
Multiomics Approach Captures Hepatic Metabolic Network Altered by Chronic Ethanol Administration
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
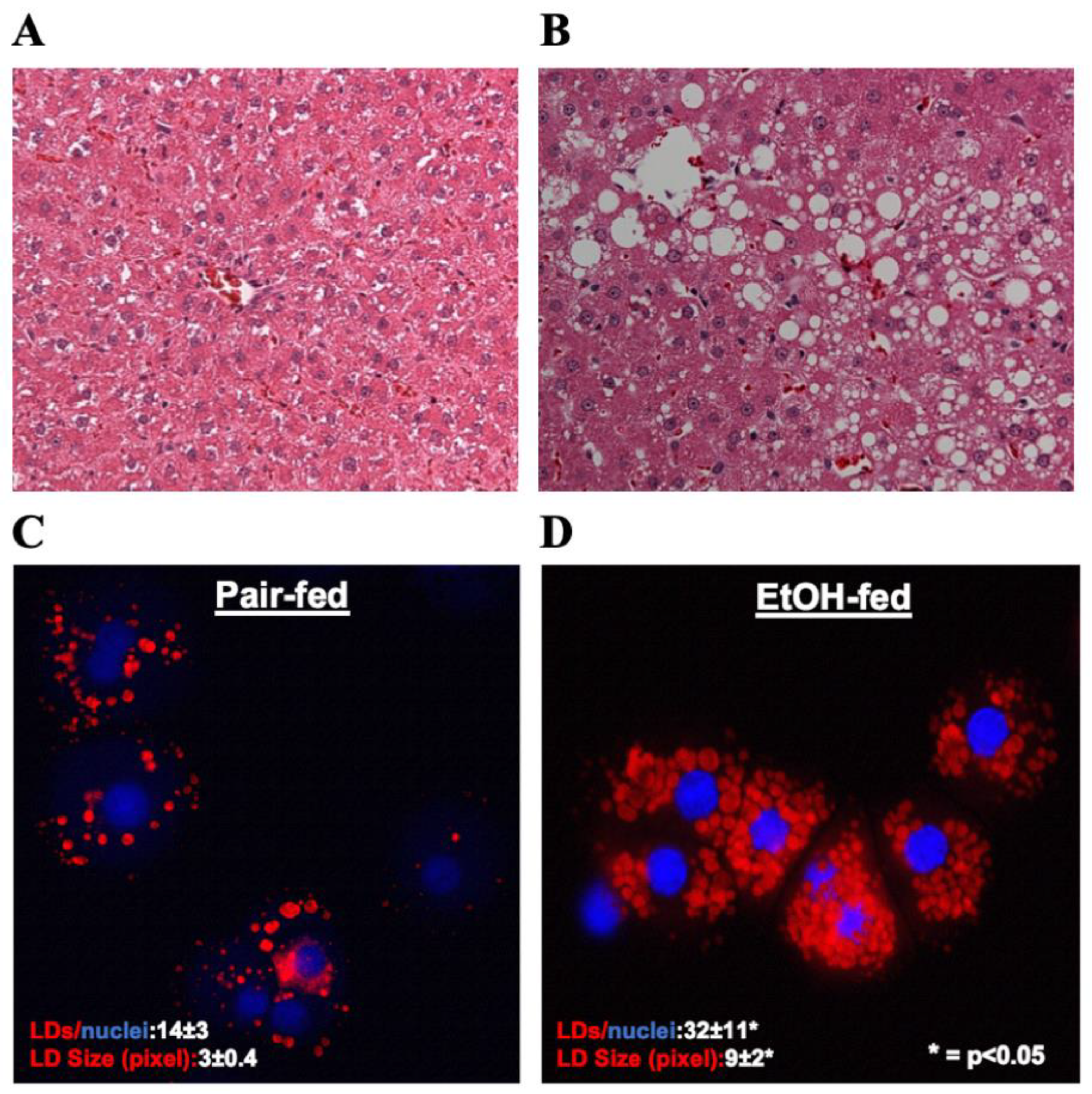
3.1. Ethanol Exposure Induced LD Accumulation in Hepatocytes
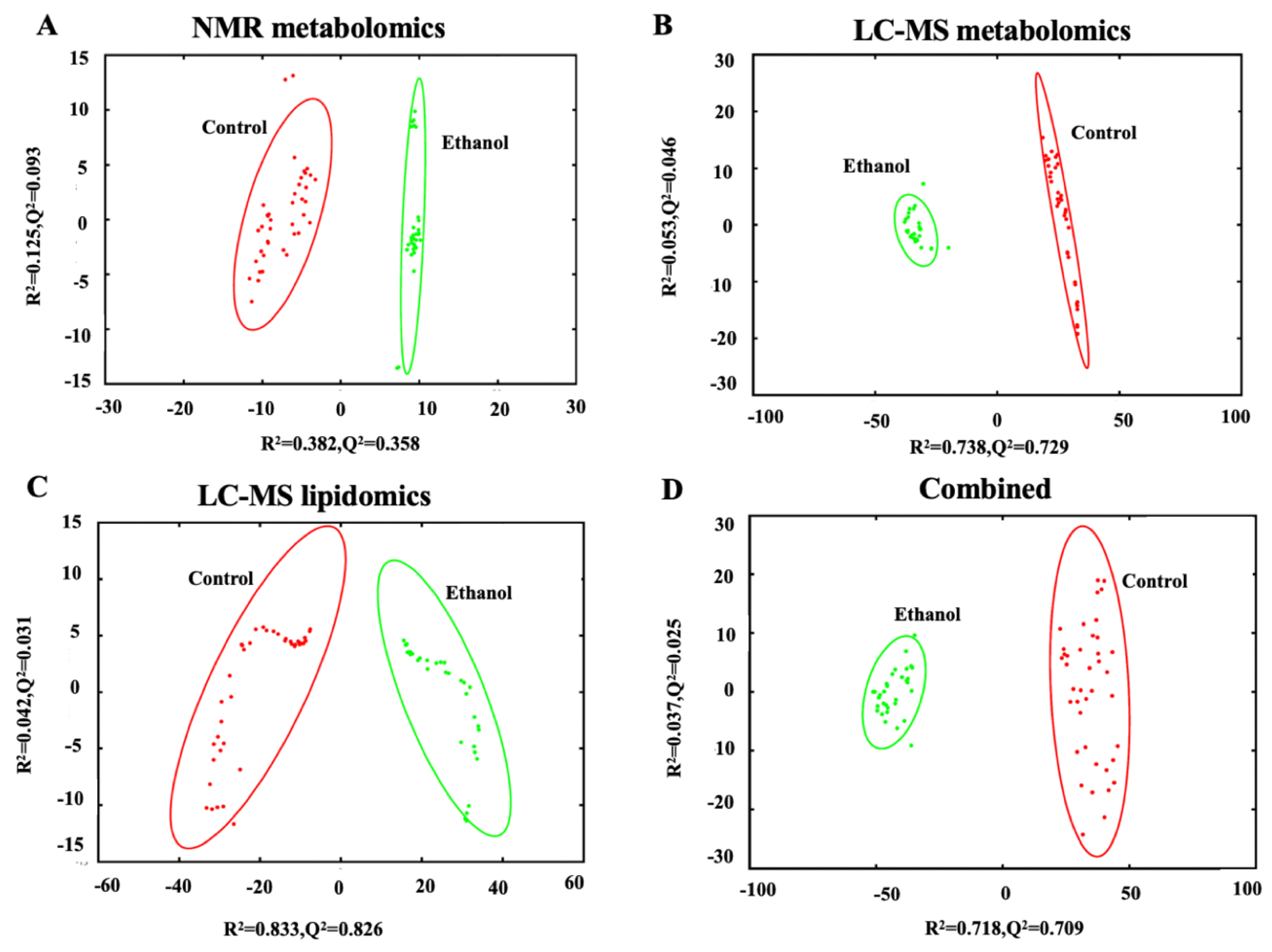
3.2. Ethanol Exposure Induces a Distinct Alteration in the Metabolome and Lipidome of Hepatocytes
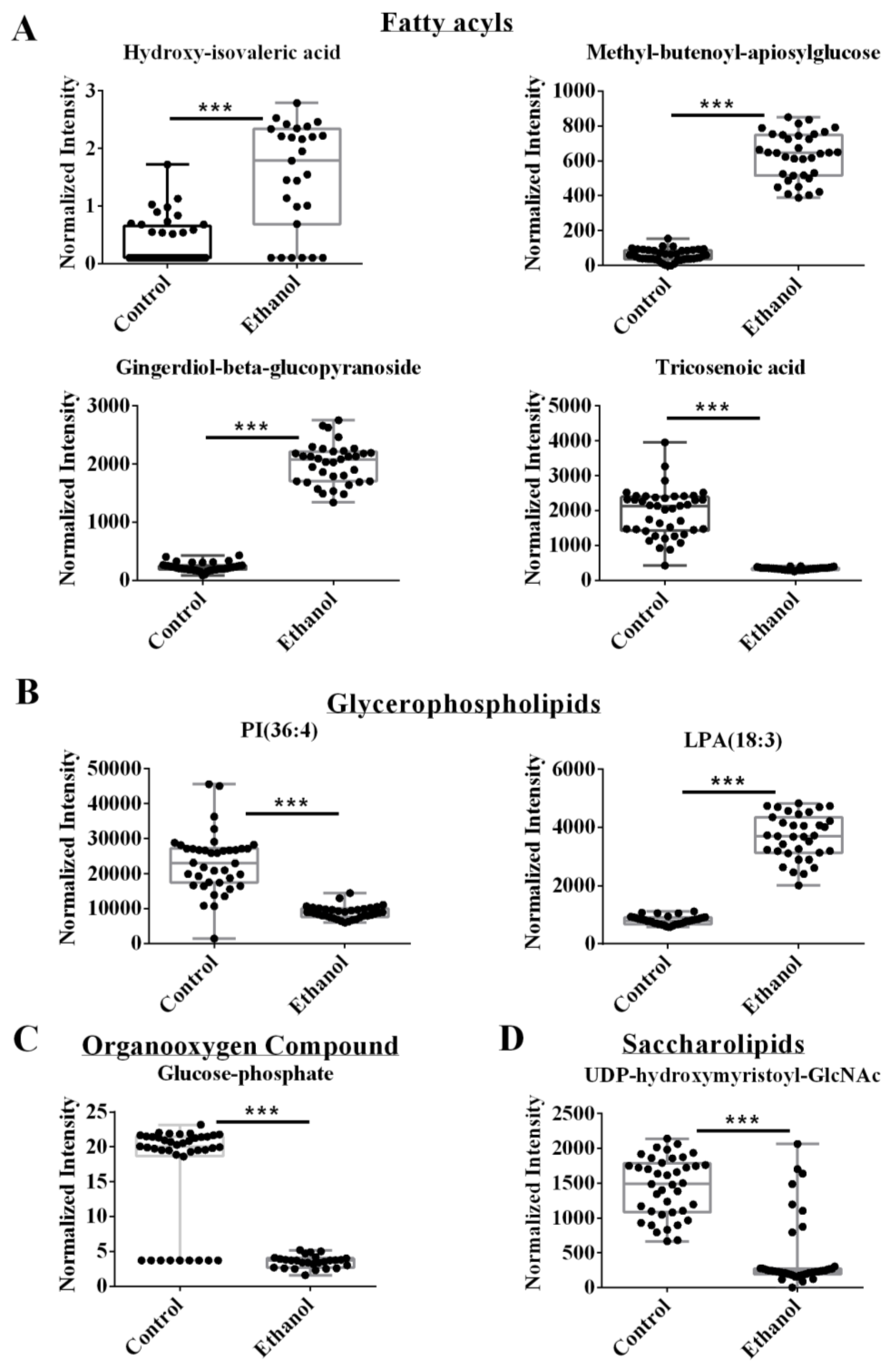
3.3. The Integrated Omics Dataset Identified Fatty Acyls and Glycerophospholipids as Being Altered by Ethanol
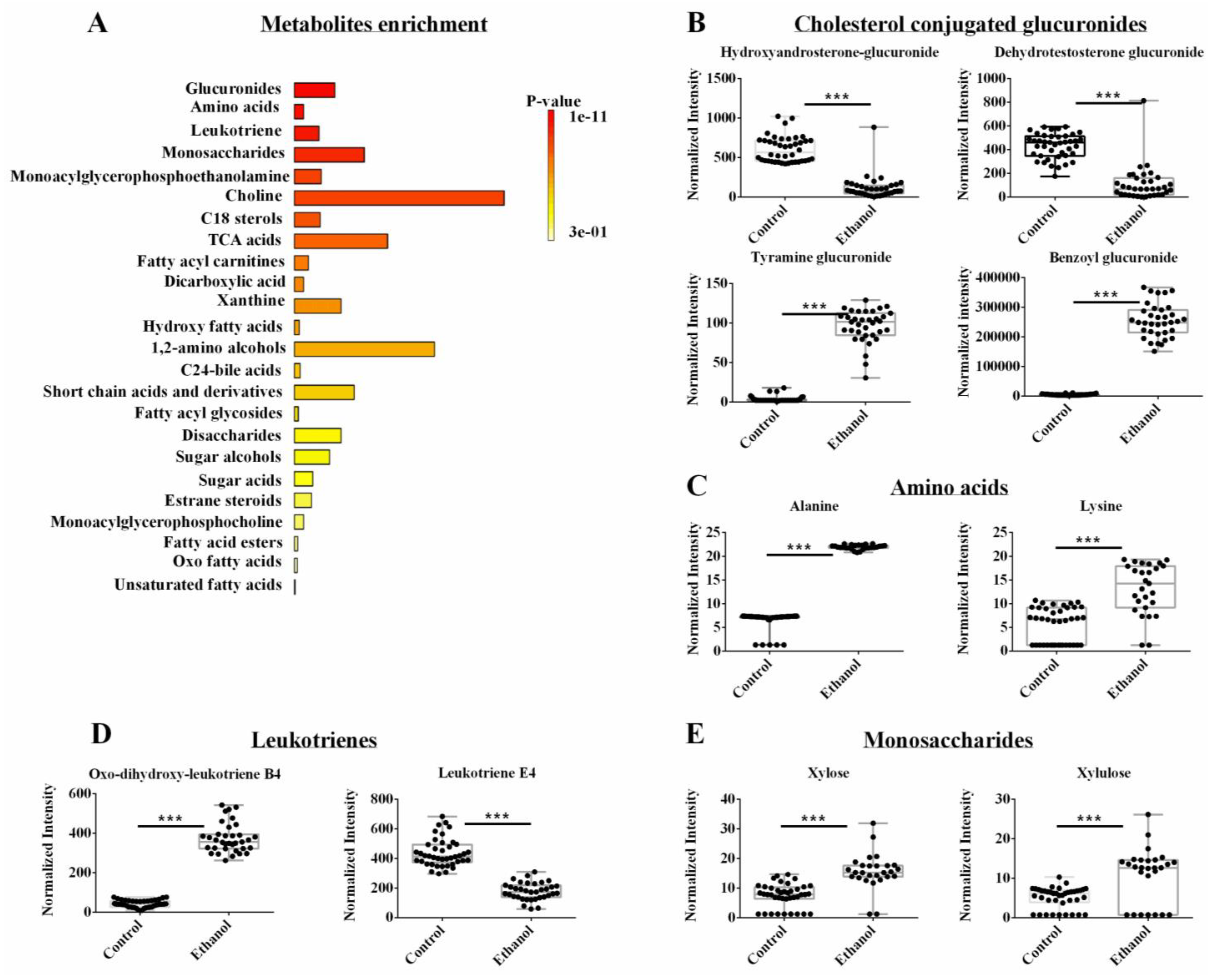
3.4. The Metabolomics Datasets Identified Glucuronides as Being Altered by Ethanol
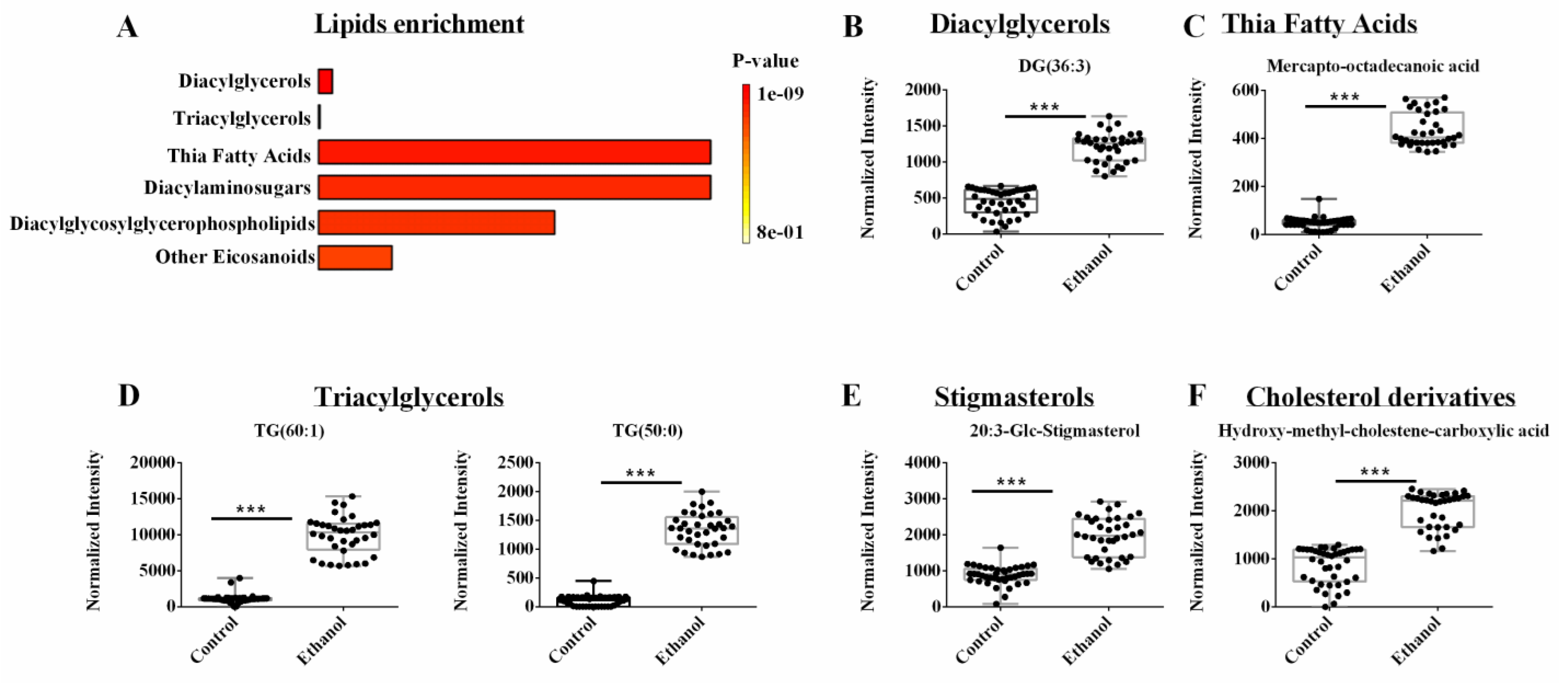
3.5. Lipidomics Reveals Lipids, Sterols, and Cholesterols Species Are Dysregulated by EtOH
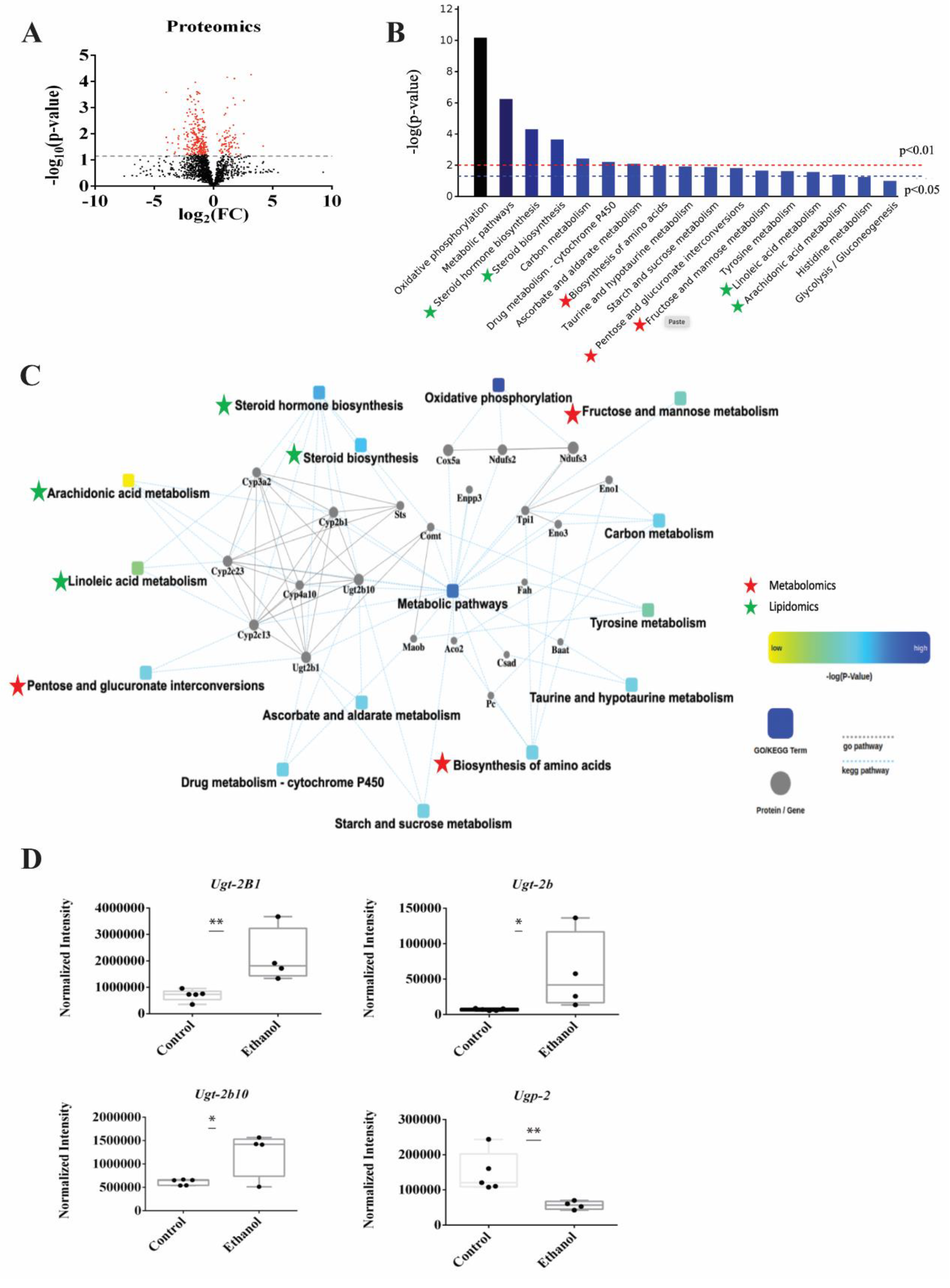
3.6. Proteomics Identified Glucuronidation as Being Significantly Altered in LDs Following Ethanol Treatment
3.7. The mRNA Levels of Ugt-1a1 Were Decreased but No Change in the Enzyme Activity for Pan UGTs Was Observed Following Ethanol Exposure
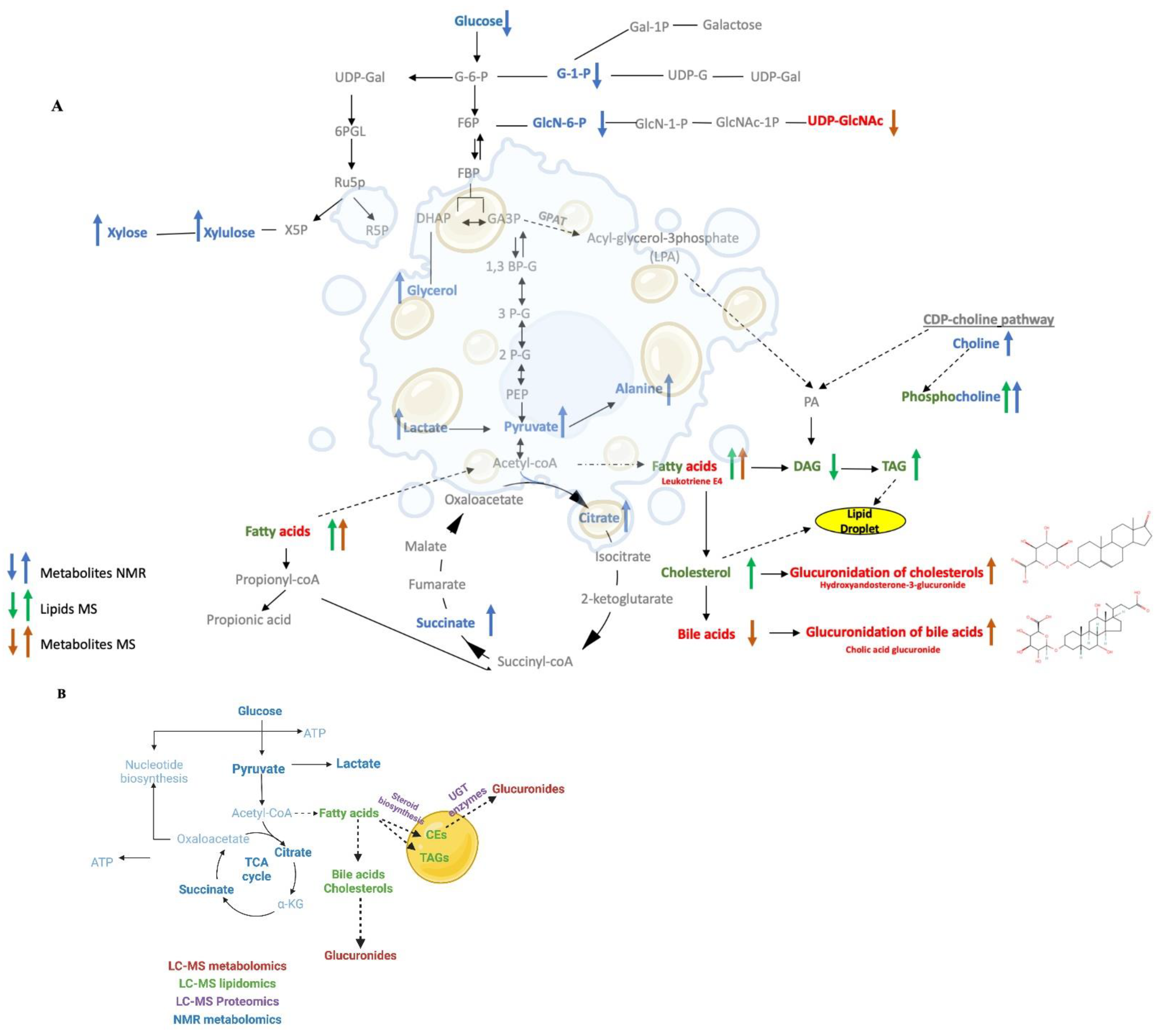
3.8. An Overall Network Map Shows Ethanol Administration Primarily Affected Glucuronides and Cholesterol Metabolism
4. Discussion
4.1. Ethanol Exposure Induced LD Accumulation and Metabolic Changes
4.2. Ethanol Exposure Induced Changes to the Fatty Acid Content of Hepatocytes
4.3. Ethanol Exposure Induced Hepatocytes to Sequester TAGs and Cholesterol in LDs
4.4. Ethanol Induced the Hydroxylation of Fatty Acids
4.5. Proteomics and Lipidomics Results Both Agree with the Accumulation of Cholesterol Synthesis in LDs
4.6. Only Metabolomics Identified Glucuronides as the Top-Altered Metabolites by EtOH
4.7. Ethanol Induced the Glucuronidation of Toxic Metabolites and the Upregulation of Glucuronosyltransferases UGT2s
4.8. A Network Map Revealed the Critical Interaction between UGTs and Cyps
4.9. Ethanol Feeding Directed Carbon Flow into Steroid Biosynthesis and Glucuronide Formation
