
The Capacity of Magnesium to Induce Osteoclast Differentiation Is Greatly Enhanced by the Presence of Zoledronate

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.1.1. Osteoclast Differentiation
2.1.2. Pharmacological Treatments
2.1.3. Cell Counts
2.1.4. Morphological Analysis
2.2. Flow Cytometry
2.2.1. Cell Cycle and Apoptosis Analysis
2.2.2. Immune Phenotype Analysis
2.3. Molecular Analysis
2.3.1. RNA Extraction
2.3.2. Reverse Transcription
2.3.3. QRT-PCR Analysis
Analysis of Cell Cycle-Related Genes
Analysis of Differentiation-Related Genes
2.4. Statistical Analysis
3. Results
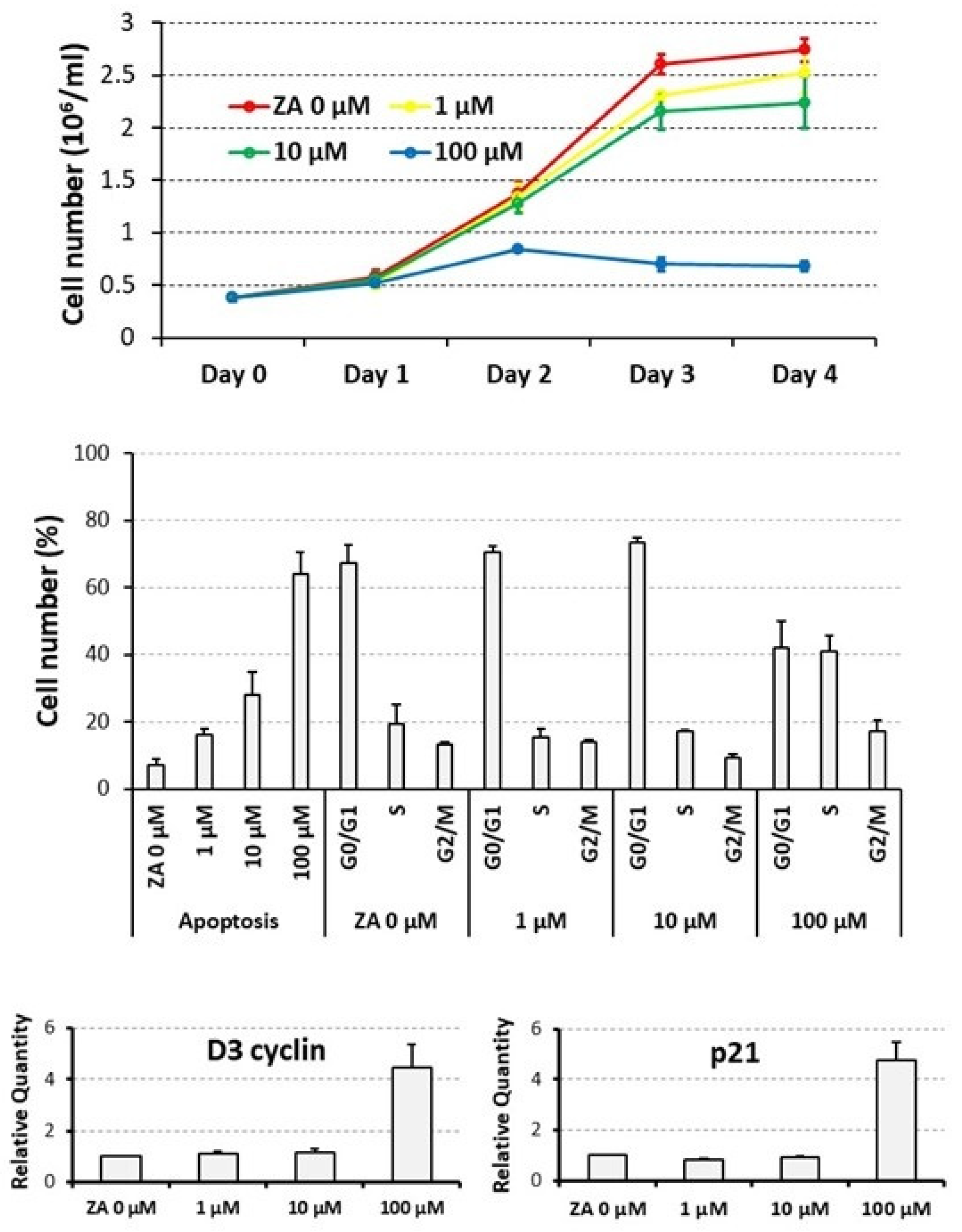
3.1. Proliferative and Apoptotic Effects Determined on Cycling U937 Cells by Scalar Concentrations of Zoledronate
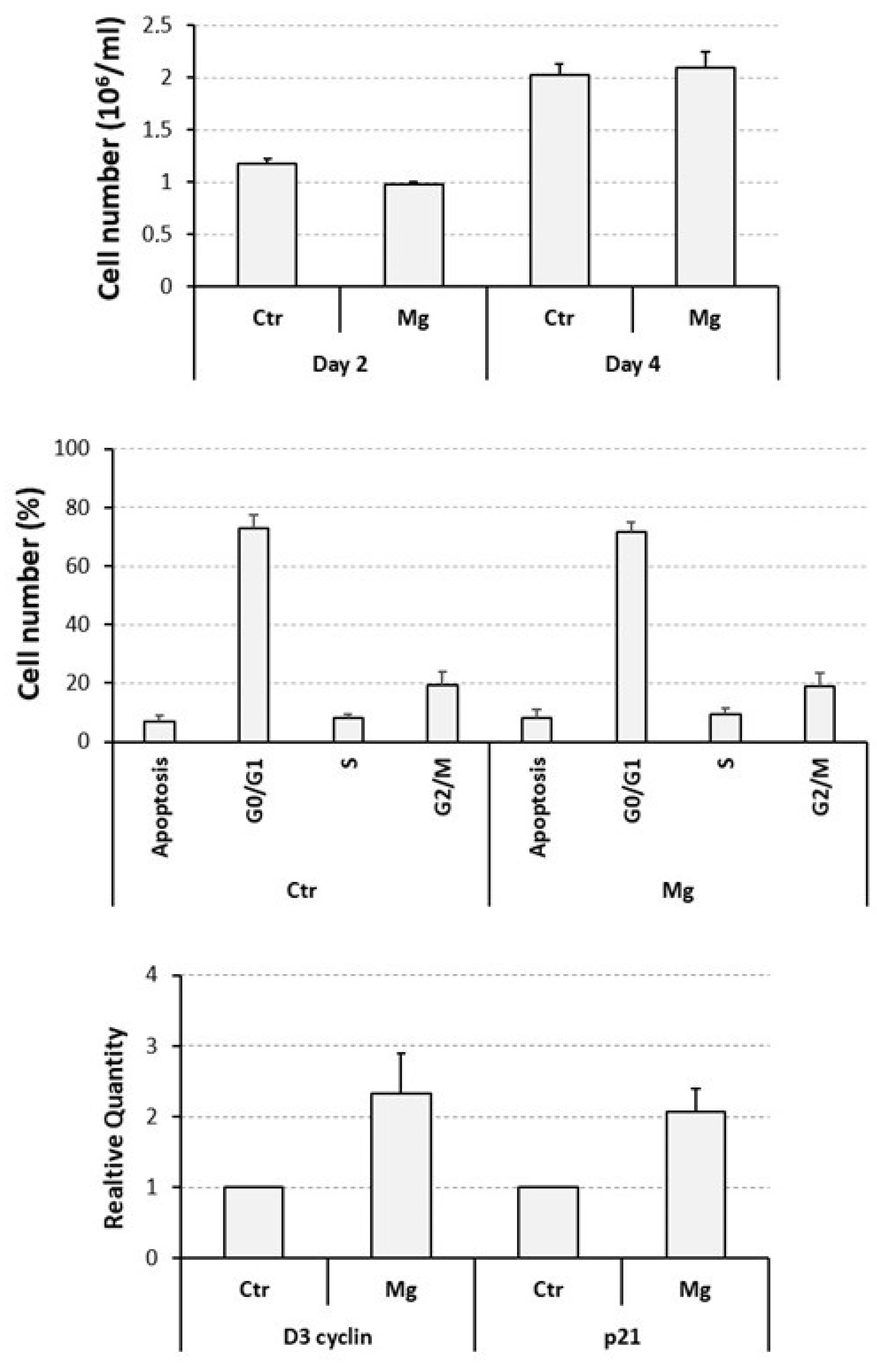
3.2. Proliferative and Apoptotic Effects Determined on Cycling U937 Cells by a Supra-Physiological Concentration of MgCl2
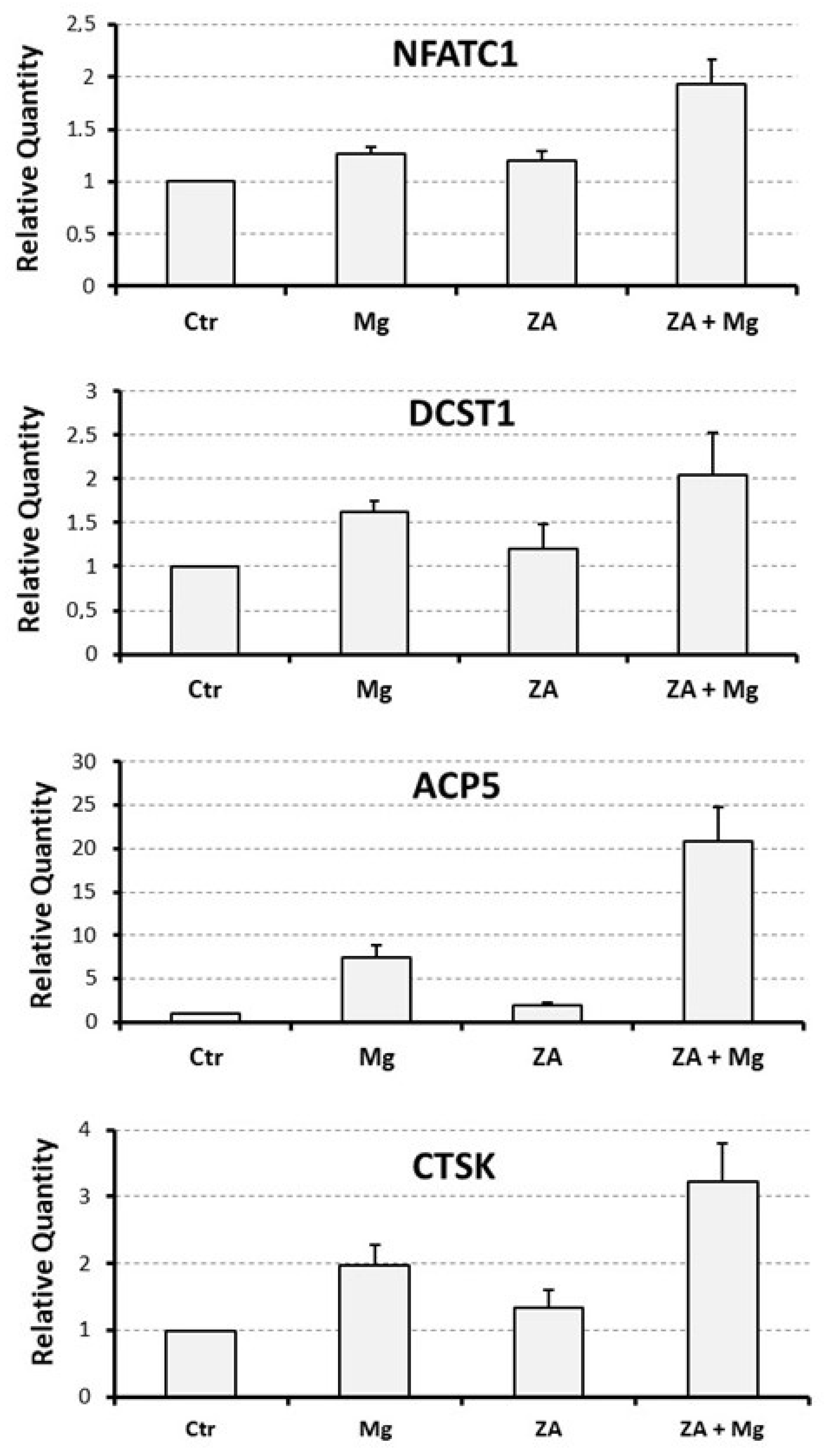
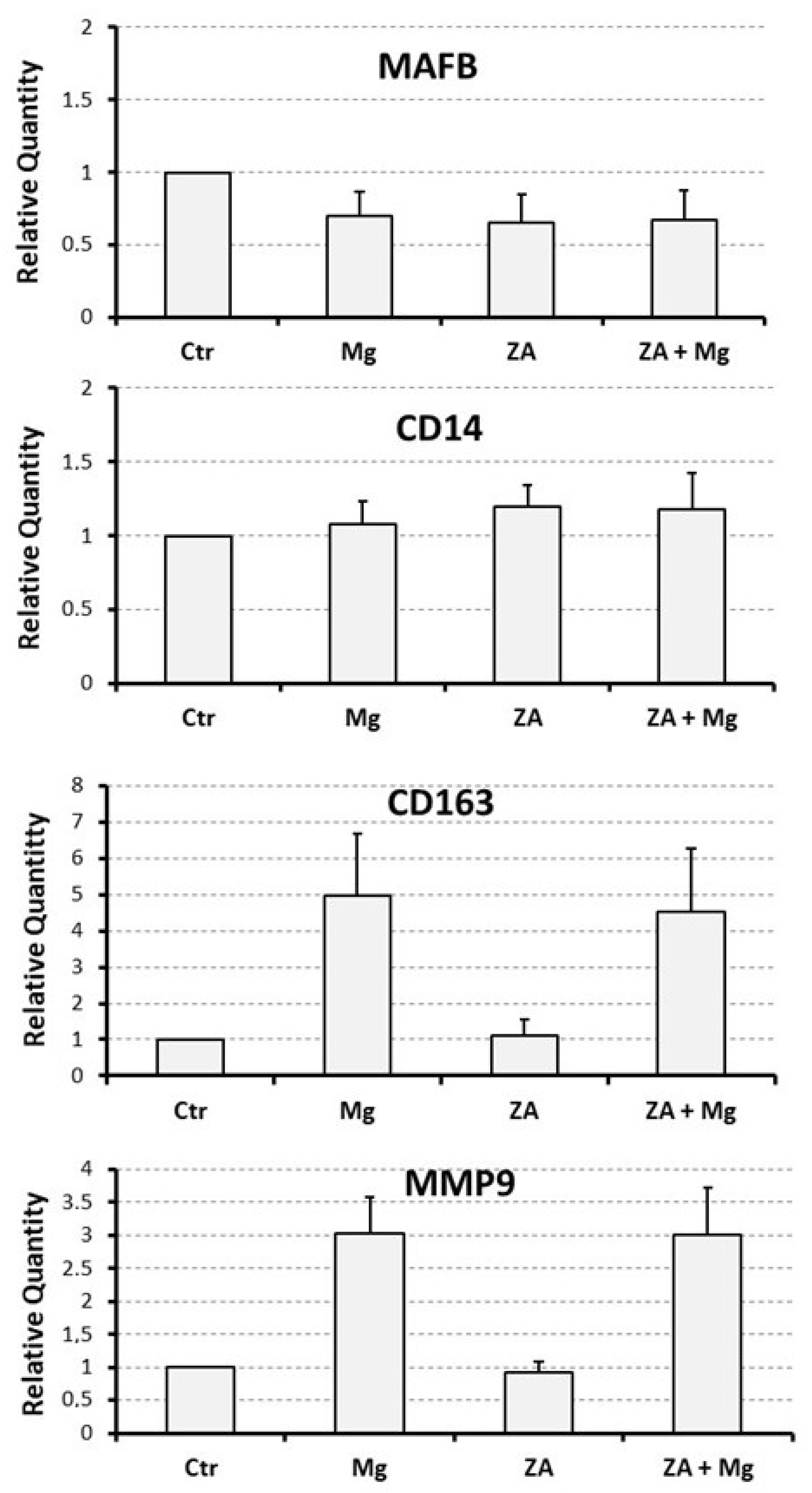
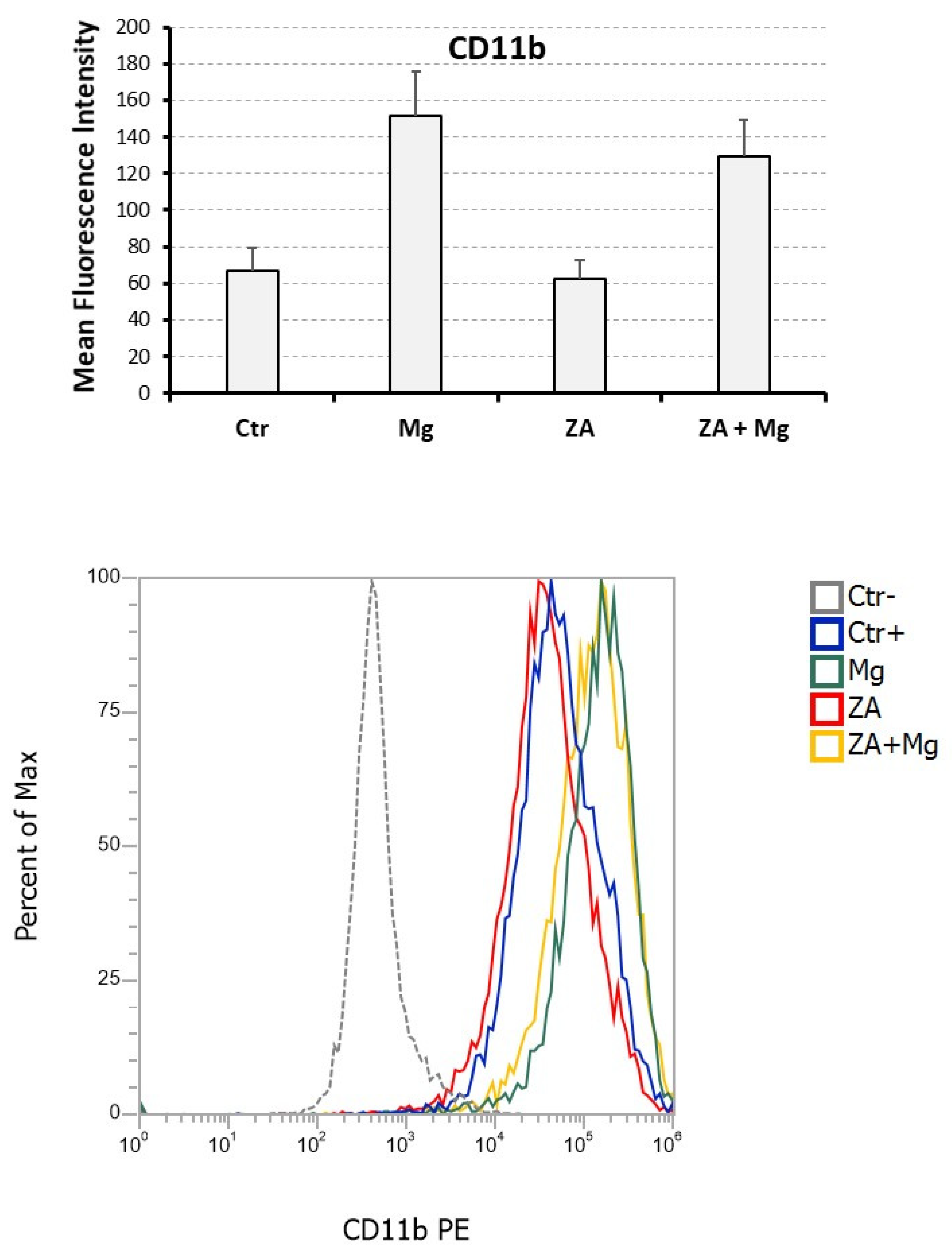
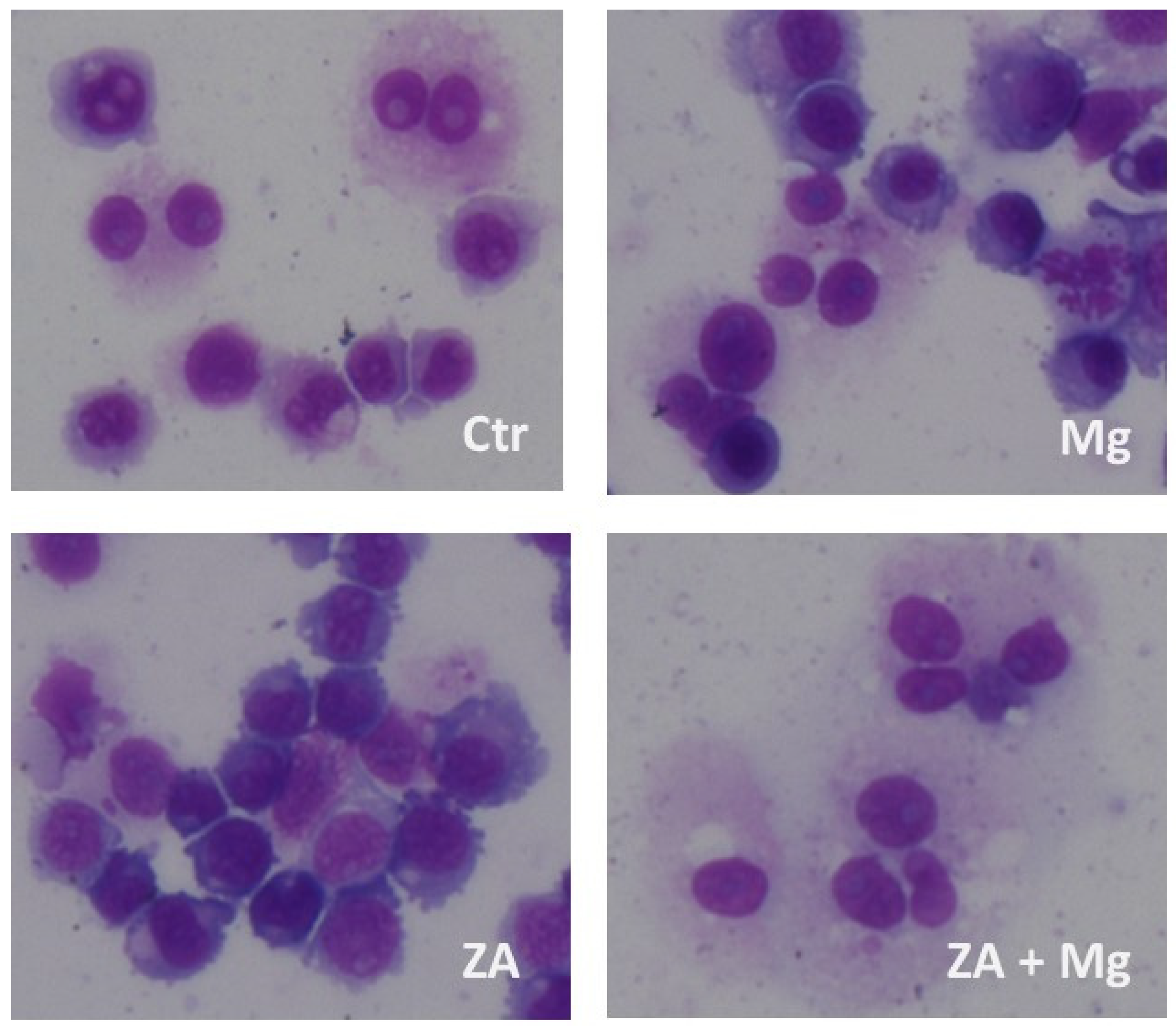
3.3. Capacity of MgCl2 and ZA to Modulate the Osteoclast Differentiation of U937 Cells Induced by Stimulation with Phorbol Esters and Vitamin D3

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyzed Marker | Ctr | Mg | ZA | ZA + Mg | Anova, p Value |
|---|---|---|---|---|---|
| NFATC1 | 1 | 1.3 ± 0.1 | 1.2 ± 0.1 | 1.9 ± 0.2 | 0.005 |
| DCST1 | 1 | 1.6 ± 0.1 | 1.2 ± 0.3 | 2.1 ± 0.5 | 0.3 |
| ACP5 | 1 | 7.5 ± 1.4 | 2.0 ± 0.3 | 20.9 ± 4.0 | 0.002 |
| CTSK | 1 | 2.0 ± 0.3 | 1.3 ± 0.3 | 3.2 ± 0.6 | 0.01 |
| MMP9 | 1 | 3.0 ± 0.6 | 0.9 ± 0.2 | 3.0 ± 0.7 | 0.01 |
| MAFB | 1 | 0.7 ± 0.2 | 0.7 ± 0.2 | 0.7 ± 0.2 | 0.4 |
| CD14 | 1 | 1.1 ± 0.2 | 1.2 ± 0.2 | 1.1 ± 0.3 | 0.8 |
| CD163 | 1 | 5.0 ± 1.7 | 1.1 ± 0.4 | 4.5 ± 1.7 | 0.01 |
