
Comparative Chloroplast Genomes of Six Magnoliaceae Species Provide New Insights into Intergeneric Relationships and Phylogeny


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Sequencing
2.2. Chloroplast Genome Assembly and Annotation
2.3. Sequence Divergence, Genome Comparison and Single-Sequence Repeat Analysis
2.4. Phylogenetic Analysis
3. Results
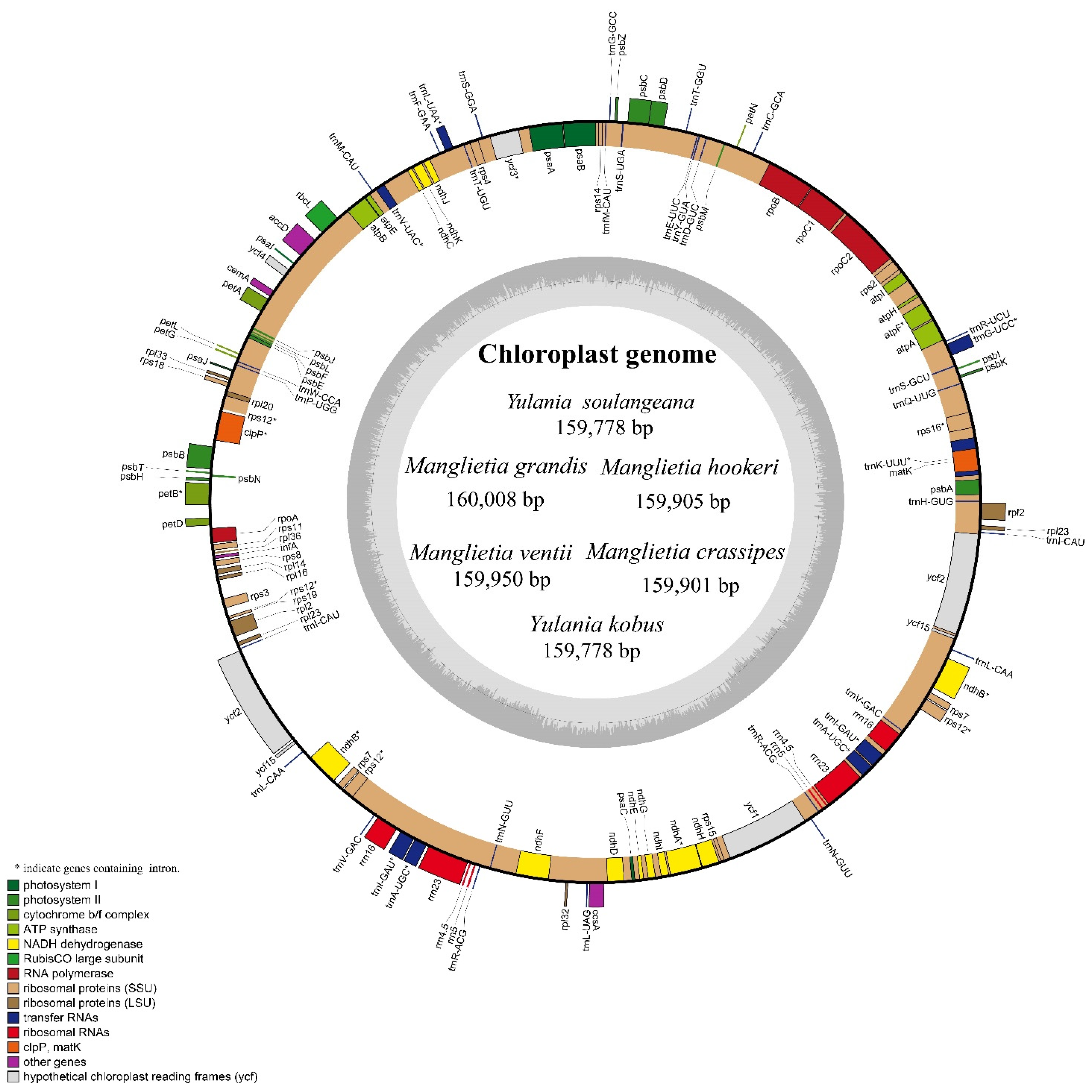
3.1. Characteristics of the Six cp Genomes
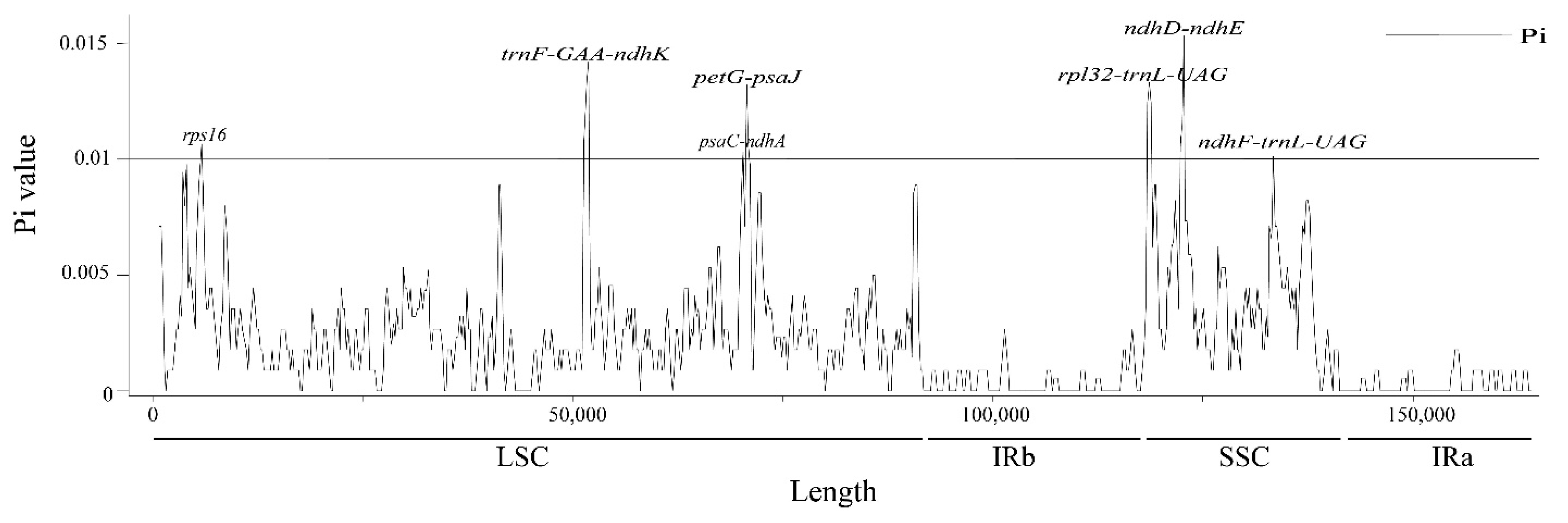
