

Detecting Intestinal Goblet Cells of the Broadgilled Hagfish Eptatretus cirrhatus (Forster, 1801): A Confocal Microscopy Evaluation
 , , ,
, , ,  , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Tissue Preparation
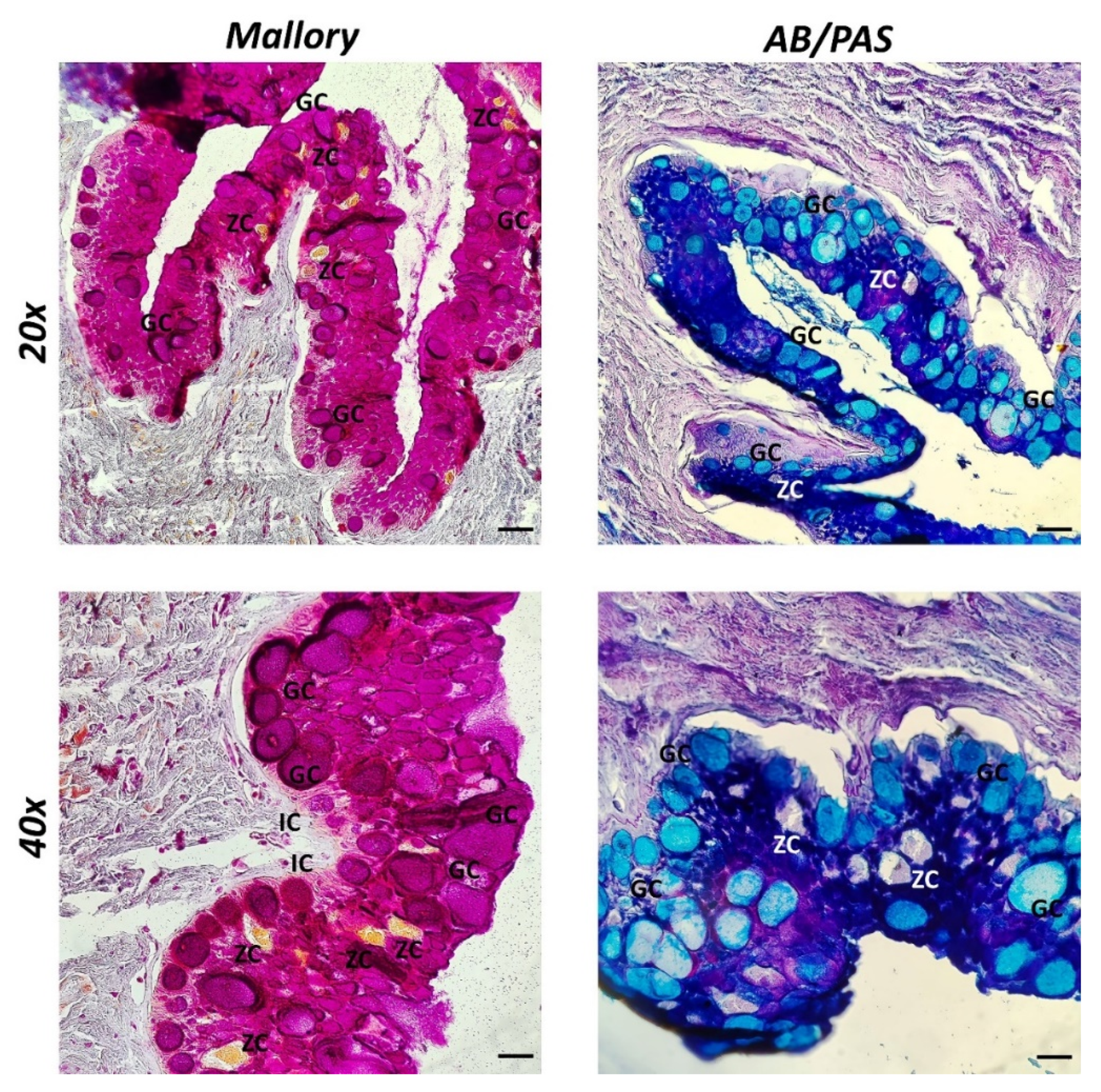
2.3. Histology and Histochemistry
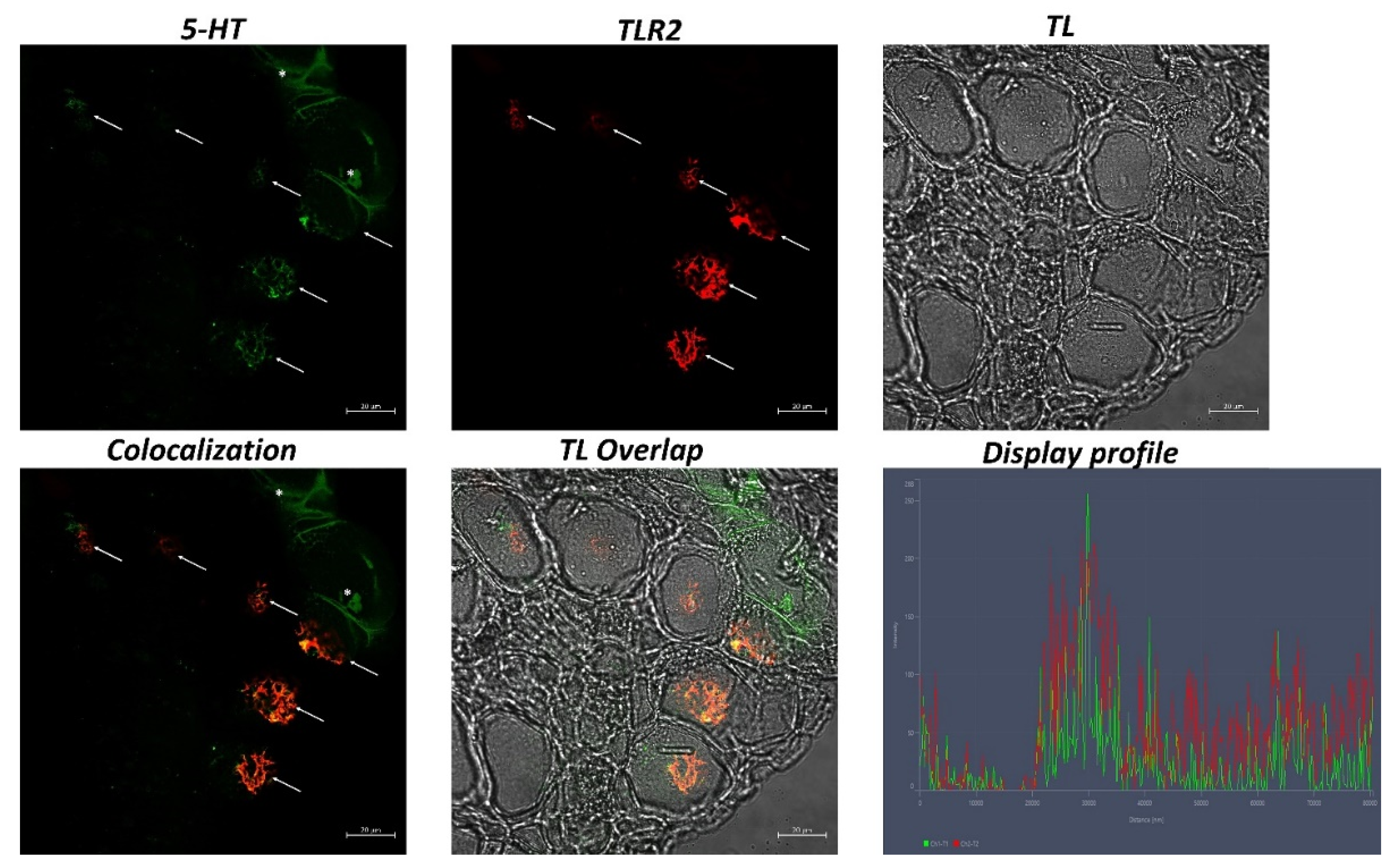
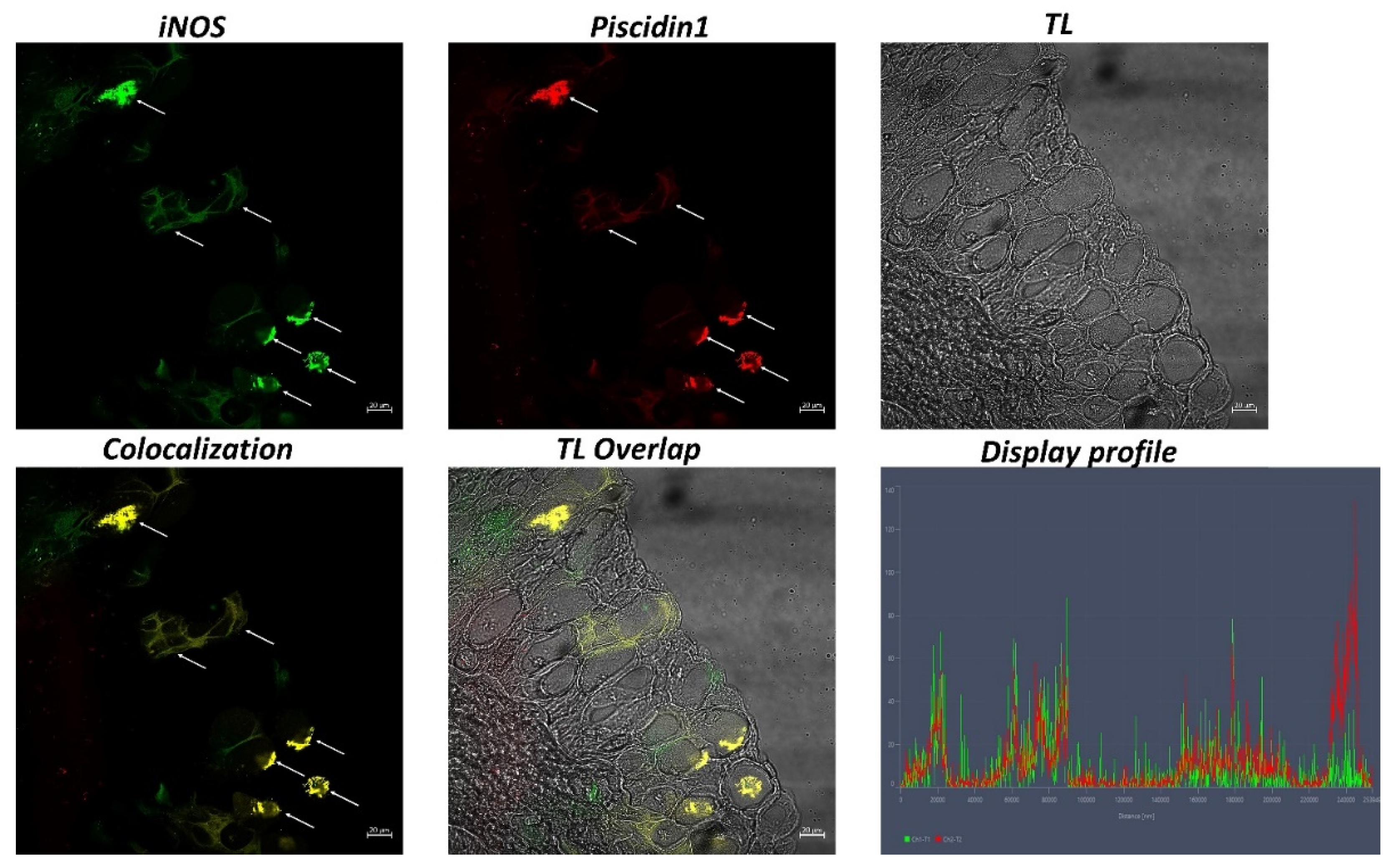
2.4. Immunofluorescence
2.5. Laser Confocal Immunofluorescence
2.6. Quantitative Analysis
2.7. Phylogenetic Conservation of Primary Antibodies
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Su, Y.-N.; Wu, P.; Feng, L.; Jiang, W.-D.; Jiang, J.; Zhang, Y.-A.; Figueiredo-Silva, C.; Zhou, X.-Q.; Liu, Y. The Improved Growth Performance and Enhanced Immune Function by DL-Methionyl-DL-Methionine Are Associated with NF-ΚB and TOR Signalling in Intestine of Juvenile Grass Carp (Ctenopharyngodon Idella). Fish Shellfish Immunol. 2018, 74, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Pergolizzi, S.; Fumia, A.; Calabrò, C.; Lo Cascio, P.; Lauriano, E.R. Mast Cells in Goldfish (Carassius Auratus) Gut: Immunohistochemical Characterization. Acta Zool. 2022, 85, 2651–2658. [Google Scholar] [CrossRef]
- Segner, H.; Bailey, C.; Tafalla, C.; Bo, J. Immunotoxicity of Xenobiotics in Fish: A Role for the Aryl Hydrocarbon Receptor (AhR)? Int. J. Mol. Sci. 2021, 22, 9460. [Google Scholar] [CrossRef] [PubMed]
- Peatman, E.; Lange, M.; Zhao, H.; Beck, B.H. Physiology and Immunology of Mucosal Barriers in Catfish (Ictalurus Spp.). Tissue Barriers 2015, 3, e1068907. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The Mucosal Immune System of Fish: The Evolution of Tolerating Commensals While Fighting Pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Ou, W.; Hu, H.; Yang, P.; Dai, J.; Ai, Q.; Zhang, W.; Zhang, Y.; Mai, K. Dietary Daidzein Improved Intestinal Health of Juvenile Turbot in Terms of Intestinal Mucosal Barrier Function and Intestinal Microbiota. Fish Shellfish Immunol. 2019, 94, 132–141. [Google Scholar] [CrossRef]
- Rombout, J.H.W.M.; Yang, G.; Kiron, V. Adaptive Immune Responses at Mucosal Surfaces of Teleost Fish. Fish Shellfish Immunol. 2014, 40, 634–643. [Google Scholar] [CrossRef]
- Ryo, S.; Wijdeven, R.H.M.; Tyagi, A.; Hermsen, T.; Kono, T.; Karunasagar, I.; Rombout, J.H.W.M.; Sakai, M.; Kemenade, B.M.L.V.; Savan, R. Common Carp Have Two Subclasses of Bonyfish Specific Antibody IgZ Showing Differential Expression in Response to Infection. Dev. Comp. Immunol. 2010, 34, 1183–1190. [Google Scholar] [CrossRef]