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cremers, S.; Drake, M.T.; Ebetino, F.; Bilezikian, J.P.; Graham, R.; Russell, G. Pharmacology of Bisphosphonates. Br. J. Clin. Pharmacol. 2019, 85, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhan, Y.; Yan, L.; Hao, D. How Zoledronic Acid Improves Osteoporosis by Acting on Osteoclasts. Front. Pharmacol. 2022, 13, 961941. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fang, D.; Xu, J.; Luo, R. Various Pathways of Zoledronic Acid against Osteoclasts and Bone Cancer Metastasis: A Brief Review. BMC Cancer 2020, 20, 1059. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Zhao, J. Functionalized Selenium Nanotherapeutics Synergizes With Zoledronic Acid to Suppress Prostate Cancer Cell Growth Through Induction of Mitochondria-Mediated Apoptosis and Cell Cycle S Phase Arrest. Front. Oncol. 2021, 11, 685784. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Altman, R.B.; Klein, T.E. Bisphosphonates Pathway. Pharmacogenet 2011, 21, 50–53. [Google Scholar] [CrossRef]
- Kimmel, D.B. Mechanism of Action, Pharmacokinetic and Pharmacodynamic Profile, and Clinical Applications of Nitrogen-Containing Bisphosphonates. J. Dent. Res. 2007, 86, 1022–1033. [Google Scholar] [CrossRef]
- Moriyama, Y.; Nomura, M. Clodronate: A Vesicular ATP Release Blocker. Trends Pharmacol. Sci. 2018, 39, 13–23. [Google Scholar] [CrossRef]
- Kenkre, J.S.; Bassett, J. The Bone Remodelling Cycle. Ann. Clin. Biochem. 2018, 55, 308–327. [Google Scholar] [CrossRef]
- Marx, R.E. Pamidronate (Aredia) and Zoledronate (Zometa) Induced Avascular Necrosis of the Jaws: A Growing Epidemic. J. Oral Maxillofac. Surg. 2003, 61, 1115–1117. [Google Scholar] [CrossRef]
- Khan, A.A.; Morrison, A.; Hanley, D.A.; Felsenberg, D.; McCauley, L.K.; O’Ryan, F.; Reid, I.R.; Ruggiero, S.L.; Taguchi, A.; Tetradis, S.; et al. Diagnosis and Management of Osteonecrosis of the Jaw: A Systematic Review and International Consensus. J. Bone Miner. Res. 2015, 30, 3–23. [Google Scholar] [CrossRef]
