

Bcl-xL Promotes the Survival of Motor Neurons Derived from Neural Stem Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Murine Neural Stem Cell Culture
2.2. Plasmid Construction
2.3. Generation of Stable mNSCs
2.4. Luciferase Assay
2.5. Motor Neuron Differentiation
2.6. Immunofluorescence
2.7. Isolation and Culture of Primary Neonatal Glial Cells
2.8. Replating-Induced Stress
2.9. Glutamate Toxicity Assay
2.10. Apoptosis Assay
2.11. Patch-Clamp Recordings
2.12. In Vivo Transplantation Studies
2.13. Statistical Analysis
3. Results
3.1. Generation of Directionally Inducible and Pro-Survival mNSCs
3.2. Ectopic Bcl-xL Does Not Affect Motor Neuron Differentiation
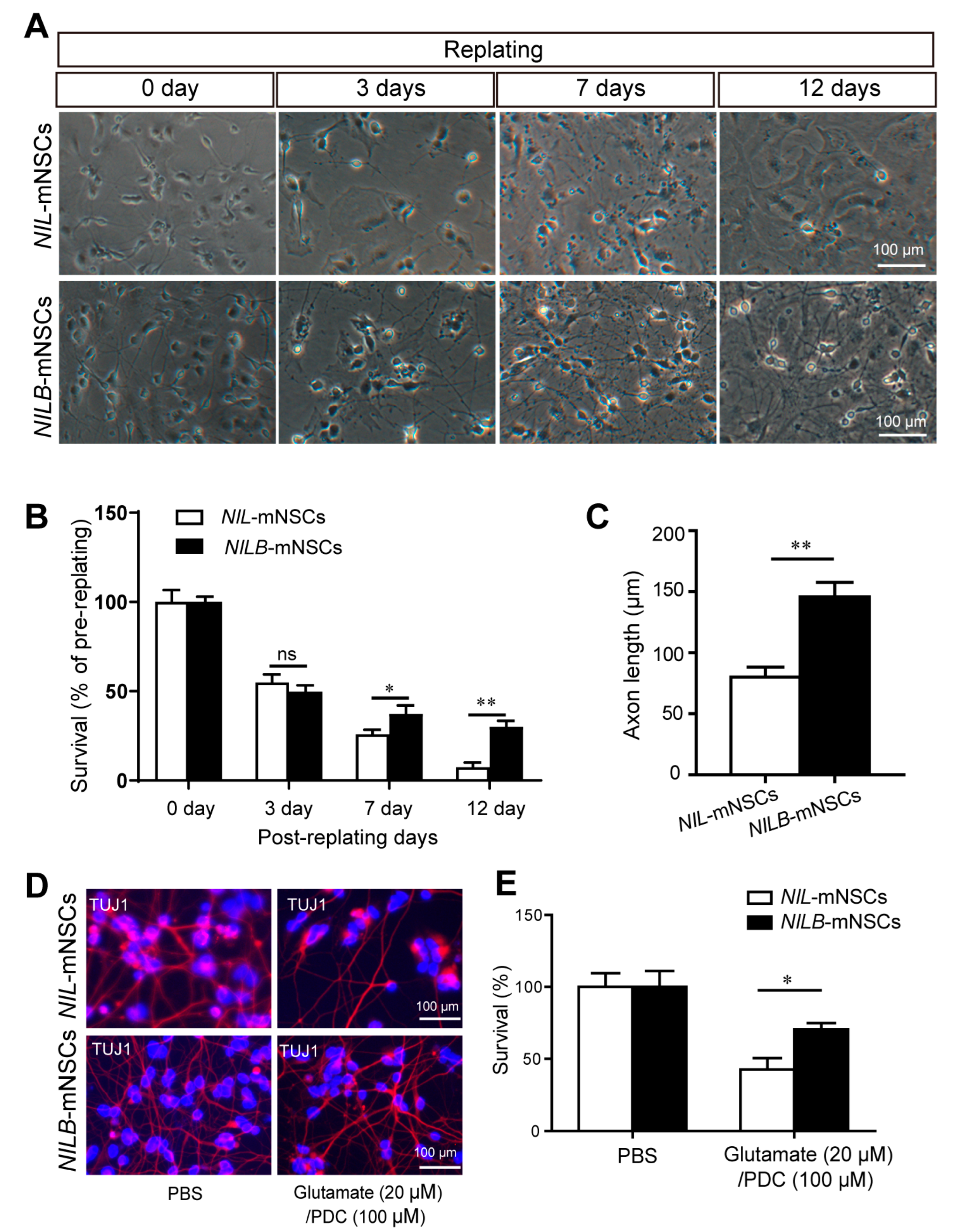
3.3. Ectopic Bcl-xL Promotes the Survival of Induced Motor Neurons
3.4. Motor Neurons Derived from NILB-mNSCs Exhibit Functional Membrane Properties
3.5. In Vivo Survival and Direct Differentiation of NILB-mNSCs Induced by Dox
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nijssen, J.; Comley, L.H.; Hedlund, E. Motor neuron vulnerability and resistance in amyotrophic lateral sclerosis. Acta Neuropathol. 2017, 133, 863–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciervo, Y.; Ning, K.; Jun, X.; Shaw, P.J.; Mead, R.J. Advances, challenges and future directions for stem cell therapy in amyotrophic lateral sclerosis. Mol. Neurodegener. 2017, 12, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trounson, A.; Thakar, R.G.; Lomax, G.; Gibbons, D. Clinical trials for stem cell therapies. BMC Med. 2011, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Petrou, P.; Gothelf, Y.; Argov, Z.; Gotkine, M.; Levy, Y.S.; Kassis, I.; Vaknin-Dembinsky, A.; Ben-Hur, T.; Offen, D.; Abramsky, O.; et al. Safety and Clinical Effects of Mesenchymal Stem Cells Secreting Neurotrophic Factor Transplantation in Patients with Amyotrophic Lateral Sclerosis: Results of Phase 1/2 and 2a Clinical Trials. JAMA Neurol. 2016, 73, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzini, L.; Fagioli, F.; Boccaletti, R.; Mareschi, K.; Oliveri, G.; Olivieri, C.; Pastore, I.; Marasso, R.; Madon, E. Stem cell therapy in amyotrophic lateral sclerosis: A methodological approach in humans. Amyotroph. Lateral Scler. Other Mot. Neuron Disord. 2003, 4, 158–161. [Google Scholar] [CrossRef]
- Mothe, A.; Tator, C.H. Isolation of Neural Stem/Progenitor Cells from the Periventricular Region of the Adult Rat and Human Spinal Cord. J. Vis. Exp. 2015, 99, e52732. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Xu, L.; Welsh, A.M.; Chen, D.; Hazel, T.; Johe, K.; Koliatsos, V.E. Combined immunosuppressive agents or CD4 antibodies prolong survival of human neural stem cell grafts and improve disease outcomes in amyotrophic lateral sclerosis transgenic mice. Stem Cells 2006, 24, 1976–1985. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, S.; Nishimura, S.; Iwai, H.; Sugai, K.; Zhang, L.; Shinozaki, M.; Iwanami, A.; Toyama, Y.; Liu, M.; Okano, H.; et al. Functional recovery from neural stem/progenitor cell transplantation combined with treadmill training in mice with chronic spinal cord injury. Sci. Rep. 2016, 6, 30898. [Google Scholar] [CrossRef] [Green Version]