- Yang, S.; Yu, M. Role of Goblet Cells in Intestinal Barrier and Mucosal Immunity. J. Inflamm. Res. 2021, 14, 3171–3183. [Google Scholar] [CrossRef]
- Birchenough, G.M.H.; Nyström, E.E.L.; Johansson, M.E.V.; Hansson, G.C. A Sentinel Goblet Cell Guards the Colonic Crypt by Triggering Nlrp6-Dependent Muc2 Secretion. Science 2016, 352, 1535–1542. [Google Scholar] [CrossRef] [Green Version]
- McKernan, D.P. Toll-like Receptors and Immune Cell Crosstalk in the Intestinal Epithelium. AIMS Allergy Immunol. 2019, 3, 13–31. [Google Scholar] [CrossRef]
- Knoop, K.A.; Newberry, R.D. Goblet Cells: Multifaceted Players in Immunity at Mucosal Surfaces. Mucosal Immunol. 2018, 11, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Zaccone, G.; Capillo, G.; Fernandes, J.M.O.; Kiron, V.; Lauriano, E.R.; Alesci, A.; Lo Cascio, P.; Guerrera, M.C.; Kuciel, M.; Zuwala, K.; et al. Expression of the Antimicrobial Peptide Piscidin 1 and Neuropeptides in Fish Gill and Skin: A Potential Participation in Neuro-Immune Interaction. Mar. Drugs 2022, 20, 145. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Capillo, G.; Cascio, P.L.; Lauriano, E.R. Rodlet Cells in Kidney of Goldfish (Carassius Auratus, Linnaeus 1758): A Light and Confocal Microscopy Study. Acta Histochem. 2022, 124, 151876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wu, C. The Relationship between Intestinal Goblet Cells and the Immune Response. Biosci. Rep. 2020, 40, BSR20201471. [Google Scholar] [CrossRef]
- McDole, J.R.; Wheeler, L.W.; McDonald, K.G.; Wang, B.; Konjufca, V.; Knoop, K.A.; Newberry, R.D.; Miller, M.J. Goblet Cells Deliver Luminal Antigen to CD103+ Dendritic Cells in the Small Intestine. Nature 2012, 483, 345–349. [Google Scholar] [CrossRef]
- Engevik, M.A.; Chang-Graham, A.; Hyser, J.M.; Versalovic, J. Serotonin Promotes Epithelial Restitution through Goblet Cell Mediated Secretion of Muc2 and TFF3. FASEB J. 2019, 33, 869.1. [Google Scholar] [CrossRef]
- Uyttebroek, L.; Van Remoortel, S.; Buyssens, L.; Popowycz, N.; Hubens, G.; Timmermans, J.-P.; van Nassauw, L. The Effect of Diet Induced Obesity on Serotonin in Zebrafish. J. Cell. Signal. 2022, 3, 115–128. [Google Scholar]