- Ruggiero, S.L.; Dodson, T.B.; Aghaloo, T.; Carlson, E.R.; Ward, B.B.; Kademani, D. American Association of Oral and Maxillofacial Surgeons’ Position Paper on Medication-Related Osteonecrosis of the Jaws—2022 Update. J. Oral Maxillofac. Surg. 2022, 80, 920–943. [Google Scholar] [CrossRef]
- Kawahara, M.; Kuroshima, S.; Sawase, T. Clinical Considerations for Medication-Related Osteonecrosis of the Jaw: A Comprehensive Literature Review. Int. J. Implant Dent. 2021, 7, 47. [Google Scholar] [CrossRef]
- Yamazaki, T.; Yamori, M.; Ishizaki, T.; Asai, K.; Goto, K.; Takahashi, K.; Nakayama, T.; Bessho, K. Increased Incidence of Osteonecrosis of the Jaw after Tooth Extraction in Patients Treated with Bisphosphonates: A Cohort Study. Int. J. Oral Maxillofac. Surg. 2012, 41, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Otto, S.; Tröltzsch, M.; Jambrovic, V.; Panya, S.; Probst, F.; Ristow, O.; Ehrenfeld, M.; Pautke, C. Tooth Extraction in Patients Receiving Oral or Intravenous Bisphosphonate Administration: A Trigger for BRONJ Development? J. Craniomaxillofac. Surg. 2015, 43, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Aghaloo, T.; Hazboun, R.; Tetradis, S. Pathophysiology of Osteonecrosis of the Jaws. Oral Maxillofac. Surg. Clin. N. Am. 2015, 27, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Hakam, A.E.; McCauley, L.K. Current Understanding of the Pathophysiology of Osteonecrosis of the Jaw. Curr. Osteoporos. Rep. 2018, 16, 584–595. [Google Scholar] [CrossRef]
- Hoefert, S.; Schmitz, I.; Weichert, F.; Gaspar, M.; Eufinger, H. Macrophages and Bisphosphonate-Related Osteonecrosis of the Jaw (BRONJ): Evidence of Local Immunosuppression of Macrophages in Contrast to Other Infectious Jaw Diseases. Clin. Oral Investig. 2015, 19, 497–508. [Google Scholar] [CrossRef]
- Pazianas, M.; Miller, P.; Blumentals, W.A.; Bernal, M.; Kothawala, P. A review of the literature on osteonecrosis of the jaw in patients with osteoporosis treated with oral bisphosphonates: Prevalence, risk factors, and clinical characteristics. Clin. Ther. 2007, 29, 1548–1558. [Google Scholar] [CrossRef]