4.10. The Upregulation of UGTs and the Accumulation of Fats and Steroids Induced LD Formation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bajaj, J.S. Alcohol, liver disease and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Rasineni, K.; Penrice, D.D.; Natarajan, S.K.; McNiven, M.A.; McVicker, B.L.; Kharbanda, K.K.; Casey, C.A.; Harris, E.N. Alcoholic vs non-alcoholic fatty liver in rats: Distinct differences in endocytosis and vesicle trafficking despite similar pathology. BMC. Gastroenterol. 2016, 16, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, A.; Muntane, J. Alcoholic liver disease, oxidative stress, and antioxidants. Rev. Esp. Enferm. Dig. 2011, 103, 393–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomes, P.G.; Rasineni, K.; Saraswathi, V.; Kharbanda, K.K.; Clemens, D.L.; Sweeney, S.A.; Kubik, J.L.; Donohue, T.M., Jr.; Casey, C.A. Natural Recovery by the Liver and Other Organs after Chronic Alcohol Use. Alcohol. Res. 2021, 41, 5. [Google Scholar] [CrossRef] [PubMed]
- Werner, J.; Saghir, M.; Warshaw, A.L.; Lewandrowski, K.B.; Laposata, M.; Iozzo, R.V.; Carter, E.A.; Schatz, R.J.; Fernandez-Del Castillo, C. Alcoholic pancreatitis in rats: Injury from nonoxidative metabolites of ethanol. Am. J. Physiol. Gastrointest. Liver. Physiol. 2002, 283, G65–G73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, C.P. Who gets alcoholic liver disease: Nature or nurture? J. R Coll. Physicians. Lond. 2000, 34, 557–562. [Google Scholar]
- Casey, C.A.; Macke, A.J.; Gough, R.R.; Pachikov, A.N.; Morris, M.E.; Thomes, P.G.; Kubik, J.L.; Holzapfel, M.S.; Petrosyan, A. Alcohol-Induced Liver Injury: Down-regulation and Redistribution of Rab3D Results in Atypical Protein Trafficking. Hepatol. Commun. 2022, 6, 374–388. [Google Scholar] [CrossRef]
- Casey, C.A.; Donohue, T.M., Jr.; Kubik, J.L.; Kumar, V.; Naldrett, M.J.; Woods, N.T.; Frisbie, C.P.; McNiven, M.A.; Thomes, P.G. Lipid droplet membrane proteome remodeling parallels ethanol-induced hepatic steatosis and its resolution. J. Lipid. Res. 2021, 62, 100049. [Google Scholar] [CrossRef]
- Zhou, Z.; Xu, M.J.; Gao, B. Hepatocytes: A key cell type for innate immunity. Cell Mol. Immunol. 2016, 13, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Schulze, R.J.; Schott, M.B.; Casey, C.A.; Tuma, P.L.; McNiven, M.A. The cell biology of the hepatocyte: A membrane trafficking machine. J. Cell Biol. 2019, 218, 2096–2112. [Google Scholar] [CrossRef]