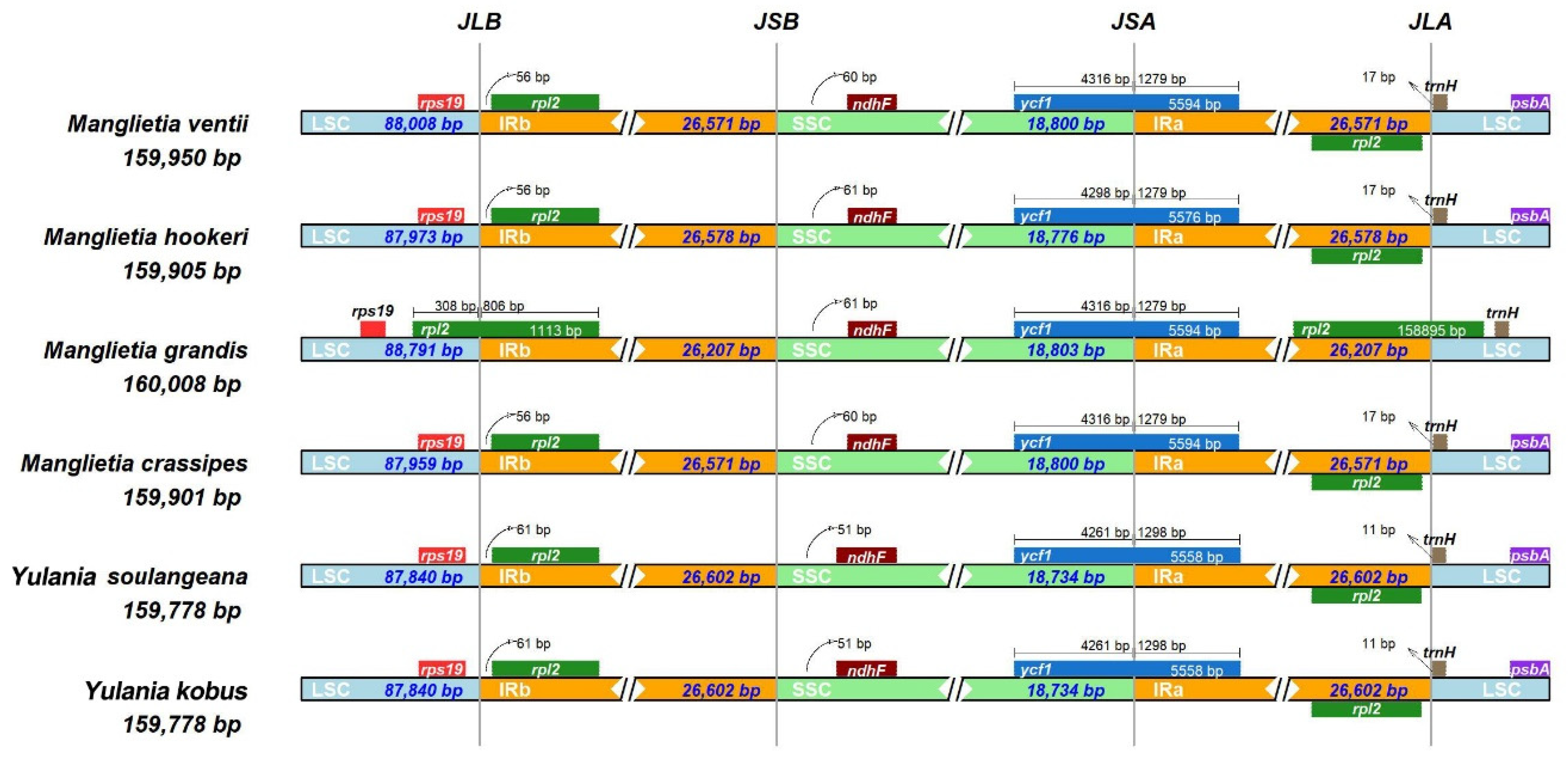
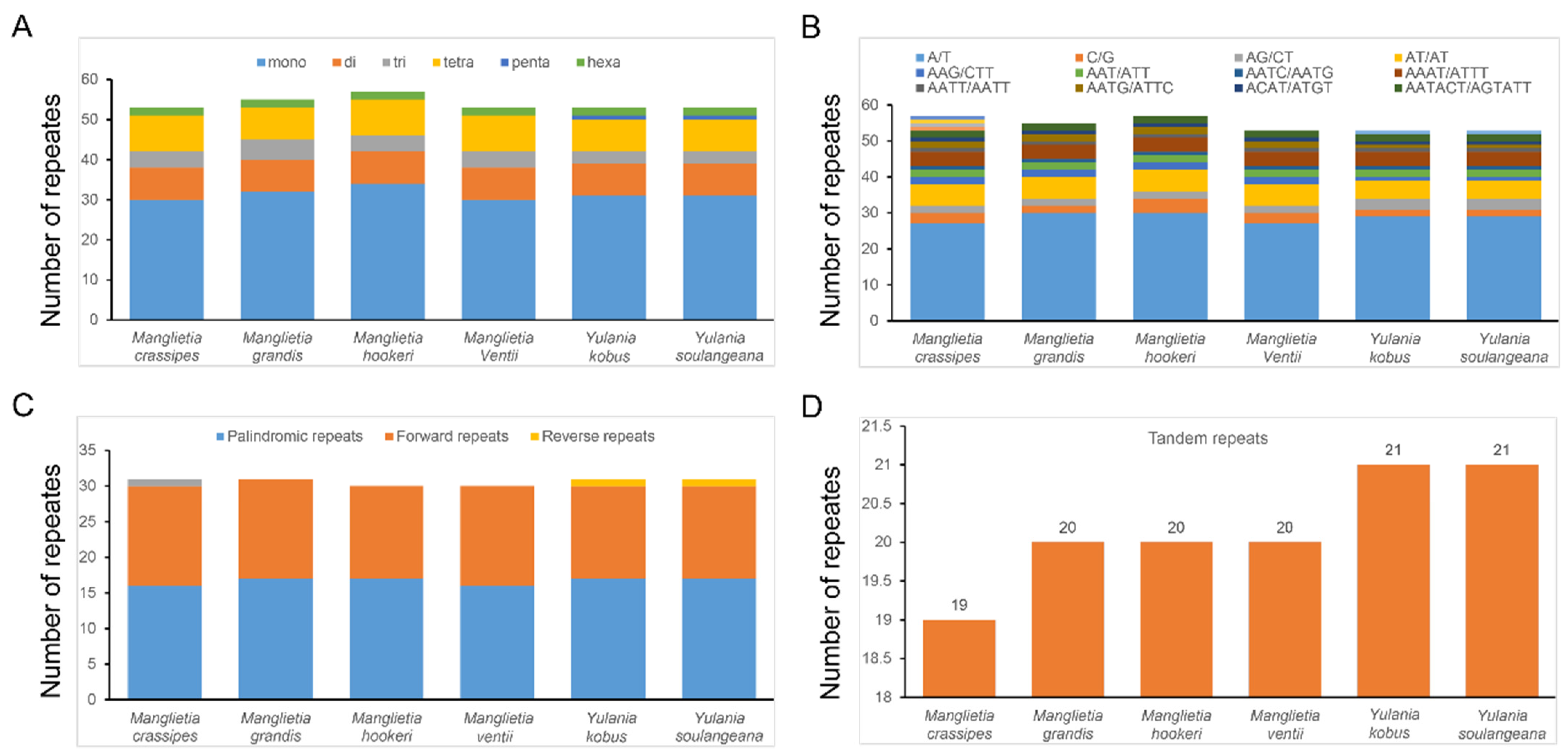
3.2. Comparative Genomic, IR Expansion and Contraction, and SSR Analysis
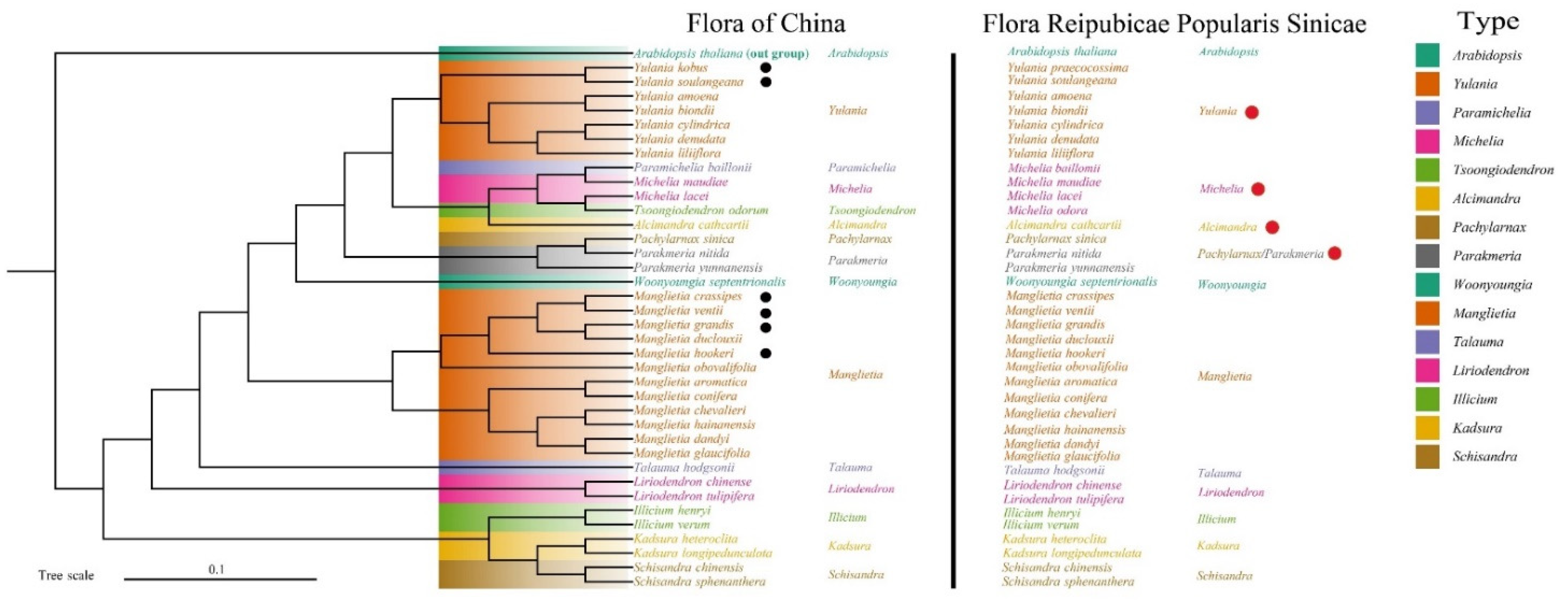
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saldaña, C.L.; Rodriguez-Grados, P.; Chávez-Galarza, J.C.; Feijoo, S.; Guerrero-Abad, J.C.; Vásquez, H.V.; Maicelo, J.L.; Jhoncon, J.H.; Carlos, I.; Arbizu, C.I. Unlocking the complete chloroplast genome of a native tree species