- Khan, A.; Morrison, A.; Cheung, A.; Hashem, W.; Compston, J. Osteonecrosis of the Jaw (ONJ): Diagnosis and Management in 2015. Osteoporos. Int. 2016, 27, 853–859. [Google Scholar] [CrossRef]
- Rupel, K.; Ottaviani, G.; Gobbo, M.; Contardo, L.; Tirelli, G.; Vescovi, P.; Di Lenarda, R.; Biasotto, M. A Systematic Review of Therapeutical Approaches in Bisphosphonates-Related Osteonecrosis of the Jaw (BRONJ). Oral Oncol. 2014, 50, 1049–1057. [Google Scholar] [CrossRef]
- Fliefel, R.; Tröltzsch, M.; Kühnisch, J.; Ehrenfeld, M.; Otto, S. Treatment Strategies and Outcomes of Bisphosphonate-Related Osteonecrosis of the Jaw (BRONJ) with Characterization of Patients: A Systematic Review. Int. J. Oral Maxillofac. Surg. 2015, 44, 568–585. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, S.; Soutome, S.; Yanamoto, S.; Fujita, S.; Hasegawa, T.; Komori, T.; Kojima, Y.; Miyamoto, H.; Shibuya, Y.; Ueda, N.; et al. Evaluation of the Treatment Strategies for Medication-Related Osteonecrosis of the Jaws (MRONJ) and the Factors Affecting Treatment Outcome: A Multicenter Retrospective Study with Propensity Score Matching Analysis. J. Bone Miner. Res. 2017, 32, 2022–2029. [Google Scholar] [CrossRef] [PubMed]
- Anesi, A.; Generali, L.; Sandoni, L.; Pozzi, S.; Grande, A. From Osteoclast Differentiation to Osteonecrosis of the Jaw: Molecular and Clinical Insights. Int. J. Mol. Sci. 2019, 20, 4925. [Google Scholar] [CrossRef]
- Takahashi, N.; Udagawa, N.; Suda, T. Vitamin D Endocrine System and Osteoclasts. BoneKEy Rep. 2014, 3, 495. [Google Scholar] [CrossRef] [PubMed]
- Amoui, M.; Suhr, S.-M.; Baylink, D.J.; Lau, K.-H.W. An Osteoclastic Protein-Tyrosine Phosphatase May Play a Role in Differentiation and Activity of Human Monocytic U-937 Cell-Derived, Osteoclast-like Cells. Am. J. Physiol. Cell. Physiol. 2004, 287, C874–C884. [Google Scholar] [CrossRef]