- Ziavra, D.; Makri, G.; Giompres, P.; Taraviras, S.; Thomaidou, D.; Matsas, R.; Ada, M.; Elias, D.K. Neural stem cells transplanted in a mouse model of Parkinson’s disease differentiate to neuronal phenotypes and reduce rotational deficit. CNS Neurol. Disord. -Drug Targets 2012, 11, 829–835. [Google Scholar] [CrossRef]
- Swijnenburg, R.-J.; Schrepfer, S.; Govaert, J.A.; Cao, F.; Ransohoff, K.; Sheikh, A.Y.; Haddad, M.; Connolly, A.J.; Davis, M.M.; Robbins, R.C.; et al. Immunosuppressive therapy mitigates immunological rejection of human embryonic stem cell xenografts. Proc. Natl. Acad. Sci. USA 2008, 105, 12991–12996. [Google Scholar] [CrossRef]
- Zvibel, I.; Smets, F.; Soriano, H. Anoikis: Roadblock to cell transplantation? Cell Transpl. 2002, 11, 621–630. [Google Scholar] [CrossRef]
- Laflamme, M.; Chen, K.; Naumova, A.; Muskheli, V.; Fugate, J.; Dupras, S.; Reinecke, H.; Xu, C.; Hassanipour, M.; Police, S.; et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat. Biotechnol. 2007, 25, 1015–1024. [Google Scholar] [CrossRef]
- De Santis, R.; Garone, M.G.; Pagani, F.; de Turris, V.; Di Angelantonio, S.; Rosa, A. Direct conversion of human pluripotent stem cells into cranial motor neurons using a piggyBac vector. Stem Cell Res. 2018, 29, 189–196. [Google Scholar] [CrossRef]
- Rasmussen, M.A.; Hall, V.J.; Hyttel, P. Isolation and culture of porcine neural progenitor cells from embryos and pluripotent stem cells. Methods Mol Biol. 2013, 1074, 185–198. [Google Scholar]
- Chen, M.; Wang, X.; Li, C.; Lan, T.; Wei, Y.; Tang, C.; Zhou, X.; Zhou, R.; Rosa, A.; Zheng, X.; et al. Inducible motor neuron differentiation of human induced pluripotent stem cells in vivo. Cell Prolif. 2022, 55, e13319. [Google Scholar] [CrossRef]
- Vierbuchen, T.; Ostermeier, A.; Pang, Z.P.; Kokubu, Y.; Sudhof, T.C.; Wernig, M. Direct conversion of fibroblasts to functional neurons by defined factors. Nature 2010, 463, 1035–1041. [Google Scholar] [CrossRef] [Green Version]
- Hedlund, E.; Karlsson, M.; Osborn, T.; Ludwig, W.; Isacson, O. Global gene expression profiling of somatic motor neuron popu-lations with different vulnerability identify molecules and pathways of degeneration and protection. Brain 2010, 133, 2313–2330. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.L.; Zang, T.; Zhang, C.L. Direct Lineage Reprogramming Reveals Disease-Specific Phenotypes of Motor Neurons from Human ALS Patients. Cell Rep. 2016, 14, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Rothstein, J.D.; Van Kammen, M.; Levey, A.I.; Martin, L.J.; Kuncl, R.W. Selective loss of glial glutamate transporter GLT-1 in amy-otrophic lateral sclerosis. Ann. Neurol. 1995, 38, 73–84. [Google Scholar]