- Losada, A.P.; Bermúdez, R.; Faílde, L.D.; Quiroga, M.I. Quantitative and Qualitative Evaluation of INOS Expression in Turbot (Psetta Maxima) Infected with Enteromyxum Scophthalmi. Fish Shellfish Immunol. 2012, 32, 243–248. [Google Scholar] [CrossRef]
- Mistri, A.; Kumari, U.; Mittal, S.; Mittal, A.K. Immunohistochemical Localization of Nitric Oxide Synthase (NOS) Isoforms in Epidermis and Gill Epithelium of an Angler Catfish, Chaca Chaca (Siluriformes, Chacidae). Tissue Cell 2018, 55, 25–30. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Żuwała, K.; Kuciel, M.; Budzik, K.A.; Capillo, G.; Alesci, A.; Pergolizzi, S.; Dugo, G.; Zaccone, G. Confocal Immunohistochemistry of the Dermal Glands and Evolutionary Considerations in the Caecilian, Typhlonectes Natans (Amphibia: Gymnophiona). Acta Zool. 2016, 97, 154–164. [Google Scholar] [CrossRef]
- Alesci, A.; Capillo, G.; Fumia, A.; Messina, E.; Albano, M.; Aragona, M.; Lo Cascio, P.; Spanò, N.; Pergolizzi, S.; Lauriano, E.R. Confocal Characterization of Intestinal Dendritic Cells from Myxines to Teleosts. Biology 2022, 11, 1045. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Cicero, N.; Fumia, A.; Petrarca, C.; Mangifesta, R.; Nava, V.; Lo Cascio, P.; Gangemi, S.; Di Gioacchino, M.; Lauriano, E.R. Histological and Chemical Analysis of Heavy Metals in Kidney and Gills of Boops Boops: Melanomacrophages Centers and Rodlet Cells as Environmental Biomarkers. Toxics 2022, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Pergolizzi, S.; Rizzo, G.; Favaloro, A.; Alesci, A.; Pallio, S.; Melita, G.; Cutroneo, G.; Lauriano, E.R. Expression of VAChT and 5-HT in Ulcerative Colitis Dendritic Cells. Acta Histochem. 2021, 123, 151715. [Google Scholar] [CrossRef] [PubMed]
- Zaccone, G.; Fudge, D.S.; Winegard, T.M.; Capillo, G.; Kuciel, M.; Funakoshi, K.; Lauriano, E.R. Confocal Imaging and Phylogenetic Considerations of the Subcutaneous Neurons in the Atlantic Hagfish Myxine Glutinosa. Acta Zool. 2015, 96, 209–217. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Backström, T.; Winberg, S. Serotonin Coordinates Responses to Social Stress—What We Can Learn from Fish. Front. Neurosci. 2017, 11, 595. [Google Scholar] [CrossRef] [PubMed]
- Kaizuka, T.; Hayashi, T. Comparative Analysis of Palmitoylation Sites of Serotonin (5-HT) Receptors in Vertebrates. Neuropsychopharmacol. Rep. 2018, 38, 75–85. [Google Scholar] [CrossRef]
- Yamaguchi, F.; Brenner, S. Molecular Cloning of 5-Hydroxytryptamine (5-HT) Type 1 Receptor Genes from the Japanese Puffer Fish, Fugu Rubripes. Gene 1997, 191, 219–223. [Google Scholar] [CrossRef]
