12. Sci. Prog. 2022, 105, 368504221109212. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Zhao, Y.X.; Li, X.F.; Huang, C.; Meng, X.M.; Li, J. beta-Arrestin 2 Promotes Hepatocyte Apoptosis by Inhibiting Akt Pathway in Alcoholic Liver Disease. Front. Pharmacol. 2018, 9, 1031. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pair-Fed | Alcohol-Fed | |
|---|---|---|
| ALT, U/L | 15 ± 5.8 | 348.3 ± 169.01 ** |
| AST, U/L | 53.5 ± 8.9 | 289 ± 190.4 * |
| MPO activity, Units/mg | 9.6 ± 1.97 | 32.4 ± 16.68 * |
| TNFα, pg/g | 4.3 ± 2.63 | 10 ± 3.8 * |
| IP-10, pg/g | 156.8 ± 25.69 | 367.5 ± 117.97 ** |
| KC-GRO, pg/g | 155.3 ± 27.18 | 362.7 ± 106.15 ** |
| GM-CSF, pg/g | 0.15 ± 0.05 | 0.3 ± 0.08 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montoya-Durango, D.; Walter, M.N.; Rodriguez, W.; Wang, Y.; Chariker, J.H.; Rouchka, E.C.; Maldonado, C.; Barve, S.; McClain, C.J.; Gobejishvili, L. Dysregulated Cyclic Nucleotide Metabolism in Alcohol-Associated Steatohepatitis: Implications for Novel Targeted Therapies. Biology 2023, 12, 1321. https://doi.org/10.3390/biology12101321
Montoya-Durango D, Walter MN, Rodriguez W, Wang Y, Chariker JH, Rouchka EC, Maldonado C, Barve S, McClain CJ, Gobejishvili L. Dysregulated Cyclic Nucleotide Metabolism in Alcohol-Associated Steatohepatitis: Implications for Novel Targeted Therapies. Biology. 2023; 12(10):1321. https://doi.org/10.3390/biology12101321
Chicago/Turabian StyleMontoya-Durango, Diego, Mary Nancy Walter, Walter Rodriguez, Yali Wang, Julia H. Chariker, Eric C. Rouchka, Claudio Maldonado, Shirish Barve, Craig J. McClain, and Leila Gobejishvili. 2023. "Dysregulated Cyclic Nucleotide Metabolism in Alcohol-Associated Steatohepatitis: Implications for Novel Targeted Therapies" Biology 12, no. 10: 1321. https://doi.org/10.3390/biology12101321