
Virulence Characteristics and Emerging Therapies for Biofilm-Forming Acinetobacter baumannii: A Review
 , , ,
, , ,  , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
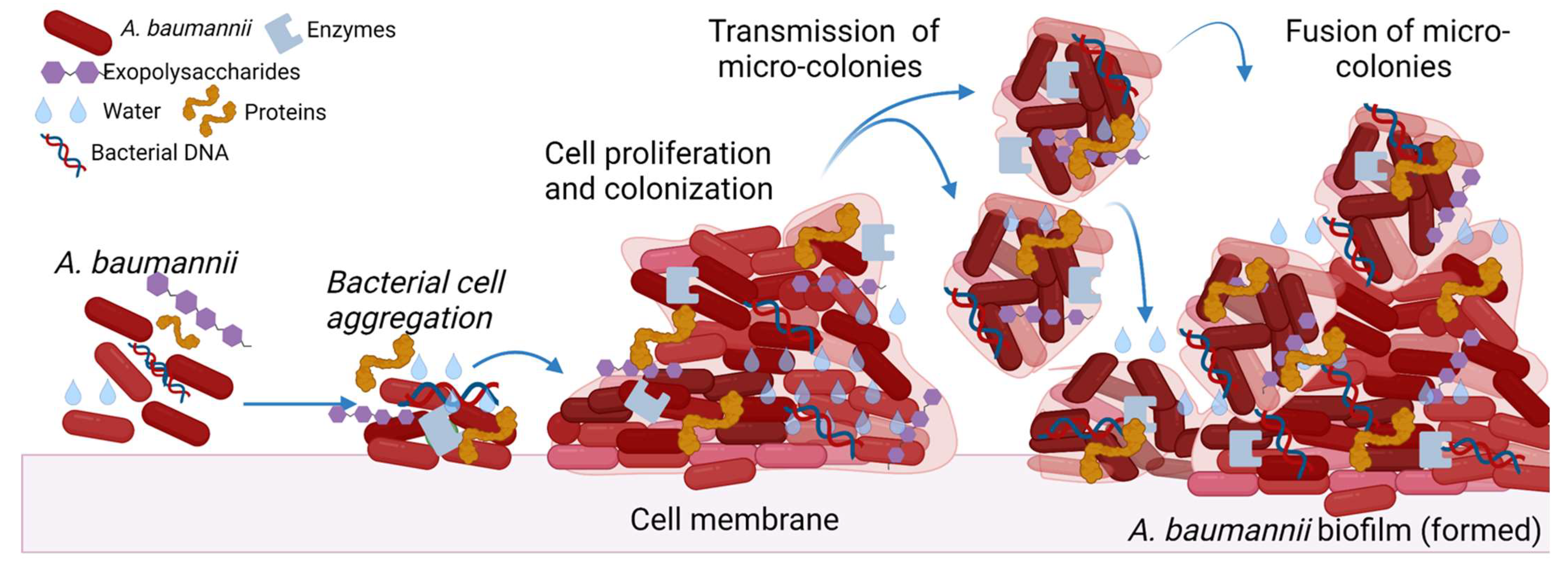
2. Factors Involved in Biofilm Formation
2.1. Physiological Factors
2.2. Type and Nutrient Availability
2.3. Growth Surface
2.4. Iron Concentration
2.5. Expression of the Gene Involved in Biofilms
2.6. Virulence Factors Associated with Biofilms
2.7. Outer Membrane Proteins
2.8. Biofilm-Associated Protein
2.9. Chaperon-Usher Pilus Assembly System of Pili (Csu BABCDE)
2.10. Extracellular Polysaccharides (EPS)
2.11. Quorum Sensing (QS)
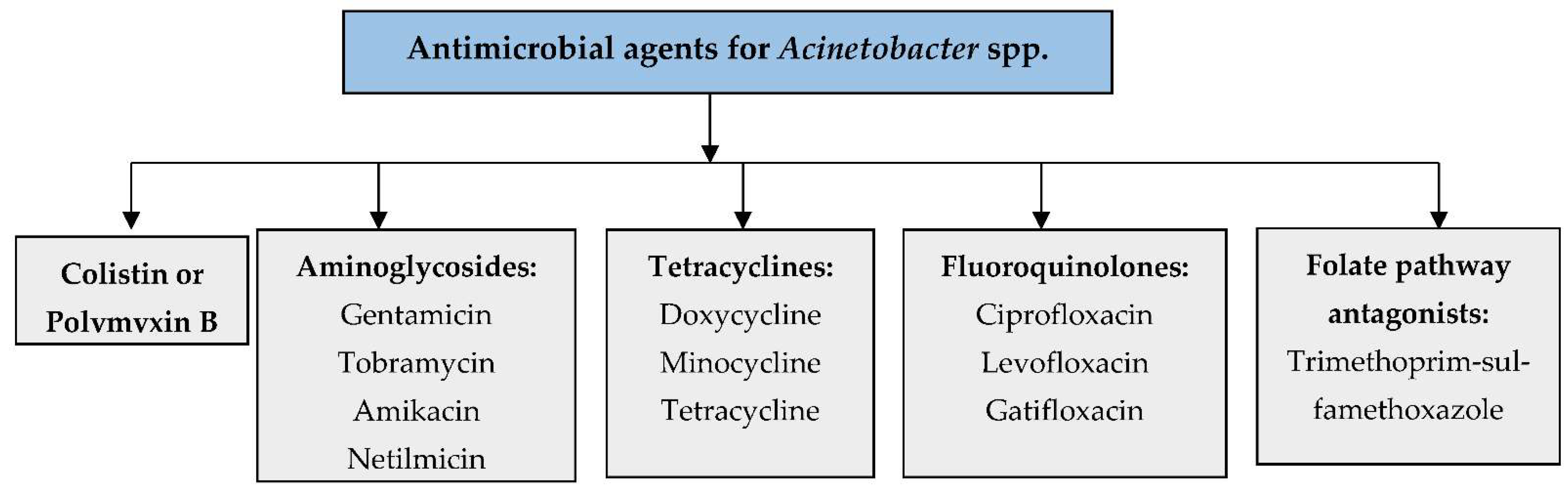
3. Current Antimicrobials for the Treatment of Acinetobacter Infections
Combination Therapy
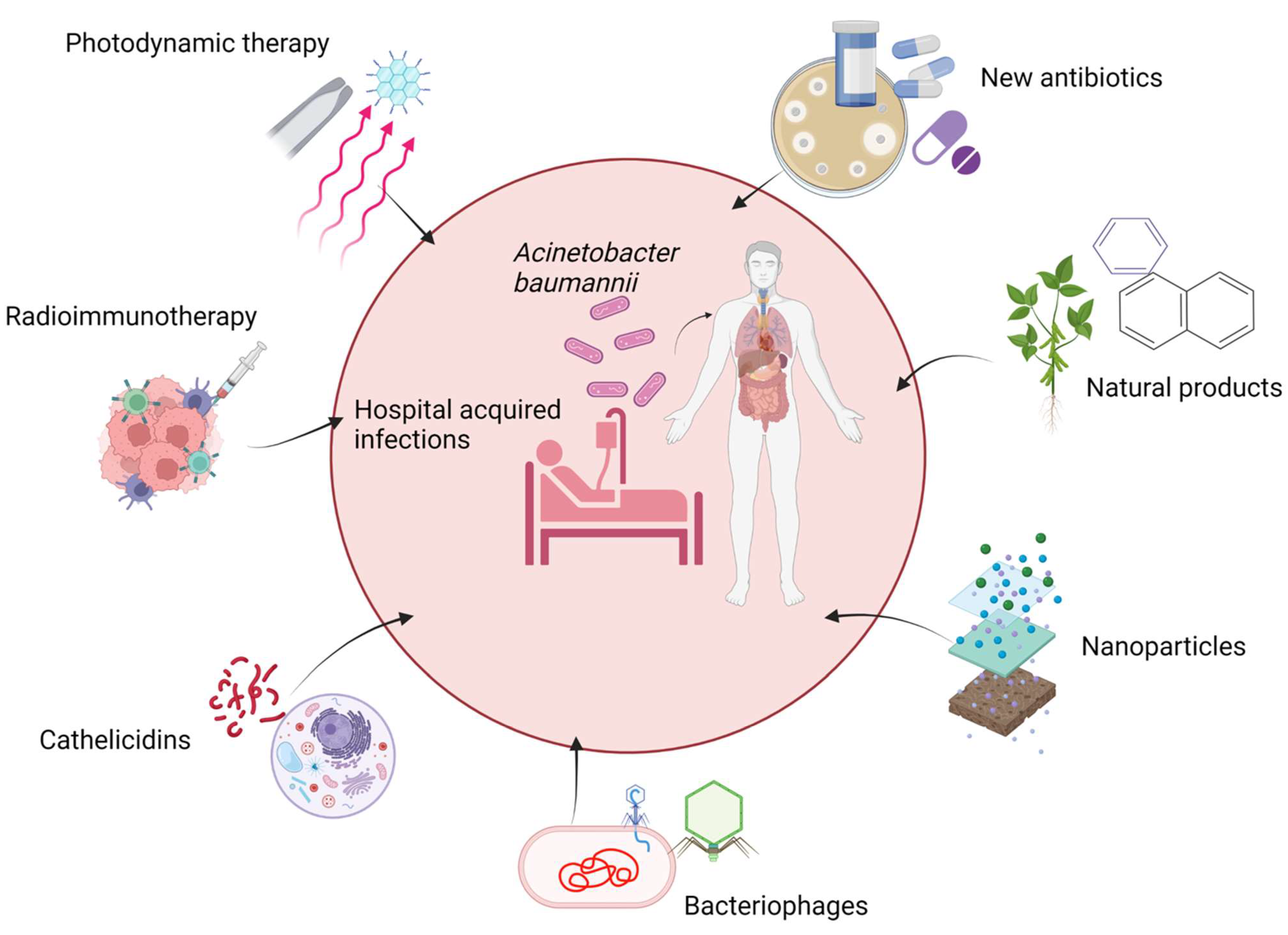
4. Future Therapies for the Treatment of Acinetobacter Infections
4.1. New Antibiotics
4.2. Natural Products
4.3. Nanoparticle Formulation
4.4. Bacteriophage and Bactericidal Gene Transfer Therapy
4.5. Other Products
4.6. Cathelicidins
4.7. Radioimmunotherapy
4.8. Photodynamic Therapy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Duarte, A.; Ferreira, S.; Almeida, S.; Domingues, F.C. Clinical isolates of Acinetobacter baumannii from a Portuguese hospital: PFGE characterization, antibiotic susceptibility and biofilm-forming ability. Comp. Immunol. Microbiol. Infect. Dis. 2016, 45, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Gaddy, J.A.; Tomaras, A.P.; Actis, L.A. Acinetobacter baumannii outer membrane protein A is involved in attachment to biotic and abiotic surfaces and killing of Candida albicans filaments. In Proceedings of the 108th American Society for Microbiology General Meeting, Boston, MA, USA, 1–5 June 2008. [Google Scholar]
- Abdi-Ali, A.; Hendiani, S.; Mohammadi, P.; Gharavi, S. Assessment of biofilm formation and resistance to imipenem and ciprofloxacin among clinical isolates of Acinetobacter baumannii in Tehran. Jundishapur J. Microbiol. 2014, 7, 8606. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, N.; Sharma, P.; Capalash, N. N-acyl homoserine lactone mediated interspecies interactions between A. baumannii and P. aeruginosa. Biofouling 2012, 28, 813–822. [Google Scholar] [CrossRef]
- Rumbo-Feal, S.; Gómez, M.J.; Gayoso, C.; Álvarez-Fraga, L.; Cabral, M.P.; Aransay, A.M.; Rodríguez-Ezpeleta, N.; Fullaondo, A.; Valle, J.; Tomás, M.; et al. Whole transcriptome analysis of Acinetobacter baumannii assessed by RNA-sequencing reveals different mRNA expression profiles in biofilm compared to planktonic cells. PLoS ONE 2013, 8, e72968. [Google Scholar] [CrossRef]
- Avila-Novoa, M.G.; Solís-Velázquez, O.A.; Rangel-López, D.E.; González-Gómez, J.P.; Guerrero-Medina, P.J.; Gutiérrez-Lomelí, M. Biofilm formation and detection of fluoroquinolone- and carbapenem-resistant genes in multidrug-resistan Acinetobacter baumannii. Can. J. Infect. Dis. Med. Microbiol. 2019, 2019, 3454907. [Google Scholar] [CrossRef]
- Monem, S.; Furmanek-Blaszk, B.; Lupkowska, A.; Kuczynska-Wisnik, D.; Stojowska-Swedrzynska, K.; Laskowska, E. Mechanisms protecting Acinetobacter baumannii against multiple stresses triggered by the host immune response, antibiotics and outside-host environment. Int. J. Mol. Sci. 2020, 21, 5498. [Google Scholar] [CrossRef]
- Assaidi, A.; Ellouali, M.; Latrache, H.; Mabrouki, M.; Hamadi, F.; Timinouni, M.; Zahir, H.; El Mdaghri, N.; Barguigua, A.; Mliji, E.M. Effect of temperature and plumbing materials on biofilm formation by Legionella pneumophila serogroup 1 and 2–15. J. Adhes Sci. Technol. 2018, 13, 1–14. [Google Scholar]
- Martí, S.; Rodríguez-Baño, J.; Catel-Ferreira, M.; Jouenne, T.; Vila, J.; Seifert, H.; Dé, E. Biofilm formation at the solid-liquid and air-liquid interfaces by Acinetobacter species. BMC Res. Notes 2011, 4, 5. [Google Scholar] [CrossRef]
- Pour, N.K.; Dusane, D.H.; Dhakephalkar, P.K.; Zamin, F.R.; Zinjarde, S.S.; Chopade, B.A. Biofilm formation by Acinetobacter baumannii strains isolated from urinary tract infection and urinary catheters. FEMS Immunol. Med. Microbiol. 2011, 62, 328–338. [Google Scholar] [CrossRef]
- Wei, X.; Shen, D.; Luo, Y. Molecular mechanism of biofilm formation in Acinetobacter baumannii. Chin. J. Nosocomiol. 2010, 18, 2735–2738. [Google Scholar]