- Müller-Ehmsen, J.; Krausgrill, B.; Burst, V.; Schenk, K.; Neisen, U.; Fries, J.; Fleischmann, B.; Hescheler, J.; Robert, H.G. SchwingerEffective engraftment but poor mid-term per-sistence of mononuclear and mesenchymal bone marrow cells in acute and chronic rat myocardial infarction. J. Mol. Cell Cardiol. 2006, 41, 876–884. [Google Scholar] [CrossRef]
- Skuk, D.; Caron, N.J.; Goulet, M.; Roy, B.; Tremblay, J.P. Resetting the problem of cell death following muscle-derived cell trans-plantation: Detection, dynamics and mechanisms. J. Neuropathol. Exp. Neurol. 2003, 62, 951–967. [Google Scholar]
- Emgard, M.; Hallin, U.; Karlsson, J.; Bahr, B.A.; Brundin, P.; Blomgren, K. Both apoptosis and necrosis occur early after intracerebral grafting of ventral mesencephalic tissue: A role for protease activation. J. Neurochem. 2003, 86, 1223–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Pei, W.; Ge, H.; Liang, Q.; Luo, Y.; Sharp, F.R.; Lu, A.; Ran, R.; Graham, S.H.; Chen, J. In vivo delivery of a Bcl-xL fusion protein containing the TAT protein transduction domain protects against ischemic brain injury and neuronal apoptosis. J. Neurosci. 2002, 22, 5423–5431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, N.A.; Fisher, J.K.; Austgen, K.; VandenBerg, S.; Huang, E.J.; Oakes, S.A. Blocking the mitochondrial apoptotic pathway preserves motor neuron viability and function in a mouse model of amyotrophic lateral sclerosis. J. Clin. Investig. 2010, 120, 3673–3679. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.-W.; Koh, H.-C.; Chang, M.-Y.; Roh, E.; Choi, C.-Y.; Oh, Y.; Son, H.; Lee, Y.-S.; Studer, L.; Lee, S.-H. Enhanced in vitro midbrain dopamine neuron differentiation, dopaminergic function, neurite outgrowth, and 1-methyl-4-phenylpyridium resistance in mouse embryonic stem cells overexpressing Bcl-XL. J. Neurosci. 2004, 24, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Liste, I.; Garcia-Garcia, E.; Bueno, C.; Martinez-Serrano, A. Bcl-XL modulates the differentiation of immortalized human neural stem cells. Cell Death Differ. 2007, 14, 1880–1892. [Google Scholar] [CrossRef] [Green Version]
- Bas, J.; Nguyen, T.; Gillet, G. Involvement of bcl-xL in neuronal function and development. Int. J. Mol. Sci. 2021, 22, 3202. [Google Scholar] [CrossRef]
- Sances, S.; Bruijn, L.; Chandran, S.; Eggan, K.; Ho, R.; Klim, J.; Livesey, M.; Lowry, E.; Macklis, J.; Rushton, D.; et al. Modeling ALS with motor neurons derived from human induced pluripotent stem cells. Nat. Neurosci. 2016, 19, 542–553. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Chanda, S.; Marro, S.; Ng, Y.-H.; Janas, J.; Haag, D.; Ang, C.E.; Tang, Y.; Flores, Q.; Mall, M.; et al. Generation of pure GABAergic neurons by transcription factorprogramming. Nat. Methods 2017, 14, 621–628. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Pak, C.; Han, Y.; Ahlenius, H.; Zhang, Z.; Chanda, S.; Marro, S.; Patzke, C.; Acuna, C.; Covy, J.; et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron 2013, 78, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Cuvillier, J.M.; Lee, B.; Shen, R.; Lee, J.W.; Lee, S.K. Fusion protein Isl1-Lhx3 specifies motor neuron fate by inducing motor neuron genes and concomitantly suppressing the interneuron programs. Proc. Natl. Acad. Sci. USA 2012, 109, 3383–3388. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.-L.; Zang, T.; Zou, Y.; Chang, J.; Gibson, J.; Huber, K.; Zhang, C.-L. Small molecules enable neurogenin 2 to efficiently convert human fibroblasts into cholinergic neurons. Nat. Commun. 2013, 4, 2183. [Google Scholar] [CrossRef]
- Goto, K.; Imamura, K.; Komatsu, K.; Mitani, K.; Aiba, K.; Nakatsuji, N.; Inoue, M.; Kawata, A.; Yamashita, H.; Takahashi, R.; et al. Simple Derivation of Spinal Motor Neurons from ESCs/iPSCs Using Sendai Virus Vectors. Mol. Ther. Methods Clin. Dev. 2017, 4, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Wichterle, H.; Lieberam, I.; Porter, J.A.; Jessell, T.M. Directed differentiation of embryonic stem cells into motor neurons. Cell 2002, 110, 385–397. [Google Scholar] [CrossRef] [Green Version]
- Barth, L.; Sütterlin, R.; Nenniger, M.; Vogt, K.E. Functional differentiation of stem cell-derived neurons from different murine backgrounds. Front. Cell Neurosci. 2014, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Yan, J.; Chen, D.; Welsh, A.M.; Hazel, T.; Johe, K.; Hatfield, G.; Koliatsos, V.E. Human neural stem cell grafts ameliorate motor neuron disease in SOD-1 transgenic rats. Transplantation 2006, 82, 865–875. [Google Scholar] [CrossRef]
- Fischer, I.; Dulin, J.N.; Lane, M.A. Transplanting neural progenitor cells to restore connectivity after spinal cord injury. Nat. Rev. Neurosci. 2020, 21, 366–383. [Google Scholar] [CrossRef]
- Phillips, M.I.; Tang, Y.L. Genetic modification of stem cells for transplantation. Adv. Drug Deliv. Rev. 2008, 60, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, C.D.; Slotkin, J.; Yu, D.; Dai, H.; Lawrence, M.; Bronson, R.; Reynolds, F.M.; Teng, Y.; Woodard, E.; Langer, R. Establishing a model spinal cord injury in the African green monkey for the preclinical evaluation of biodegradable polymer scaffolds seeded with human neural stem cells. J. Neurosci. Meth. 2010, 188, 258–269. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Peng, X.; Ang, S.; Gao, Y.; Chi, Y.; Wang, J.; Tang, C.; Zhou, X.; Feng, Y.; Zhang, K.; et al. Bcl-xL Promotes the Survival of Motor Neurons Derived from Neural Stem Cells. Biology 2023, 12, 132. https://doi.org/10.3390/biology12010132
Wu Y, Peng X, Ang S, Gao Y, Chi Y, Wang J, Tang C, Zhou X, Feng Y, Zhang K, et al. Bcl-xL Promotes the Survival of Motor Neurons Derived from Neural Stem Cells. Biology. 2023; 12(1):132. https://doi.org/10.3390/biology12010132
Chicago/Turabian StyleWu, Yunqin, Xiaohua Peng, Song Ang, Yue Gao, Yue Chi, Jinling Wang, Chengcheng Tang, Xiaoqing Zhou, Yanxian Feng, Kun Zhang, and et al. 2023. "Bcl-xL Promotes the Survival of Motor Neurons Derived from Neural Stem Cells" Biology 12, no. 1: 132. https://doi.org/10.3390/biology12010132