- Schneider, H.; Fritzky, L.; Williams, J.; Heumann, C.; Yochum, M.; Pattar, K.; Noppert, G.; Mock, V.; Hawley, E. Cloning and Expression of a Zebrafish 5-HT2C Receptor Gene. Gene 2012, 502, 108–117. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Lo Cascio, P.; Fumia, A.; Lauriano, E.R. Neuronal Regeneration: Vertebrates Comparative Overview and New Perspectives for Neurodegenerative Diseases. Acta Zool. 2021, 103, 129–140. [Google Scholar] [CrossRef]
- Marino, A.; Pergolizzi, S.; Lauriano, E.R.; Santoro, G.; Spataro, F.; Cimino, F.; Speciale, A.; Nostro, A.; Bisignano, G. TLR2 Activation in Corneal Stromal Cells by Staphylococcus Aureus -Induced Keratitis. APMIS 2015, 123, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Aragona, M.; Cicero, N.; Lauriano, E.R. Can Nutraceuticals Assist Treatment and Improve Covid-19 Symptoms? Nat. Prod. Res. 2021, 36, 2672–2691. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Lauriano, E.R.; Fumia, A.; Irrera, N.; Mastrantonio, E.; Vaccaro, M.; Gangemi, S.; Santini, A.; Cicero, N.; Pergolizzi, S. Relationship between Immune Cells, Depression, Stress, and Psoriasis: Could the Use of Natural Products Be Helpful? Molecules 2022, 27, 1953. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Lo Cascio, P.; Capillo, G.; Lauriano, E.R. Localization of Vasoactive Intestinal Peptide and Toll-like Receptor 2 Immunoreactive Cells in Endostyle of Urochordate Styela plicata (Lesueur, 1823). Microsc. Res. Tech. 2022, 85, 2651–2658. [Google Scholar] [CrossRef]
- Anandhakumar, C.; Lavanya, V.; Pradheepa, G.; Tirumurugaan, K.G.; Dhinakar Raj, G.; Raja, A.; Pazhanivel, N.; Balachandran, C. Expression Profile of Toll-like Receptor 2 MRNA in Selected Tissues of Shark (Chiloscyllium Sp.). Fish Shellfish Immunol. 2012, 33, 1174–1182. [Google Scholar] [CrossRef]
- Jault, C.; Pichon, L.; Chluba, J. Toll-like Receptor Gene Family and TIR-Domain Adapters in Danio Rerio. Mol. Immunol. 2004, 40, 759–771. [Google Scholar] [CrossRef]
- Oshiumi, H.; Matsumoto, M.; Funami, K.; Akazawa, T.; Seya, T. TICAM-1, an Adaptor Molecule That Participates in Toll-like Receptor 3–Mediated Interferon-β Induction. Nat. Immunol. 2003, 4, 161–167. [Google Scholar] [CrossRef]