- Dong, R.; Guan, C.; Hu, D.; Xin, T.T.; Qu, Y. The correlation study on antimicrobial resistance and biofilm-related genes in clinical isolates of Acinetobacter baumannii. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2013, 25, 493–494. [Google Scholar] [PubMed]
- Jacobs, A.C.; Blanchard, C.E.; Catherman, S. Anribonuclease T2 family protein modulates Acinetobacter baumannii. Abiotic Surf. Colonization. 2014, 9, e85729. [Google Scholar]
- Wright, M.S.; Jacobs, M.R.; Bonomo, R.A.; Adams, M.D. Transcriptome remodeling of Acinetobacter baumannii during infection and treatment. MBio 2017, 8, e02193-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumbo, C.; Gato, E.; López, M.; Ruiz de Alegría, C.; Fernández-Cuenca, F.; Martínez-Martínez, L.; Vila, J.; Pachón, J.; Cisneros, J.M.; Rodríguez-Baño, J.; et al. The contribution of efflux pumps, porins and β-lactamases to multidrugresistance in clinical isolates of Acinetobacter baumannii. Antimicrob Agents Chemother 2013, 57, 5247–5257. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, C.H.; Yarhui, N.B.; Nastro, M.; Quezada, T.N.; Cañarte, G.C.; Ventura, R.M.; Cuba, T.U.; Valenzuela, N.; Roach, F.; Mota, M.I.; et al. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii in South America. J. Med. Microbiol. 2016, 65, 1088–1091. [Google Scholar] [CrossRef]
- Badave, G.K. Biofilm producing multidrug resistant Acinetobacter baumannii: An emerging challenge. J. Clin. Diagn. Res. 2015, 9, DC08-10. [Google Scholar] [CrossRef]
- Cao, Q.; Wang, Y.; Chen, F.; Xia, Y.; Lou, J.; Zhang, X.; Yang, N.; Sun, X.; Zhang, Q.; Zhuo, C.; et al. A novel signal transduction pathway that modulates rhl quorum sensing and bacterial virulence in Pseudomonas aeruginosa. PLOS Pathog. 2014, 10, e1004340. [Google Scholar] [CrossRef]
- Russo, T.A.; MacDonald, U.; Beanan, J.M.; Olson, R.; MacDonald, I.J.; Sauberan, S.L.; Luke, N.R.; Wayne Schultz, L.; Umland, T.C. Penicillin-binding protein 7/8 contributes to the survival of Acinetobacter baumanniiin in vitro and in vivo. J. Infect Immun. 2009, 199, 513–521. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, M.S.; Kim, S.; Son, J.H.; Kim, S.; Lee, Y.C.; Shin, M.; Oh, M.H.; Lee, J.C. The sensor kinase BfmS controls production of outer membrane vesicles in Acinetobacter baumannii. BMC Microbiol. 2019, 19, 301. [Google Scholar] [CrossRef]
- Aly, M.M.; Abu Alsoud, N.M.; Elrobh, M.S.; Al Johani, S.M.; Balkhy., H.H. High prevalence of the PER-1 gene among carbapenem-resistant Acinetobacter baumannii in Riyadh, Saudi Arabia. Eur. J. Clin. Microbiol. Infect Dis. 2016, 35, 1759–1766. [Google Scholar] [CrossRef]
- Ryu, S.Y.; Baek, W.K.; Kim, H.A. Association of biofilm production with colonization among clinical isolates of Acinetobater baumannii. Korean J. Intern. Med. 2017, 32, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Dahdouh, E.; Gómez-Gil, R.; Pacho, S.; Mingorance, J.; Daoud, Z.; Suárez, M. Clonality, virulence determinants, and profiles of resistance of clinical Acinetobacter baumannii isolates obtained from a Spanish hospital. PLoS ONE 2017, 12, e0176824. [Google Scholar] [CrossRef] [PubMed]
- Chapartegui-González, I.; lázaro-Díez, M.; Bravo, Z.; Navas, J.; Icardo, J.M.; Ramos-Vivas, J. Acinetobacter baumannii maintains its virulence after long-time starvation. PLoS ONE 2018, 13, e0201961. [Google Scholar] [CrossRef]
- Choi, C.H.; Lee, J.S.; Lee, Y.C.; Park, T.I.; Lee, J.C. Acinetobacter baumannii invades epithelial cells and outer membrane protein A mediates interactions with epithelial cells. BMC Microbiol. 2008, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Smani, Y.; Fàbrega, A.; Roca, I.; Sánchez-Encinales, V.; Vila, J.; Pachón, J. Role of OmpA in the multidrug resistance phenotype of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2014, 58, 1806–1808. [Google Scholar] [CrossRef]
- Gaddy, J.A.; Actis, L.A. Regulation of Acinetobacter baumannii biofilm formation. Futur. Microbiol. 2009, 4, 273–278. [Google Scholar] [CrossRef]
- Fattahian, Y.; Rasooli, I.; Gargari, S.L.M.; Rahbar, M.R.; Astaneh, S.D.A.; Amani, J. Protection against Acinetobacter baumannii infection via its functional deprivation of biofilm-associated protein (Bap). Microb. Pathog. 2011, 51, 402–406. [Google Scholar] [CrossRef]