- You, M.; Arteel, G.E. Effect of ethanol on lipid metabolism. J. Hepatol. 2019, 70, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Donohue, T.M., Jr.; Thomes, P.G. Ethanol-induced oxidant stress modulates hepatic autophagy and proteasome activity. Redox. Biol. 2014, 3, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sozen, E.; Demirel-Yalciner, T.; Sari, D.; Avcilar, C.; Samanci, T.F.; Ozer, N.K. Deficiency of SREBP1c modulates autophagy mediated lipid droplet catabolism during oleic acid induced steatosis. Metab. Open 2021, 12, 100138. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Fang, W.; Cui, K.; Chen, Q.; Xiang, X.; Zhang, J.; Zhang, Y.; Mai, K.; Ai, Q. Endoplasmic reticulum stress induces hepatic steatosis by transcriptional upregulating lipid droplet protein perilipin2. FASEB J. 2021, 35, e21900. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.; Zhang, J.; Davis, M.A.; Helsley, R.N.; Vedin, L.L.; Lee, R.G.; Crooke, R.M.; Graham, M.J.; Allende, D.S.; Parini, P.; et al. The lipid droplet-associated protein perilipin 3 facilitates hepatitis C virus-driven hepatic steatosis. J. Lipid. Res. 2017, 58, 420–432. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.K.; Seo, M.J.; Juvekar, V.; Jo, J.H.; Kim, W.; Choi, K.S.; Kim, H.M. Screening of Drug-Induced Steatosis and Phospholipidosis Using Lipid Droplet-Selective Two-Photon Probes. Anal. Chem. 2020, 92, 11223–11231. [Google Scholar] [CrossRef]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome. Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- Niu, L.; Sulek, K.; Vasilopoulou, C.G.; Santos, A.; Wewer Albrechtsen, N.J.; Rasmussen, S.; Meier, F.; Mann, M. Defining NASH from a Multi-Omics Systems Biology Perspective. J. Clin. Med. 2021, 10, 4673. [Google Scholar] [CrossRef]
- Beyoglu, D.; Idle, J.R. Metabolomic and Lipidomic Biomarkers for Premalignant Liver Disease Diagnosis and Therapy. Metabolites. 2020, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Iturrospe, E.; da Silva, K.M.; Robeyns, R.; van de Lavoir, M.; Boeckmans, J.; Vanhaecke, T.; van Nuijs, A.L.N.; Covaci, A. Metabolic Signature of Ethanol-Induced Hepatotoxicity in HepaRG Cells by Liquid Chromatography-Mass Spectrometry-Based Untargeted Metabolomics. J. Proteome. Res. 2022, 21, 1153–1166. [Google Scholar] [CrossRef]