- Steinert, J.R.; Chernova, T.; Forsythe, I.D. Nitric Oxide Signaling in Brain Function, Dysfunction, and Dementia. Neuroscientist 2010, 16, 435–452. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric Oxide Synthases: Regulation and Function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef]
- Moncada, S.; Higgs, E.A. The Discovery of Nitric Oxide and Its Role in Vascular Biology: The Discovery of Nitric Oxide and Its Role in Vascular Biology. Br. J. Pharmacol. 2006, 147, S193–S201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namba, T.; Koike, H.; Murakami, K.; Aoki, M.; Makino, H.; Hashiya, N.; Ogihara, T.; Kaneda, Y.; Kohno, M.; Morishita, R. Angiogenesis Induced by Endothelial Nitric Oxide Synthase Gene Through Vascular Endothelial Growth Factor Expression in a Rat Hindlimb Ischemia Model. Circulation 2003, 108, 2250–2257. [Google Scholar] [CrossRef] [PubMed]
- Annona, G.; Sato, I.; Pascual-Anaya, J.; Braasch, I.; Voss, R.; Stundl, J.; Soukup, V.; Kuratani, S.; Postlethwait, J.H.; D’Aniello, S. Evolution of the Nitric Oxide Synthase Family in Vertebrates and Novel Insights in Gill Development. Proc. R. Soc. B. 2022, 289, 20220667. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, F.; Murakami, Y.; Kuratani, S. Gene Expression Analysis of Lamprey Embryos. In In Situ Hybridization Methods; Hauptmann, G., Ed.; Neuromethods; Springer: New York, NY, USA, 2015; Volume 99, pp. 263–278. ISBN 9781493923021. [Google Scholar]
- Saeij, J.P.J.; Stet, R.J.M.; Groeneveld, A.; Verburg-van Kemenade, L.B.M.; van Muiswinkel, W.B.; Wiegertjes, G.F. Molecular and Functional Characterization of a Fish Inducible-Type Nitric Oxide Synthase. Immunogenetics 2000, 51, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-M.; Zhou, X.-M.; Zhou, Y.-L.; Kuang, W.-M.; Chen, Y.-J.; Luo, L.; Dai, F.-Y. Intestinal Morphology, Immunity and Microbiota Response to Dietary Fibers in Largemouth Bass, Micropterus Salmoide. Fish Shellfish Immunol. 2020, 103, 135–142. [Google Scholar] [CrossRef]
- Zhang, H.; Ran, C.; Teame, T.; Ding, Q.; Hoseinifar, S.H.; Xie, M.; Zhang, Z.; Yang, Y.; Olsen, R.E.; Gatlin, D.M. Research Progress on Gut Health of Farmers Teleost Fish: A Viewpoint Concerning the Intestinal Mucosal Barrier and the Impact of Its Damage. Rev. Fish Biol. Fish. 2020, 30, 569–586. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ho, S.B. Intestinal Goblet Cells and Mucins in Health and Disease: Recent Insights and Progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef]