- Ghasemi, E.; Ghalavand, Z.; Goudarzi, H.; Yeganeh, F.; Hashemi, A.; Dabiri, H.; Mirsamadi, E.S.; Foroumand, M. Phenotypic and genotypic investigation of biofilm formation in clinical and environmental isolates of Acinetobacter baumannii. Arch. Clin. Infect. Dis. 2018, 13, 12914. [Google Scholar] [CrossRef]
- Loehfelm, T.W.; Luke, N.R.; Campagnari, A.A. Identification and characterization of an Acinetobacter baumannii biofilm associated protein. J. Bacteriol. 2008, 190, 1036–1044. [Google Scholar] [CrossRef]
- Bodelon, G.; Palomino, C.; Fernandez, L.A. Immunoglobulin domains in Escherichia coli and other enterobacteria: From pathogenesis to applications in antibody technologies. FEMS Microbiol. Rev. 2013, 37, 204–250. [Google Scholar] [CrossRef]
- Hazenbos, W.L.W.; Kajihara, K.K.; Vandlen, R.; Morisaki, J.H.; Lehar, S.M.; Kwakkenbos, M.J.; Beaumont, T.; Bakker, A.Q.; Phung, Q.; Swem, L.R.; et al. Novel staphylococcal glyco-syltransferases SdgA and SdgB mediate immunogenicity and protection of virulence-associated cell wall proteins. PLoS Pathog. 2013, 9, e1003653. [Google Scholar] [CrossRef] [PubMed]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii biofilm-associated protein plays a role in adherence to human epithelial cells. Infect. Immun. 2012, 80, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Lasa, I.; Penadés, J.R. Bap: A family of surface proteins involved in biofilm formation. Res. Microbiol. 2006, 157, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, A.P.; Dorsey, C.W.; Edelmann, R.E.; Actis, L.A. Attachment to and biofilm formation on abiotic surfaces by Acineto-bacter baumannii: Involvement of a novel chaperone-usher pili assembly system. Microbiology 2003, 149, 3473–3484. [Google Scholar] [CrossRef]
- Pakharukova, N.; Tuittila, M.; Paavilainen, S.; Malmi, H.; Parilova, O.; Teneberg, S.; Knight, S.D.; Zavialov, A.V. Structural basis for Acinetobacter baumannii biofilm formation. Proc. Natl. Acad. Sci. USA 2018, 115, 5558–5563. [Google Scholar] [CrossRef]
- Pakharukova, N.; Garnett, J.A.; Tuittila, M.; Paavilainen, S.; Diallo, M.; Xu, Y.; Matthews, S.J.; Zavialov, A.V. Structural insight into archaic and alternative chaperone usher pathways reveals a novel mechanism of pilus biogenesis. PLoS Pathog. 2015, 11, e1005269.8. [Google Scholar] [CrossRef]
- Nuccio, S.-P.; Baumler, A.J. Evolution of the chaperone/usher assembly pathway: Fimbrial classification goes greek. Microbiol. Mol. Biol. Rev. 2007, 71, 551–575. [Google Scholar] [CrossRef]
- Choudhury, D.; Thompson, A.; Stojanoff, V.; Langermann, S.; Pinkner, J.; Hultgren, S.J.; Knight, S.D. X-ray structure of the FimC-FimH chaperone-adhesin complex from uropathogenic Escherichia coli. Science 1999, 285, 1061–1066. [Google Scholar] [CrossRef]
- Morgan, D.J.; Liang, S.Y.; Smith, C.L.; Kristie Johnson, J.; Harris, A.D.; Furuno, J.P.; Thom, K.A.; Snyder, G.M.; Day, H.R.; Perencevich, E.N. Frequent multidrug-resistant Acinetobacter baumannii contamination of gloves, gowns, and hands of healthcare workers. Infect. Control. Hosp. Epidemiol. 2010, 31, 716–721. [Google Scholar] [CrossRef]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Genet. 2017, 16, 91–102. [Google Scholar] [CrossRef]
- Lewis, K. Multidrug tolerance of biofilms and persister cells. In Current Topics in Microbiology and Immunology; Romeo, T., Ed.; Springer: Berlin, Heidelberg, 2008; Volume 322, pp. 107–131. [Google Scholar] [CrossRef]
- Cuccui, J.; Wren, B.W. Bacteria like sharing their sweets. Mol. Microbiol. 2013, 89, 811–815. [Google Scholar] [CrossRef] [PubMed]
- Bitrian, M.; Solari, C.M.; González, R.H.; Nudel, C.B. Identification of virulence markers in clinically relevant strains of Acineto-bacter genospecies. Int. Microbiol. 2012, 15, 79–88. [Google Scholar] [PubMed]
- Stacy, D.M.; Welsh, M.A.; Rather, P.N.; Blackwell, H.E. Attenuation of quorum sensing in the pathogen Acinetobacter baumannii using non-native N-Acyl homoserine lactones. ACS Chem. Biol. 2012, 7, 1719–1728. [Google Scholar] [CrossRef]
- Liou, M.L.; Soo, P.C.; Ling, S.R.; Kuo, H.Y.; Tang, C.Y.; Chang, K.C. The sensor kinase BfmS mediates virulence in Acinetobacter baumannii. J. Microbiol. Immunol. Infect. 2014, 47, 275–281. [Google Scholar] [CrossRef]
- Kim, I.H.; Wen, Y.; Son, J.S.; Lee, K.H.; Kim, K.S. The fur-iron complex modulates expression of the quorum-sensing master regulator, SmcR, to control expression of virulence factors in Vibrio vulnificus. Infect. Immun. 2013, 81, 2888–2898. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed]