- Mammoli, F.; Castiglioni, S.; Parenti, S.; Cappadone, C.; Farruggia, G.; Iotti, S.; Davalli, P.; Maier, J.; Grande, A.; Frassineti, C. Magnesium Is a Key Regulator of the Balance between Osteoclast and Osteoblast Differentiation in the Presence of Vitamin D3. Int. J. Mol. Sci. 2019, 20, 385. [Google Scholar] [CrossRef]
- Gong, X.; Yu, W.; Zhao, H.; Su, J.; Sheng, Q. Skeletal Site-Specific Effects of Zoledronate on in Vivo Bone Remodeling and in Vitro BMSCs Osteogenic Activity. Sci. Rep. 2017, 7, 36129. [Google Scholar] [CrossRef]
- Selmi, T.; Alecci, C.; dell’ Aquila, M.; Montorsi, L.; Martello, A.; Guizzetti, F.; Volpi, N.; Parenti, S.; Ferrari, S.; Salomoni, P.; et al. ZFP36 stabilizes RIP1 via degradation of XIAP and cIAP2 thereby promoting ripoptosome assembly. BMC Cancer 2015, 6, 357. [Google Scholar] [CrossRef]
- Manzotti, G.; Parenti, S.; Ferrari-Amorotti, G.; Soliera, A.R.; Cattelani, S.; Montanari, M.; Cavalli, D.; Ertel, A.; Grande, A.; Calabretta, B. Monocyte-macrophage differentiation of acute myeloid leukemia cell lines by small molecules identified through interrogation of the Connectivity Map database. Cell Cycle 2015, 14, 2578–2589. [Google Scholar] [CrossRef]
- Parenti, S.; Sandoni, L.; Montanari, M.; Zanocco-Marani, T.; Anesi, A.; Iotti, S.; Manfredini, R.; Frassineti, C.; Davalli, P.; Grande, A. Magnesium Favors the Capacity of Vitamin D3 to Induce the Monocyte Differentiation of U937 Cells. Magnes. Res. 2021, 34, 114–129. [Google Scholar]
- Li, P.; Zhao, Z.; Wang, L.; Jin, X.; Shen, Y.; Nan, C.; Liu, H. Minimally Effective Concentration of Zoledronic Acid to Suppress Osteoclasts In Vitro. Exp. Ther. Med. 2018, 15, 5330–5336. [Google Scholar] [PubMed]
- Boyle, W.J.; Scott Simonet, W.; Lacey, D.L. Osteoclast Differentiation and Activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Takayanagi, H. The Molecular Understanding of Osteoclast Differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 Expression Determines Its Essential Role in Bone Homeostasis. Exp. Biol. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T. Regulators of Osteoclast Differentiation and Cell-Cell Fusion. Keio J. Med. 2011, 60, 101–105. [Google Scholar] [CrossRef]