- Gieryńska, M.; Szulc-Dąbrowska, L.; Struzik, J.; Mielcarska, M.B.; Gregorczyk-Zboroch, K.P. Integrity of the Intestinal Barrier: The Involvement of Epithelial Cells and Microbiota—A Mutual Relationship. Animals 2022, 12, 145. [Google Scholar] [CrossRef]
- Dawood, M.A. Nutritional Immunity of Fish Intestines: Important Insights for Sustainable Aquaculture. Rev. Aquac. 2021, 13, 642–663. [Google Scholar] [CrossRef]
- Da Cruz, T.M.; Moretti, D.B.; Nordi, W.M.; Cyrino, J.E.; Machado-Neto, R. Dietary Lyophilized Colostrum Alters Distribution of Goblet Cells and the Intestinal Epithelium of Piaractus Mesopotamicus. Aquaculture 2017, 468, 286–292. [Google Scholar] [CrossRef]
- Weinrauch, A.; Edwards, S.; Goss, G. Anatomy of the Pacific Hagfish (Epatatretus Stoutii). In Hagfish Biology; CRC Press: Boca Raton, FL, USA, 2015; pp. 1–39. ISBN 9781482233452. [Google Scholar]
- Koopman, N.; Katsavelis, D.; Ten Hove, A.S.; Brul, S.; de Jonge, W.J.; Seppen, J. The Multifaceted Role of Serotonin in Intestinal Homeostasis. Int. J. Mol. Sci. 2021, 22, 9487. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, N.d.N.; Firmiano, E.M.d.S.; Gomes, I.D.; Nascimento, A.A.d.; Sales, A.; Araújo, F.G. Histochemical and Immunohistochemical Study on Endocrine Cells (5HT, GAS, and SST) of the Gastrointestinal Tract of a Teleost, the Characin Astyanax Bimaculatus. Acta Histochem. 2015, 117, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.R.; Capillo, G.; Icardo, J.M.; Fernandes, J.M.O.; Kiron, V.; Kuciel, M.; Zuwala, K.; Guerrera, M.C.; Aragona, M.; Zaccone, G. Neuroepithelial Cells (NECs) and Mucous Cells Express a Variety of Neurotransmitters and Neurotransmitter Receptors in the Gill and Respiratory Air-Sac of the Catfish Heteropneustes Fossilis (Siluriformes, Heteropneustidae): A Possible Role in Local Immune Defence. Zoology 2021, 148, 125958. [Google Scholar] [CrossRef] [PubMed]
- Kadota, T. Distribution of 5-HT (Serotonin) Immunoreactivity in the Central Nervous System of the Inshore Hagfish, Eptatretus Burgeri. Cell Tissue Res. 1991, 266, 107–116. [Google Scholar] [CrossRef]
- Bernier, N.; Perry, S. Control of Catecholamine and Serotonin Release from the Chromaffin Tissue of the Atlantic Hagfish. J. Exp. Biol. 1996, 199, 2485–2497. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.T.; Nebo, C.; Paula Naves, L.; Fortes-Silva, R.; Regina Cardoso de Oliveira, I.; Paulino, R.R.; Drummond, C.D.; Rosa, P.V. Distribution of Goblet and Endocrine Cells in the Intestine: A Comparative Study in Amazonian Freshwater Tambaqui and Hybrid Catfish. J. Morphol. 2020, 281, 55–67. [Google Scholar] [CrossRef]