- Maragakis, L.L.; Perl, T.M. Acinetobacter baumannii: Epidemiology, antimicrobial resistance, and treatment options. Clin. Infect. Dis. 2008, 46, 1254–1263. [Google Scholar] [CrossRef]
- Emmanuel, C.E.; Hafizah, Y.C.; Mohamed, E.Z. Acinetobacter baumannii biofilms: Effects of physico-chemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect. Drug Resist. 2018, 11, 2277–2299. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI supplement M100 (ISBN 978-1-68440-134-5 [Print]; ISBN 978-1-68440-135-2 [Electronic]); Clinical and Laboratory Standards Institute: Wayne, PN, USA, 2022. [Google Scholar]
- Petrosillo, N.; Ioannidou, E.; Falagas, M. Colistin monotherapy vs. combination therapy: Evidence from microbiological, animal and clinical studies. Clin. Microbiol. Infect. 2008, 14, 816–827. [Google Scholar] [CrossRef]
- Bassetti, M.; Poulakou, G.; Giamarellou, H. Is there a future for tigecycline? Intensiv. Care Med. 2014, 40, 1039–1045. [Google Scholar] [CrossRef]
- Sader, H.S.; Castanheira, M.; Flamm, R.K.; Mendes, R.E.; Farrell, D.J.; Jones, R.N. Ceftazidime/avibactam tested against Gram-negative bacteria from intensive care unit (ICU) and non-ICU patients, including those with ventilator-associated pneumonia. Int. J. Antimicrob. Agents 2015, 46, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Dijkshoorn, L.; Aucken, H.; Gerner-Smidt, P.; Janssen, P.; Kaufmann, M.E.; Garaizar, J.; Ursing, J.; Pitt, T.L. Comparison of outbreak and non-outbreak Acinetobacter baumannii strains by genotypic and phenotypic methods. J. Clin. Microbiol. 1996, 34, 1519–1525. [Google Scholar] [CrossRef]
- Song, J.Y.; Cheong, H.J.; Noh, J.Y.; Kim, W.J. In vitro comparison of antibiofilm effects against carbapenem-resistant Acineto-bacter baumannii: Imipenem, colistin, tigecycline, rifampicin and combinations. Infect. Chemother. 2015, 47, 27–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLeod, S.M.; Moussa, S.H.; Hackel, M.A.; Miller, A.A. In vitro activity of sulbactam-durlobactam against Acinetobacter baumannii- calcoaceticus complex isolates collected globally in 2016 and 2017. Antimicrob. Agents Chemother. 2020, 64, e02534-19. [Google Scholar] [CrossRef] [PubMed]
- Rhomberg, P.R.; Shortridge, D.; Huband, M.D.; Butler, D.; West, J.; Flamm, R.K. Multilaboratory broth microdilution MIC reproducibility study for GSK3342830, a novel catechol-cephem, abstr SATURDAY287. In Proceedings of the ASM Microbe, New Orleans, LA, USA, 1–5 June 2017; GlaxoSmithKline: Brentford, UK, 2017. [Google Scholar]
- Geom Therapeutics. Geom Therapeutics Announces Agreement with National Institute of Allergy and Infectious Diseases (NIAID) for Clinical Advancement of GT-1; Geom Therapeutics: San Francisco, CA, USA, 2017. [Google Scholar]
- Ito, A.; Sato, T.; Ota, M.; Takemura, M.; Nishikawa, T.; Toba, S.; Kohira, N.; Miyagawa, S.; Ishibashi, N.; Matsumoto, S.; et al. In vitro antibacterial properties of cefiderocol, a novel siderophore cephalosporin, against Gram-negative bacteria. Antimicrob. Agents Chemother. 2017, 23, e01454-17. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, M.; Kazmierczak, K.M.; Hackel, M.; Echols, R.; Yamano, Y.; Sahm, D.F. Cefiderocol (S-649266) susceptibility against globally isolated meropenem non-susceptible Gram-negative bacteria containing serine- and metal-lo-carbapenemase genes. abstr SUNDAY-25. In Proceedings of the ASM Microbe, New Orleans, LA, USA, 1–5 June 2017. [Google Scholar]
- Ghosh, M.; Miller, P.A.; Möllmann, U.; Claypool, W.D.; Schroeder, V.A.; Wolter, W.R.; Suckow, M.; Yu, H.; Li, S.; Huang, W.; et al. Targeted antibiotic delivery: Selective siderophore conjugation with daptomycin confers potent activity against multidrug resistant Acinetobacter baumannii both in Vitro and in vivo. J. Med. Chem. 2017, 60, 4577–4583. [Google Scholar] [CrossRef]
- Entasis Therapeutics. Evaluation of Safety and Efficacy of Intravenous Sulbactam-ETX2514 in the Treatment of Hospitalized Adults with Complicated Urinary Tract Infections. Accession no. NCT03445195. ClinicalTrials.gov, NIH. 2018. Available online: https://clinicaltrials.gov/ct2/show/NCT03445195?termNCT03445195&rank1.LA (accessed on 17 August 2022).