- Fang, H.; Zhang, A.H.; Sun, H.; Yu, J.B.; Wang, L.; Wang, X.J. High-throughput metabolomics screen coupled with multivariate statistical analysis identifies therapeutic targets in alcoholic liver disease rats using liquid chromatography-mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1109, 112–120. [Google Scholar] [CrossRef]
- Manna, S.K.; Patterson, A.D.; Yang, Q.; Krausz, K.W.; Li, H.; Idle, J.R.; Fornace, A.J., Jr.; Gonzalez, F.J. Identification of noninvasive biomarkers for alcohol-induced liver disease using urinary metabolomics and the Ppara-null mouse. J. Proteome. Res. 2010, 9, 4176–4188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhotra, P.; Gill, R.K.; Saksena, S.; Alrefai, W.A. Disturbances in Cholesterol Homeostasis and Non-alcoholic Fatty Liver Diseases. Front. Med. 2020, 7, 467. [Google Scholar] [CrossRef]
- Olzmann, J.A.; Carvalho, P. Dynamics and functions of lipid droplets. Nat. Rev. Mol. Cell Biol. 2019, 20, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.E.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic. Acids. Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Worley, B.; Powers, R. MVAPACK: A complete data handling package for NMR metabolomics. ACS Chem. Biol. 2014, 9, 1138–1144. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids. Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Simpson, A.J.; Brown, S.A. Purge NMR: Effective and easy solvent suppression. J. Magn. Reson. 2005, 175, 340–346. [Google Scholar] [CrossRef]
- Geva, S.; Sitte, J. Adaptive nearest neighbor pattern classification. IEEE Trans. Neural Netw. Learn. Syst. 1991, 2, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Perperoglou, A.; Sauerbrei, W.; Abrahamowicz, M.; Schmid, M. A review of spline function procedures in R. BMC Med. Res. Methodol. 2019, 19, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emwas, A.H.; Saccenti, E.; Gao, X.; McKay, R.T.; Dos Santos, V.; Roy, R.; Wishart, D.S. Recommended strategies for spectral processing and post-processing of 1D (1)H-NMR data of biofluids with a particular focus on urine. Metabolomics 2018, 14, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, H.; Ji, F.; Chen, Y.; Cai, Z. statTarget: A streamlined tool for signal drift correction and interpretations of quantitative mass spectrometry-based omics data. Anal. Chim. Acta 2018, 1036, 66–72. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, Y.; Zhang, Y.; Li, F.; Xia, W.; Zhou, Y.; Qiu, Y.; Li, H.; Zhu, F. NOREVA: Enhanced normalization and evaluation of time-course and multi-class metabolomic data. Nucleic Acids Res. 2020, 48, W436–W448. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vazquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef]
- Crook, A.; De Lima Leite, A.; Payne, T.; Bhinderwala, F.; Woods, J.; Singh, V.K.; Powers, R. Radiation exposure induces cross-species temporal metabolic changes that are mitigated in mice by amifostine. Sci. Rep. 2021, 11, 14004. [Google Scholar] [CrossRef]
- Chatterjee, A.; Sakallioglu, I.T.; Murthy, D.; Kosmacek, E.A.; Singh, P.K.; McDonald, J.T.; Powers, R.; Oberley-Deegan, R.E. MnTE-2-PyP protects fibroblast mitochondria from hyperglycemia and radiation exposure. Redox Biol. 2022, 52, 102301. [Google Scholar] [CrossRef]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0--making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [Green Version]
- Kutsukake, T.; Furukawa, Y.; Ondo, K.; Gotoh, S.; Fukami, T.; Nakajima, M. Quantitative Analysis of UDP-Glucuronosyltransferase Ugt1a and Ugt2b mRNA Expression in the Rat Liver and Small Intestine: Sex and Strain Differences. Drug Metab. Dispos. 2019, 47, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.R.; Jankevics, A.; Weber, R.J.M. Struct: An R/bioconductor-based framework for standardised metabolomics data analysis and beyond. Bioinformatics 2020, 36, 5551–5552. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Dai, Z.; Liu, A.B.; Huang, J.; Narsipur, N.; Guo, G.; Kong, B.; Reuhl, K.; Lu, W.; Luo, Z.; et al. Intake of stigmasterol and β-sitosterol alters lipid metabolism and alleviates NAFLD in mice fed a high-fat western-style diet. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2018, 1863, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Shkumatov, A.A.; Alexander, S.T.; Ason, B.L.; Zhou, M. Stigmasterol accumulation causes cardiac injury and promotes mortality. Commun. Biol. 2019, 2, 20. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gu, Y.; Jiang, J.; Cui, X.; Cheng, S.; Liu, L.; Huang, Z.; Liao, R.; Zhao, P.; Yu, J.; et al. Stigmasterol attenuates hepatic steatosis in rats by strengthening the intestinal barrier and improving bile acid metabolism. NPJ. Sci. Food. 2022, 6, 38. [Google Scholar] [CrossRef]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Denise, N.S.; Hanspers, K.; Miller, R.A.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. WikiPathways: Connecting communities. Nucleic Acids Res. 2020, 49, D613–D621. [Google Scholar] [CrossRef]
- Seebacher, F.; Zeigerer, A.; Kory, N.; Krahmer, N. Hepatic lipid droplet homeostasis and fatty liver disease. Semin. Cell Dev. Biol. 2020, 108, 72–81. [Google Scholar] [CrossRef]
- Chen, K.; Li, J.; Wang, J.; Xia, Y.; Dai, W.; Wang, F.; Shen, M.; Cheng, P.; Zhang, Y.; Wang, C.; et al. 15-Deoxy- gamma 12,14-prostaglandin J2 Reduces Liver Impairment in a Model of ConA-Induced Acute Hepatic Inflammation by Activation of PPAR gamma and Reduction in NF- kappa B Activity. PPAR Res. 2014, 2014, 215631. [Google Scholar] [CrossRef] [Green Version]
- Skrede, S.; Sorensen, H.N.; Larsen, L.N.; Steineger, H.H.; Hovik, K.; Spydevold, O.S.; Horn, R.; Bremer, J. Thia fatty acids, metabolism and metabolic effects. Biochim. Biophys. Acta 1997, 1344, 115–131. [Google Scholar] [CrossRef]
- Kuzela, L.; Placer, Z. Liver steatosis and lipid peroxidation. Cesk. Gastroenterol. Vyz. 1968, 22, 380–383. [Google Scholar]
- Hu, C.; Wang, M.; Han, X. Shotgun lipidomics in substantiating lipid peroxidation in redox biology: Methods and applications. Redox. Biol. 2017, 12, 946–955. [Google Scholar] [CrossRef] [PubMed]
- Shirley, M.A.; Murphy, R.C. Metabolism of leukotriene B4 in isolated rat hepatocytes. Involvement of 2,4-dienoyl-coenzyme A reductase in leukotriene B4 metabolism. J. Biol. Chem. 1990, 265, 16288–16295. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.D.; Liu, J.Y. The Functions of Cytochrome P450 omega-hydroxylases and the Associated Eicosanoids in Inflammation-Related Diseases. Front. Pharm. 2021, 12, 716801. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T.; Murayama, N.; Umeyama, H.; Shimizu, M.; Yamazaki, H. Human liver enzymes responsible for metabolic elimination of tyramine; a vasopressor agent from daily food. Drug. Metab. Lett. 2011, 5, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Meech, R.; Rogers, A.; Zhuang, L.; Lewis, B.C.; Miners, J.O.; Mackenzie, P.I. Identification of residues that confer sugar selectivity to UDP-glycosyltransferase 3A (UGT3A) enzymes. J. Biol. Chem. 2012, 287, 24122–24130. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Sun, R.B.; Liao, X.Y.; Aa, J.Y.; Wang, G.J. UDP-glucuronosyltransferases (UGTs) and their related metabolic cross-talk with internal homeostasis: A systematic review of UGT isoforms for precision medicine. Pharm. Res. 2017, 121, 169–183. [Google Scholar] [CrossRef]
- Allain, E.P.; Rouleau, M.; Levesque, E.; Guillemette, C. Emerging roles for UDP-glucuronosyltransferases in drug resistance and cancer progression. Br. J. Cancer. 2020, 122, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Heier, C.; Xie, H.; Zimmermann, R. Nonoxidative ethanol metabolism in humans-from biomarkers to bioactive lipids. IUBMB Life 2016, 68, 916–923. [Google Scholar] [CrossRef] [Green Version]
- Sutliff, A.K.; Shi, J.; Watson, C.J.W.; Hunt, M.S.; Chen, G.; Zhu, H.J.; Lazarus, P. Potential Regulation of UGT2B10 and UGT2B7 by miR-485-5p in Human Liver. Mol. Pharm. 2019, 96, 674–682. [Google Scholar] [CrossRef]
- Kurzawski, M.; Szelag-Pieniek, S.; Lapczuk-Romanska, J.; Wrzesinski, M.; Oswald, S.; Drozdzik, M. The reference liver-CYP450 and UGT enzymes in healthy donor and metastatic livers: The impact of genotype. Pharm. Rep. 2022, 74, 204–215. [Google Scholar] [CrossRef]
- McCommis, K.S.; Finck, B.N. Mitochondrial pyruvate transport: A historical perspective and future research directions. Biochem. J. 2015, 466, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, J.M.; Kurkela, M.; Sonka, J.; Jäntti, S.; Ketola, R.; Bratton, S.; Finel, M.; Radominska-Pandya, A. Glucuronidation of oxidized fatty acids and prostaglandins B1 and E2 by human hepatic and recombinant UDP-glucuronosyltransferases. J. Lipid. Res. 2004, 45, 1694–1703. [Google Scholar] [CrossRef] [PubMed]