- Yang, G.; Chen, X.; Yan, Z.; Zhu, Q.; Yang, C. CD11b Promotes the Differentiation of Osteoclasts Induced by RANKL through the Spleen Tyrosine Kinase Signalling Pathway. J. Cell Mol. Med. 2017, 21, 3445–3452. [Google Scholar] [CrossRef]
- Palacios, C. The role of nutrients in bone health, from A to Z. Crit. Rev. Food Sci. Nutr. 2006, 46, 621–628. [Google Scholar] [CrossRef]
- Groenendijk, I.; van Delft, M.; Versloot, P.; van Loon, L.J.C.; de Groot, L.C.P.G.M. Impact of magnesium on bone health in older adults: A systematic review and meta-analysis. Bone 2022, 154, 116233. [Google Scholar] [CrossRef]
- Belluci, M.M.; Schoenmaker, T.; Rossa-Junior, C.; Orrico, S.R.; de Vries, T.J.; Everts, V. Magnesium deficiency results in an increased formation of osteoclasts. J. Nutr. Biochem. 2013, 24, 1488–1498. [Google Scholar] [CrossRef]
- Belluci, M.M.; de Molon, R.S.; Rossa, C., Jr.; Tetradis, S.; Giro, G.; Cerri, P.S.; Marcantonio, E., Jr.; Orrico, S.R.P. Severe magnesium deficiency compromises systemic bone mineral density and aggravates inflammatory bone resorption. J. Nutr. Biochem. 2020, 77, 108301. [Google Scholar] [CrossRef]
- Wu, L.; Feyerabend, F.; Schilling, A.F.; Willumeit-Römer, R.; Luthringer, B.J.C. Effects of Extracellular Magnesium Extract on the Proliferation and Differentiation of Human Osteoblasts and Osteoclasts in Coculture. Acta Biomater. 2015, 27, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Schacht, E.; Dukasù, L.; Richy, F. Combined therapies in osteoporosis: Bisphosphonates and vitamin D-hormone analogs. J. Musculoskelet. Neuronal. Interact. 2007, 7, 174–184. [Google Scholar] [PubMed]