- Hackel, M.; Bouchillon, S.; deJonge, B.; Lawrence, K.; Mueller, J.; Tommasi, R.; Miller, A. Global Surveillance of the Activity of Sulbactam Combined with the Novel-Lactamase Inhibitor ETX2514 against Clinical Isolates of Acinetobacter baumannii from 2014; Entasis Therapeutics: Waltham, MA, USA, 2017. [Google Scholar]
- Evaluation of the Pharmacokinetics, Safety, and Tolerability of Intravenous ETX2514 and Sulbactam Administered Concurrently to Subjects with Various Degrees of Renal Impairment and Healthy Matched Control Subjects. Accession no. NCT03310463. ClinicalTrials.gov, NIH. Available online: https://clinicaltrials.gov/ct2/show/NCT03310463term-NCT03310463&rank1 (accessed on 17 August 2022).
- Spero Therapeutics Unveils Data on Lead Potentiator Candidate for the Treatment of Multidrug-Resistant Gram-Negative Infections at ASM Microbe 2016; Spero Therapeutics: Cambridge, MA, USA, 2016.
- Papp-Wallace, K.M.; Nguyen, N.Q.; Jacobs, M.R.; Bethel, C.R.; Barnes, M.D.; Kumar, V.; Bajaksouzian, S.; Rudin, S.D.; Rather, P.N.; Bhavsar, S.; et al. Strategic approaches to overcome resistance against gram-negative pathogens using β-lactamase inhibitors and β-lactam enhancers: Activity of three novel diazabicyclooctanes WCK 5153, Zidebactam (WCK 5107), and WCK 4234. J. Med. Chem. 2018, 61, 4067–4086. [Google Scholar] [CrossRef]
- Castanheira, M.; Rhomberg, P.R.; Lindley, J.M.; Jones, R.N.; Sader, H.S. Activity of the new carbapenem/-lactamase inhibitor combination WCK 5999 against Gram-negative isolates producing oxacillinases (OXAs), abstr MON-DAY-422. In Proceedings of the ASM Microbe, Boston, MA, USA, 16–20 June 2016. [Google Scholar]
- Vázquez-Ucha, J.C.; Maneiro, M.; Martínez-Guitián, M.; Buynak, J.; Bethel, C.R.; Bonomo, R.A.; Bou, G.; Poza, M.; González-Bello, C.; Beceiro, A. Activity of the β-lactamase inhibitor LN-1-255 against carbapenem-hydrolyzing class D β-lactamases from Acinetobacter baumannii. Antimicrob. Agents Chemother. 2017, 61, e01172-17. [Google Scholar] [CrossRef]
- Joo, H.Y.; Kim, D.I.; Kowalik, E.; Li, Y.; Mao, S.; Liu, S.; Hager, M.W.; Choi, W.B. FSI-1671, a novel anti-Acinetobacter carbapenem; in vitro activities of FSI-1671 and FSI-1671/sulbactam against MDR-A. baumannii, abstr F-1202. In Proceedings of the 53rd Annual Interscience Conference on Antimicrobial Agents and Chemotherapy, Denver, CO, USA, 10–13 September 2013. [Google Scholar]
- Kang, A.D.; Smith, K.P.; Eliopoulos, G.M.; Berg, A.H.; McCoy, C.; Kirby, J.E. In vitro Apramycin Activity against multidrug-resistant Acinetobacter baumannii and Pseudomonas aeruginosa. Diagn. Microbiol. Infect. Dis. 2017, 88, 188–191. [Google Scholar] [CrossRef]
- Vickers, A.; Mushtaq, S.; Woodford, N.; Doumith, M.; David, M. Activity of RX-04 Pyrrolocytosine protein synthesis inhibitors against multidrug-resistant Gram-negative bacteria. Antimicrob. Agents Chemother. 2018, 62, e00689-18. [Google Scholar] [CrossRef]
- Tetra Phase Pharmaceuticals. Tetra Phase Pharmaceuticals Provides Update on Eravacycline Regulatory and Development Status; Tetraphase Pharmaceuticals: Watertown, MA, USA, 2016. [Google Scholar]
- Seifert, H.; Stefanik, D.; Sutcliffe, J.A.; Higgins, P.G. In-vitro activity of the novel fluorocycline eravacycline against carbapenem non-susceptible Acinetobacter baumannii. Int. J. Antimicrob. Agents 2018, 51, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Skalweit, M.J.; Li, M. Bulgecin A as a β-lactam enhancer for carbapenem-resistant Pseudomonas aeruginosa and carbapenem-resistant Acinetobacter baumannii clinical isolates containing various resistance mechanisms. Drug Des. Dev. Ther. 2016, 10, 3013–3020. [Google Scholar] [CrossRef]
- Livermore, D.M.; Mushtaq, S.; Warner, M.; Woodford, N. In-vitro activity of eravacycline against carbapenem-resistant Enterobacteriaceae and Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 4, 3840–3844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, M.; Olafisoye, O.; Cortes, C.; Urban, C.; Landman, D.; Quale, J. Activity of eravacycline against Enterobacteriaceae and Acinetobacter baumannii, including multidrug-resistant isolates, from New York City. Antimicrob. Agents Chemother. 2015, 59, 1802–1805. [Google Scholar] [CrossRef] [PubMed]
- Seifert, H.; Stefanik, D.; Sutcliffe, J.; Higgins, P.G. In-vitro activity of the novel fluorocycline TP-6076 against car-bapenem non-susceptible Acinetobacter baumannii, abstr P-1364. In Proceedings of the 27th Europe Cong Clinical Microbiology Infection Disease, Vienna, Austria, 22–25 April 2017. [Google Scholar]
- Lemaitre, N.; Liang, X.; Najeeb, J.; Lee, C.J.; Titecat, M.; Leteurtre, E.; Simonet, M.; Toone, E.J.; Zhou, P.; Sebbane, F. Curative treatment of severe Gram-negative bacterial infections by a new class of antibiotics targeting LpxC. MBio 2017, 8, e00674-17. [Google Scholar] [CrossRef] [PubMed]
- Flamm, R.K.; Rhomberg, P.R.; Jones, R.N.; Farrell, D.J. In vitro activity of RX-P873 against Enterobacteriaceae, Pseudomonas aeruginosa, and Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 2280–2285. [Google Scholar] [CrossRef] [PubMed]
- Guclu, E.; Genc, H.; Zengin, M.; Karabay, O. Antibacterial activity of Lythrum salicaria against multidrug-resistant Acinetobacter baumannii and Pseudomonas aeruginosa. Annu. Res. Rev. Biol. 2014, 4, 1099–1105. [Google Scholar] [CrossRef]
- Miyasaki, Y.; Rabenstein, J.D.; Rhea, J.; Crouch, M.-L.; Mocek, U.M.; Kittell, P.E.; Morgan, M.A.; Nichols, W.S.; Van Benschoten, M.M.; Hardy, W.D.; et al. Isolation and characterization of antimicrobial compounds in plant extracts against multidrug-resistant Acinetobacter baumannii. PLoS ONE 2013, 8, e61594. [Google Scholar] [CrossRef]
- Montagu, A.; Saulnier, P.; Cassisa, V.; Rossines, E.; Eveillard, M.; Joly-Guillou, M.L. Aromatic and terpenic compounds loaded in lipidic nanocapsules: Activity against multi-drug resistant Acinetobacter baumannii assessed in vitro and in a murine model of sepsis. J. Nanomed. Nanotechnol. 2014, 5, 1000206. [Google Scholar] [CrossRef]