- Little, J.M.; Williams, L.; Xu, J.; Radominska-Pandya, A. Glucuronidation of the dietary fatty acids, phytanic acid and docosahexaenoic acid, by human UDP-glucuronosyltransferases. Drug. Metab Dispos. 2002, 30, 531–533. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Subclass | Metabolite or Lipid Name | HMDB or Lipid Maps ID | VIP Score | F.C. b | FDR p-Value c | MWW p-Value d |
|---|---|---|---|---|---|---|---|
| Fatty acyls | Fatty acids and conjugates | Hydroxyisovaleric acid | HMDB0000754 | 1.21 | 4.11 | 2.10 × 10−9 | 1.58 × 10−6 |
| Unsaturated fatty acids | Tricosenoic acid | LMFA01030091 | 1.20 | 0.17 | 1.74 × 10−15 | 1.71 × 10−22 | |
| Fatty acyl glycosides | Methyl-butenoyl-apiosylglucose | HMDB0039952 | 1.21 | 10.70 | 1.62 × 10−39 | 6.32 × 10−15 | |
| Fatty acyl glycosides | Gingerdiol -beta-glucopyranoside | HMDB0036123 | 1.20 | 8.88 | 5.84 × 10−46 | 7.74 × 10−17 | |
| Fatty acid esters | Dimethylnonanoyl carnitine | HMDB0006202 | 1.19 | 0.04 | 2.15 × 10−35 | 5.71 × 10−14 | |
| Glycerophos-pholipids | Glycerophosphoinositols | PI (36:4) | HMDB0009899 | 1.19 | 0.37 | 1.90 × 10−13 | 6.15 × 10−10 |
| Glycerophosphates | LysoPA (18:3) | HMDB0114743 | 1.19 | 4.60 | 1.17 × 10−36 | 5.67 × 10−9 | |
| Carbohydrates and carbohydrate conjugates | Glucose-1-phosphate | HMDB0001586 | 1.21 | 0.22 | 9.33 × 10−14 | 6.15 × 10−9 | |
| Pteridines and derivatives | Alloxazines and isoalloxazines | Riboflavin | HMDB0000244 | 1.21 | 8.18 | 9.29 × 10−50 | 2.00 × 10−22 |
| Saccharolipids | Diacylaminosugars | UDP-(beta-hydroxymyristoyl)-GlcNAc | LMSL01020003 | 1.19 | 0.15 | 8.85 × 10−11 | 1.70 × 10−17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakallioglu, I.T.; Tripp, B.; Kubik, J.; Casey, C.A.; Thomes, P.; Powers, R. Multiomics Approach Captures Hepatic Metabolic Network Altered by Chronic Ethanol Administration. Biology 2023, 12, 28. https://doi.org/10.3390/biology12010028
Sakallioglu IT, Tripp B, Kubik J, Casey CA, Thomes P, Powers R. Multiomics Approach Captures Hepatic Metabolic Network Altered by Chronic Ethanol Administration. Biology. 2023; 12(1):28. https://doi.org/10.3390/biology12010028
Chicago/Turabian StyleSakallioglu, Isin Tuna, Bridget Tripp, Jacy Kubik, Carol A. Casey, Paul Thomes, and Robert Powers. 2023. "Multiomics Approach Captures Hepatic Metabolic Network Altered by Chronic Ethanol Administration" Biology 12, no. 1: 28. https://doi.org/10.3390/biology12010028