- Gustafsson, J.K.; Davis, J.E.; Rappai, T.; McDonald, K.G.; Kulkarni, D.H.; Knoop, K.A.; Hogan, S.P.; Fitzpatrick, J.A.; Lencer, W.I.; Newberry, R.D. Intestinal Goblet Cells Sample and Deliver Lumenal Antigens by Regulated Endocytic Uptake and Transcytosis. eLife 2021, 10, e67292. [Google Scholar] [CrossRef]
- Ridler, C. Sentinel Goblet Cells Flush out Bacteria from Crypts. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 438. [Google Scholar] [CrossRef]
- López Nadal, A.; Ikeda-Ohtsubo, W.; Sipkema, D.; Peggs, D.; McGurk, C.; Forlenza, M.; Wiegertjes, G.F.; Brugman, S. Feed, Microbiota, and Gut Immunity: Using the Zebrafish Model to Understand Fish Health. Front. Immunol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Koch, B.E.V.; Yang, S.; Lamers, G.; Stougaard, J.; Spaink, H.P. Intestinal Microbiome Adjusts the Innate Immune Setpoint during Colonization through Negative Regulation of MyD88. Nat. Commun. 2018, 9, 4099. [Google Scholar] [CrossRef]
- Williams, D.A.; Flood, M.H.; Lewis, D.A.; Miller, V.M.; Krause, W.J. Plasma Levels of Nitrite and Nitrate in Early and Recent Classes of Fish. Comp. Med. 2008, 58, 431–439. [Google Scholar] [PubMed]
- Torrecillas, S.; Caballero, M.J.; Mompel, D.; Montero, D.; Zamorano, M.J.; Robaina, L.; Rivero-Ramírez, F.; Karalazos, V.; Kaushik, S.; Izquierdo, M. Disease Resistance and Response against Vibrio Anguillarum Intestinal Infection in European Seabass (Dicentrarchus Labrax) Fed Low Fish Meal and Fish Oil Diets. Fish Shellfish Immunol. 2017, 67, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Pederzoli, A.; Conte, A.; Tagliazucchi, D.; Gambarelli, A.; Mola, L. Occurrence of Two NOS Isoforms in the Developing Gut of Sea Bass Dicentrarchus Labrax (L.). Histol. Histopathol. 2007, 22, 10. [Google Scholar]
- Rakers, S.; Niklasson, L.; Steinhagen, D.; Kruse, C.; Schauber, J.; Sundell, K.; Paus, R. Antimicrobial Peptides (AMPs) from Fish Epidermis: Perspectives for Investigative Dermatology. J. Investig. Dermatol. 2013, 133, 1140–1149. [Google Scholar] [CrossRef]
- Raju, S.V.; Sarkar, P.; Kumar, P.; Arockiaraj, J. Piscidin, Fish Antimicrobial Peptide: Structure, Classification, Properties, Mechanism, Gene Regulation and Therapeutical Importance. Int. J. Pept. Res. Ther. 2021, 27, 91–107. [Google Scholar] [CrossRef]