- Pelletier, R.P. Effect of Plant-Derived Molecules on Acinetobacter baumannii Biofilm on Abiotic Surfaces. Ph.D. Thesis, University of Connecticut, Storrs, CT, USA, 2012. [Google Scholar]
- Betts, J.W.; Wareham, D.W. In vitro activity of curcuminin combination with Epigallocatechingallate (EGCG) versus multidrug-resistant Acinetobacter baumannii. BMC Microbiol. 2014, 14, 172. [Google Scholar] [CrossRef]
- Betts, J.W.; Kelly, S.M.; Haswell, S.J. Antibacterial effects of theaflavin and synergy with epicatechin against clinical isolates of Acinetobacter baumannii and Stenotrophomonas maltophilia. Int. J. Antimicrob. Agents. 2011, 38, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.C.L.; Lau, C.B.S.; Jolivalt, C.; Lui, S.L.; Ganem-Elbaz, C.; Paris, J.M. Chinese medicinal herbs against an-tibiotic-resistant bacterial pathogens. In Science against Microbial Pathogens: Communicating Current Research Technology Advance; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2011; Volume 2, pp. 73–781. [Google Scholar]
- Osterburg, J.A.; Gardner, S.; Hyon, H.; Neely, A.; Babcock, G. Highly antibiotic-resistant Acinetobacter baumannii clinical isolates are killed by the green tea polyphenol (–)-epigallocatechin-3-gallate (EGCG). Clin. Microbiol. Infect. 2009, 15, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.N.; Stavrinides, J. Pantoea natural product 3 is encoded by an eight-gene biosynthetic gene cluster and exhibits antimicrobial activity against multi-drug resistant Acinetobacter baumannii and Pseudomonas aeruginosa. Microbiol. Res. 2020, 234, 126412. [Google Scholar] [CrossRef] [PubMed]
- Kostoulias, X.; Murray, G.L.; Cerqueira, G.M.; Kong, J.B.; Bantun, F.; Mylonakis, E.; Khoo, C.A.; Peleg, A.Y. Impact of a Cross-Kingdom Signaling Molecule of Candida albicans on Acinetobacter baumannii Physiology. Antimicrob. Agents Chemother. 2016, 60, 161–167. [Google Scholar] [CrossRef]
- Shin, B.; Park, W. Synergistic effect of oleanolic acid on aminoglycoside antibiotics against Acinetobacter baumannii. PLoS ONE 2015, 10, e0137751. [Google Scholar] [CrossRef]
- Mihu, M.R.; Sandkovsky, U.; Han, G.; Friedman, J.M.; Nosanchuk, J.D.; Martinez, L.R. The use of nitric oxide releasing nanoparticles as a treatment against Acinetobacter baumannii in wound infections. Virulence 2010, 1, 62–67. [Google Scholar] [CrossRef]
- Shaker, M.A.; Shaaban, M.I. Synthesis of silver nanoparticles with antimicrobial and anti-adherence activities against multidrug-resistant isolates from Acinetobacter baumannii. J. Taibah Univ. Med. Sci. 2017, 12, 291–297. [Google Scholar] [CrossRef]
- Friedman, A.; Friedman, J. New biomaterials for the sustained release of nitric oxide: Past, present and future. Expert Opin. Drug Deliv. 2009, 6, 1113–1122. [Google Scholar] [CrossRef]
- Hetta, H.F.; Al-Kadmy, M.S.I.; Khazaal, S.S.; Abbas, S.; Suhail, A.; El-Mokhtar, M.A.; Abd Ellah, N.H.; Ahmed, E.A. Antibiofilm and antivirulence potential of silver nanoparticles against multidrug-resistant Acinetobacter baumannii. Sci. Rep. 2021, 11, 10751. [Google Scholar] [CrossRef]
- Wintachai, P.; Paosen, S.; Yupanqui, C.T.; Voravuthikunchai, S.P. Silver nanoparticles synthesized with Eucalyptus critriodora ethanol leaf extract stimulate antibacterial activity against clinically multidrug-resistant Acinetobacter baumannii isolated from pneumonia patients. Microb. Pathog. 2018, 126, 245–257. [Google Scholar] [CrossRef]
- Hemeg, H.A. Nanomaterials for alternative antibacterial therapy. Int. J. Nanomed. 2017, 12, 8211–8225. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yu, X.; Huo, Q.; Yuan, X.Q.; Li, X.; Xu, C.; and Bao, H. Biomedical potentialities of silver nanoparticles for clinical multiple drug-resistant Acinetobacter baumannii. J. Nanomater. 2019, 2019, 3754018. [Google Scholar] [CrossRef]
- Banoub, N.G.; E Saleh, S.; Helal, H.S.; Aboshanab, K.M. Antibiotics combinations and chitosan nanoparticles for combating multidrug resistance Acinetobacter baumannii. Infect. Drug Resist. 2021, 14, 3327–3339. [Google Scholar] [CrossRef] [PubMed]
- Wan, G.; Ruan, L.; Yin, Y.; Yang, T.; Ge, M.; Cheng, X. Effects of silver nanoparticles in combination with antibiotics on the resistant bacteria Acinetobacter baumannii. Int. J. Nanomed. 2016, 11, 3789–3800. [Google Scholar] [CrossRef]
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef]
- Rhoads, D.D.; Wolcott, R.D.; Kuskowski, M.A.; Wolcott, B.M.; Ward, L.S.; Sulakvelidze, A. Bacteriophage therapy of venous leg ulcers in humans: Results of a phase I safety trial. J. Wound Care 2009, 18, 237–243. [Google Scholar] [CrossRef]
- Wright, A.; Hawkins, C.H.; Änggård, E.E.; Harper, D.R. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy. Clin. Otolaryngol. Allied Sci. 2009, 34, 349–357. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii Infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef]
- Yang, H.; Liang, L.; Lin, S.; Jia, S. Isolation and characterization of a virulent bacteriophage AB1 of Acinetobacter baumannii. BMC Microbiol. 2010, 10, 131. [Google Scholar] [CrossRef]
- Wintachai, P.; Voravuthikunchai, S.P. Characterization of novel lytic Myoviridae phage infecting multidrug-resistant Acinetobacter baumannii and synergistic antimicrobial efficacy between phage and Sacha Inchi oil. Pharmaceuticals 2022, 15, 291. [Google Scholar] [CrossRef]