- Shapiro, C.L. Bone-modifying Agents (BMAs) in Breast Cancer. Clin. Breast Cancer 2021, 21, e618–e630. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Li, X.; Xia, Y.; Yu, Z.; Cai, N.; Malwal, S.R.; Han, X.; Oldfield, E.; Zhang, Y. Farnesyl Pyrophosphate Synthase as a Target for Drug Development: Discovery of Natural-Product-Derived Inhibitors and Their Activity in Pancreatic Cancer Cells. J. Med. Chem. 2019, 62, 10867–10896. [Google Scholar] [CrossRef]
- Pelekanou, V.; Villarroel-Espindola, F.; Schalper, K.A.; Pusztai, L.; Rimm, D.L. CD68, CD163, and Matrix Metalloproteinase 9 (MMP-9) Co-Localization in Breast Tumor Microenvironment Predicts Survival Differently in ER-Positive and -Negative Cancers. Breast Cancer Res. 2018, 20, 154. [Google Scholar] [CrossRef]
- He, D.; Kou, X.; Luo, Q.; Yang, R.; Liu, D.; Wang, X.; Song, Y.; Cao, H.; Zeng, M.; Gan, Y.; et al. Enhanced M1/M2 Macrophage Ratio Promotes Orthodontic Root Resorption. J. Dent. Res. 2015, 94, 129–139. [Google Scholar] [CrossRef]
- Fukui, S.; Iwamoto, N.; Takatani, A.; Igawa, T.; Shimizu, T.; Umeda, M.; Nishino, A.; Horai, Y.; Hirai, Y.; Koga, T.; et al. M1 and M2 Monocytes in Rheumatoid Arthritis: A Contribution of Imbalance of M1/M2 Monocytes to Osteoclastogenesis. Front. Immunol. 2017, 8, 1958. [Google Scholar] [CrossRef]
- Rai, V.; Radwan, M.M.; Agrawal, D.K. IL-33, IL-37, and Vitamin D Interaction Mediate Immunomodulation of Inflammation in Degenerating Cartilage. Antibodies 2021, 10, 41. [Google Scholar] [CrossRef]
- Li, Z.H.; Si, Y.; Xu, G.; Chen, X.M.; Xiong, H.; Lai, L.; Zheng, Y.Q.; Zhang, Z.G. High-dose PMA with RANKL and MCSF induces THP-1 cell differentiation into human functional osteoclasts in vitro. Mol. Med. Rep. 2017, 16, 8380–8384. [Google Scholar] [CrossRef]
- Abdallah, D.; Jourdain, M.L.; Braux, J.; Guillaume, C.; Gangloff, S.C.; Jacquot, J.; Velard, F. An Optimized Method to Generate Human Active Osteoclasts from Peripheral Blood Monocytes. Front. Immunol. 2018, 9, 632. [Google Scholar] [CrossRef]