- Katzenback, B. Antimicrobial Peptides as Mediators of Innate Immunity in Teleosts. Biology 2015, 4, 607–639. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Lombardo, B.M.; Longhitano, L.; Li Volti, G.; Tibullo, D. Skin Mucus of Marine Fish as a Source for the Development of Antimicrobial Agents. Front. Mar. Sci. 2020, 760, 541853. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antibody | Supplier | Dilution | Animal Source |
|---|---|---|---|
| 5-HT | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | 1:50 | Mouse |
| TLR2 | Active Motif, La Hulpe, Belgium, Europe | 1:125 | Rabbit |
| iNOS | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | 1:200 | Mouse |
| Piscidin1 | GenScript Biotech Corporation, Rijswijk, Netherlands, Europe. Produced on demand | 1:50 | Rabbit |
| Alexa Fluor 488 donkey anti-mouse IgG FITC conjugated | Molecular Probes, Invitrogen | 1:300 | Donkey |
| Alexa Fluor 594 donkey anti-rabbit IgG TRITC conjugated | Molecular Probes, Invitrogen | 1:300 | Donkey |
| No. of Goblet Cells 1 | |
|---|---|
| 5-HT+ | 356.71 ± 24.54 * |
| TLR2+ | 294.29 ± 19.02 * |
| 5-HT + TLR2 | 263.43 ± 28.68 * |
| iNOS+ | 325.02 ± 24.94 ** |
| Piscidin1+ | 385.38 ± 28.05 * |
| iNOS+Piscidin1 | 308.39 ± 15.14 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alesci, A.; Pergolizzi, S.; Savoca, S.; Fumia, A.; Mangano, A.; Albano, M.; Messina, E.; Aragona, M.; Lo Cascio, P.; Capillo, G.; et al. Detecting Intestinal Goblet Cells of the Broadgilled Hagfish Eptatretus cirrhatus (Forster, 1801): A Confocal Microscopy Evaluation. Biology 2022, 11, 1366. https://doi.org/10.3390/biology11091366
Alesci A, Pergolizzi S, Savoca S, Fumia A, Mangano A, Albano M, Messina E, Aragona M, Lo Cascio P, Capillo G, et al. Detecting Intestinal Goblet Cells of the Broadgilled Hagfish Eptatretus cirrhatus (Forster, 1801): A Confocal Microscopy Evaluation. Biology. 2022; 11(9):1366. https://doi.org/10.3390/biology11091366
Chicago/Turabian StyleAlesci, Alessio, Simona Pergolizzi, Serena Savoca, Angelo Fumia, Angelica Mangano, Marco Albano, Emmanuele Messina, Marialuisa Aragona, Patrizia Lo Cascio, Gioele Capillo, and et al. 2022. "Detecting Intestinal Goblet Cells of the Broadgilled Hagfish Eptatretus cirrhatus (Forster, 1801): A Confocal Microscopy Evaluation" Biology 11, no. 9: 1366. https://doi.org/10.3390/biology11091366