- Mousavi, S.M.; Babakhani, S.; Moradi, L.; Karami, S.; Shahbandeh, M.; Mirshekar, M.; Mohebi, S.; Moghadam, M.T. Bacteriophage as a novel therapeutic weapon for killing colistin-resistant multi-drug-resistant and extensively drug-resistant gram-negative bacteria. Curr. Microbiol. 2021, 78, 4023–4036. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, X.; Shi, Y.; Yin, S.; Shen, W.; Chen, J.; Chen, Y.; Chen, Y.; You, B.; Gong, Y.; et al. Characterization and genome annotation of a newly detected bacteriophage infecting multidrug-resistant Acinetobacter baumannii. Arch. Virol. 2019, 164, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.C.; Lin, N.T.; Hu, A.; Lin, Y.S.; Chen, L.K.; Lai, M.J. Genomic analysis of bacteriophage varphiAB1, a varphiKMV-like virus infecting multidrug-resistant Acinetobacter baumannii. Genomics 2011, 97, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vukotic, G.; Obradovic, M.; Novovic, K.; Di Luca, M.; Jovcic, B.; Fira, D.; Neve, H.; Kojic, M.; McAuliffe, O. Characterization, antibiofilm, and depolymerizing activity of two phages active on carbapenem-resistant Acinetobacter baumannii. Front. Med. 2020, 7, 426. [Google Scholar] [CrossRef]
- Jeon, J.; Park, J.H.; Yong, D. Efficacy of bacteriophage treatment against carbapenem-resistant Acinetobacter baumannii in Galleria mellonella larvae and a mouse model of acute pneumonia. BMC Microbiol. 2019, 19, 70. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Sisakhtpour, B.; Mirzaei, A.; Karbasizadeh, V.; Moghim, S. Efficacy of isolated bacteriophage against biofilm embedded colistin-resistant Acinetobacter baumannii. Gene Rep. 2020, 22, 100984. [Google Scholar] [CrossRef]
- Bernstein, L.R. Mechanisms of therapeutic activity for gallium. Pharmacol. Rev. 1998, 50, 665–682. [Google Scholar]
- Antunes, L.C.S.; Imperi, F.; Minandri, F.; Visca, P. In vitro and in vivo antimicrobial activities of gallium nitrate against multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2012, 56, 5961–5970. [Google Scholar] [CrossRef]
- Arivett, B.A.; Fiester, S.E.; Ohneck, E.J.; Penwell, W.F.; Kaufman, C.M.; Relich, R.F.; Actis, L.A. Antimicrobial activity of gallium protoporphyrin IX AGAINST Acinetobacter baumannii strains displaying different antibiotic resistance phenotypes. Antimicrob. Agents Chemother. 2015, 59, 7657–7665. [Google Scholar] [CrossRef]
- Rumbo, C.; Fernández-Moreira, E.; Merino, M.; Poza, M.; Mendez, J.A.; Soares, N.C.; Mosquera, A.; Chaves, F.; Bou, G. Horizontal transfer of the OXA-24 carbapenemase gene via outer membrane vesicles: A new mechanism of dissemination of carbapenem resistance genes in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 3084–3090. [Google Scholar] [CrossRef]
- Reid, M. The importance of guidelines in the development and application of probiotics. Curr. Pharm. Des. 2005, 11, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Takahashi, A.; Yuki, N.; Kaji, R.; Takahashi, T.; Nomoto, K. Protective effect of a synbiotic against multidrug-resistant Acinetobacter baumannii in a murine infection model. Antimicrob. Agents Chemother. 2016, 60, 3041–3050. [Google Scholar] [CrossRef] [PubMed]
- Parra Millán, R.; Jiménez Mejías, M.E.; Sánchez Encinales, V.; Ayerbe Algaba, R.; Gutiérrez Valencia, A.; Pachón Ibáñez, M.E.; Díaz, C.; Pérez del Palacio, J.; López Cortés, L.F.; Pachón, J.; et al. Efficacy of lysophosphatidylcholine in combination with antimicrobial agents against Acinetobacter baumannii in experimental murine peritoneal sepsis and pneumonia models. Antimicrob. Agents Chemother. 2016, 60, 4464–4470. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Das, S. Mammalian antimicrobial peptides: Promising therapeutic targets against infection and chronic inflammation. Curr. Top. Med. Chem. 2015, 16, 99–129. [Google Scholar] [CrossRef]
- Bahar, A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef]
- Galdiero, S.; Falanga, A.; Berisio, R.; Grieco, P.; Morelli, G.; Galdiero, M. Antimicrobial peptides as an opportunity against bacterial diseases. Curr. Med. Chem. 2015, 22, 1665–1677. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Jang, A.R.; Park, J.Y.; Ahn, J.H.; Lee, T.S.; Kim, D.Y.; Jung, D.H.; Song, E.J.; Hong, J.J.; Park, J.H. Cathelicidin-related antimicrobial peptide contributes to host immune responses against pulmonary infection with Acinetobacter baumannii in mice. Immune Netw. 2020, 20, e25. [Google Scholar] [CrossRef]
- Esfandiyari, R.; Halabian, R.; Behzadi, E.; Sedighian, H.; Jafari, R.; Fooladi, A.A.I. Performance evaluation of antimicrobial peptide ll-37 and hepcidin and β-defensin-2 secreted by mesenchymal stem cells. Heliyon 2019, 5, e02652. [Google Scholar] [CrossRef]
- Neshani, A.; Zare, H.; Eidgahi, M.R.A.; Kakhki, R.K.; Safdari, H.; Khaledi, A.; Ghazvini, K. LL-37: Review of antimicrobial profile against sensitive and antibiotic-resistant human bacterial pathogens. Gene Rep. 2019, 17, 100519. [Google Scholar] [CrossRef]
- de Breij, A.; Riool, M.; Cordfunke, R.A.; Malanovic, N.; de Boer, L.; Koning, R.I.; Ravensbergen, E.; Franken, M.; van der Heijde, T.; Boekema, B.K.; et al. The antimicrobial peptide SAAP-148 combats drug-resistant bacteria and biofilms. Sci. Transl. Med. 2018, 10, 4044. [Google Scholar] [CrossRef]
- Mwangi, J.; Yin, Y.; Wang, G.; Yang, M.; Li, Y.; Zhang, Z.; Lai, R. The antimicrobial peptide ZY4 combats multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii infection. Proc. Natl. Acad. Sci. USA 2019, 116, 26516–26522. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Lan, X.Q.; Du, Y.; Chen, P.Y.; Zhao, J.; Zhao, F.; Lee, W.-H.; Zhang, Y. King cobra peptide OH-CATH30 as a potential candidate drug through clinic drug-resistant isolates. Zool Res. 2018, 39, 87–96. [Google Scholar] [PubMed]
- Barksdale, S.M.; Hrifko, E.J.; van Hoek, M.L. Cathelicidin antimicrobial peptide from Alligator mississippiensis has antibacterial activity against multi-drug resistant Acinetobacter baumanii and Klebsiella pneumoniae. Dev. Comp. Immunol. 2017, 70, 135–144. [Google Scholar] [CrossRef]
- Xie, F.; Zan, Y.; Zhang, X.; Zhang, H.; Jin, M.; Zhang, W.; Zhang, Y.; Liu, S. Differential abilities of mammalian cathelicidins to inhibit bacterial biofilm formation and promote multifaceted immune functions of neutrophils. Int. J. Mol. Sci. 2020, 21, 1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milenic, D.E.; Brady, E.D.; Brechbiel, M.W. Antibody-targeted radiation cancer therapy. Nat. Rev. Drug Discov. 2004, 3, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Dadachova, E.; Casadevall, A. Antibodies as delivery vehicles for radioimmunotherapy of infectious diseases. Expert Opin. Drug Deliv. 2005, 2, 1075–1084. [Google Scholar] [CrossRef]