- Nagaoka, Y.; Kajiya, H.; Ozeki, S.; Ikebe, T.; Okabe, K. Mevalonates restore zoledronic acid-induced osteoclastogenesis inhibition. J. Dent. Res. 2015, 94, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, W.; Xu, R.; Shen, X.; Fu, Y.; Cheng, J.; Liu, L.; Jiang, H. Geranylgeraniol Restores Zoledronic Acid-Induced Efferocytosis Inhibition in Bisphosphonate-Related Osteonecrosis of the Jaw. Front. Cell Dev. Biol. 2021, 9, 770899. [Google Scholar] [CrossRef] [PubMed]
- Koneski, F.; Popovic-Monevska, D.; Gjorgoski, I.; Krajoska, J.; Popovska, M.; Muratovska, I.; Velickovski, B.; Petrushevska, G.; Popovski, V. In vivo effects of geranylgeraniol on the development of bisphosphonate-related osteonecrosis of the jaws. J. Craniomaxillofac. Surg. 2018, 46, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Draenert, G.F.; Huetzen, D.O.; Kämmerer, P.W.; Palarie, V.; Nacu, V.; Wagner, W. Dexrazoxane shows cytoprotective effects in zoledronic acid-treated human cells in vitro and in the rabbit tibia model in vivo. J. Craniomaxillofac. Surg. 2012, 40, e369–e374. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricchiuto, S.; Palumbo, R.; Lami, F.; Gavioli, F.; Caselli, L.; Montanari, M.; Zappavigna, V.; Anesi, A.; Zanocco-Marani, T.; Grande, A. The Capacity of Magnesium to Induce Osteoclast Differentiation Is Greatly Enhanced by the Presence of Zoledronate. Biology 2023, 12, 1297. https://doi.org/10.3390/biology12101297
Ricchiuto S, Palumbo R, Lami F, Gavioli F, Caselli L, Montanari M, Zappavigna V, Anesi A, Zanocco-Marani T, Grande A. The Capacity of Magnesium to Induce Osteoclast Differentiation Is Greatly Enhanced by the Presence of Zoledronate. Biology. 2023; 12(10):1297. https://doi.org/10.3390/biology12101297
Chicago/Turabian StyleRicchiuto, Silvia, Rossella Palumbo, Francesca Lami, Francesca Gavioli, Lorenzo Caselli, Monica Montanari, Vincenzo Zappavigna, Alexandre Anesi, Tommaso Zanocco-Marani, and Alexis Grande. 2023. "The Capacity of Magnesium to Induce Osteoclast Differentiation Is Greatly Enhanced by the Presence of Zoledronate" Biology 12, no. 10: 1297. https://doi.org/10.3390/biology12101297