- Castano, A.P.; Mroz, P.; Wu, M.X.; Hamblin, M.R. Photodynamic therapy plus low-dose cyclophosphamide generates antitumor immunity in a mouse model. Proc. Natl. Acad. Sci. USA 2008, 105, 5495–5500. [Google Scholar] [CrossRef]
- Dai, T.; Tegos, G.P.; Lu, Z.; Huang, L.; Zhiyentayev, T.; Franklin, M.J.; Baer, D.G.; Hamblin, M.R. Photodynamic therapy for Acinetobacter baumannii burn infections in mice. Antimicrob. Agents Chemother. 2009, 53, 3929–3934. [Google Scholar] [CrossRef]
- Li, Z.; Ding, Z.; Liu, Y.; Jin, X.; Xie, J.; Li, T.; Zeng, Z.; Wang, Z.; Liu, J. Phenotypic and genotypic characteristics of biofilm formation in clinical isolates of Acinetobacter baumannii. Infect. Drug Resist. 2021, 14, 2613–2624. [Google Scholar] [CrossRef]
- Donadu, M.G.; Mazzarello, V.; Cappuccinelli, P.; Zanetti, S.; Madléna, M.; Nagy, Á.L.; Stájer, A.; Burián, K.; Gajdács, M. Relationship between the biofilm-forming capacity and antimicrobial resistance in clinical Acinetobacter baumannii isolates: Results from a laboratory-based in vitro study. Microorganisms 2021, 9, 2384. [Google Scholar] [CrossRef]
- Ababneh, Q.; Abulaila, S.; Jaradat, Z. Isolation of extensively drug resistant Acinetobacter baumannii from environmental surfaces inside intensive care units. Am. J. Infect. Control 2021, 50, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Ababneh, Q.; Al-Rousan, E.; Jaradat, Z. Fresh produce as a potential vehicle for transmission of Acinetobacter baumannii. Int. J. Food Contam. 2022, 9, 1–9. [Google Scholar] [CrossRef]
- Greene, C.; Vadlamudi, G.; Newton, D.; Foxman, B.; Xi, C. The influence of biofilm formation and multidrug resistance on environmental survival of clinical and environmental isolates of Acinetobacter baumannii. Am. J. Infect. Control 2016, 44, e65–e71. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.D.; Zhu, Y.; Azad, M.A.K.; Han, M.L.; Wang, J.; Wang, L.; Yu, H.H.; Horne, A.S.; Pinson, J.A.; Rudd, D.; et al. A synthetic lipopeptide targeting top-priority multidrug-resistant Gram-negative pathogens. Nat. Commun. 2022, 13, 1625. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Genes | Role and Functions | Authors |
|---|---|---|
| CsuC CsuD CsuE |
| Wright et al., 2017 [14] |
| OmpA |
| Rumbo et al., 2013 [15] |
| blaPER-1 |
| Rodriguez et al., 2006 [16] |
| abaI |
| Badave et al., 2015 [17] |
| Bap |
| Cao et al., 2014 [18] |
| epsA |
| Russo et al., 2009 [19] |
| bfmS&R |
| Liou et al., 2014 [20] |
| Plant’s Name | Active Compounds | References |
|---|---|---|
| Lythrum salicaria | Hexahydroxy diphenoyl ester vescalagin | [81] |
| Rosa rugosa | Ellagic acid | [82] |
| Terminalia chebula | Terchebulin, Chebulagic acid, Chebulinic acid, Corilagin | [82] |
| Scutellaria baicalensis | Norwogonin, Baicalin, Baicalein | [82] |
| Syzygium aromaticum | Eugenol | [83] |
| Cinnamomum zeylanicum | Trans-cinnamaldehyde | [84] |
| Oreganum vulgare | Carvacrol | [83,84] |
| Green tea Camellia sinensis | Epigallocatechin gallate (EGCG) | [85] |
| Epicatechin | [86] | |
| Theaflavin | [86] | |
| Lyciumchinense Mill. | (+)-Lyoniresinol-3 alpha-O-beta-D-glucopyranoside | [87] |
| Paeonia suffruticosa Andr. | Paeonol | [87] |
| Coptidischinensis Franch. | Berberine | [87] |
| Green tea (Camellia sinesis) | polyphenol, (–)-epigallocatechin- 3-gallate (EGCG) | [88] |
| Pantoea agglomerans | Pantoea Natural Product 3 (PNP-3). | [89] |
| Peptide | Structure | Activity | References |
|---|---|---|---|
| LL-37 (Human cathelicidin) | α-helical | Immunomodulation characteristics, broad-spectrum microbicidal activities. Demonstrated MIC of 16–32 μg/mL against A. baumannii inhibited and dispersed the A. baumannii biofilm in abiotic surfaces at 32 and 64 μg/mL. | [124,125] |
| SAAP-148 | α-helical AMP | Eliminate acute and biofilm-related; inhibit the growth of A. baumannii MDR at a concentration of 6 μg/mL. | [126] |
| ZY4 cathelicidin-BF-15 | Cyclic peptide | Good activity against A. baumannii, including standard clinical MDR strains with MIC values ranging between 4.6 and 9.4 μg/mL. | [127] |
| Naja atra cathelicidin (NA-CATH) | α-helical structure at N-terminal and an unstructured segment at C-terminal | Antimicrobial activity through the membrane lysis by membrane thinning or transient pore formation and inhibiting the bacterial growth at a concentration of 10 μg/mL. | [127,128] |
| AM-CATH36: two fragments AM-CATH28 and AM-CATH21 | Found in American alligator | Inhibit the growth of both drug-resistant and sensitive A. baumannii at the 2.5 μg/mL concentration. | [129] |
| BMAP-27 BMAP-34 mCRAP | Mammalian cathelicidins | It quickly disrupts the bacterial cellular integrity. It has potent action of inhibition against biofilms and exhibit immunomodulatory function. | [130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolma, K.G.; Khati, R.; Paul, A.K.; Rahmatullah, M.; de Lourdes Pereira, M.; Wilairatana, P.; Khandelwal, B.; Gupta, C.; Gautam, D.; Gupta, M.; et al. Virulence Characteristics and Emerging Therapies for Biofilm-Forming Acinetobacter baumannii: A Review. Biology 2022, 11, 1343. https://doi.org/10.3390/biology11091343
Dolma KG, Khati R, Paul AK, Rahmatullah M, de Lourdes Pereira M, Wilairatana P, Khandelwal B, Gupta C, Gautam D, Gupta M, et al. Virulence Characteristics and Emerging Therapies for Biofilm-Forming Acinetobacter baumannii: A Review. Biology. 2022; 11(9):1343. https://doi.org/10.3390/biology11091343
Chicago/Turabian StyleDolma, Karma G., Rachana Khati, Alok K. Paul, Mohammed Rahmatullah, Maria de Lourdes Pereira, Polrat Wilairatana, Bidita Khandelwal, Chamma Gupta, Deepan Gautam, Madhu Gupta, and et al. 2022. "Virulence Characteristics and Emerging Therapies for Biofilm-Forming Acinetobacter baumannii: A Review" Biology 11, no. 9: 1343. https://doi.org/10.3390/biology11091343