from the Amazon basin, capirona (Calycophyllum spruceanum, rubiaceae), and its comparative analysis with other ixoroideae species. Genes 2022, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Li, X.; Chen, X.; Huang, X.L.; Jin, S.H. The complete chloroplast genome of Carya cathayensis and phylogenetic analysis. Genes 2022, 13, 369. [Google Scholar] [CrossRef] [PubMed]
- Antunes, A.M.; Soares, T.N.; Targueta, C.P.; Novaes, E.; Guedes-Coelho, A.S.; Campos-Telles, M.P. The chloroplast genome sequence of Dipteryx alata vog. (Fabaceae: Papilionoideae): Genomic features and comparative analysis with other legume genomes. Braz. J. Bot. 2020, 43, 271–278. [Google Scholar] [CrossRef]
- Feng, S.; Zheng, K.; Jiao, K.; Cai, Y.C.; Chen, C.L.; Mao, Y.Y.; Wang, L.Y.; Zhan, X.R.; Ying, Q.C.; Wang, H.Z. Complete chloroplast genomes of four Physalis species (Solanaceae): Lights into genome structure, comparative analysis, and phylogenetic relationships. BMC Plant Biol. 2020, 20, 242. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Zhu, H.; Wang, J.; Xu, Y.C.; Xu, F.S.; Wang, X.M. Complete chloroplast genome sequence determination of Rheum species and comparative chloroplast genomics for the members of rumiceae. Plant Cell Rep. 2020, 39, 811–824. [Google Scholar] [CrossRef]
- Duan, Y.Z.; Zhang, K. Comparative analysis and phylogenetic evolution of the complete chloroplast genome of Ammopiptanthus. Acta Bot. Boreali-Occident. Sin. 2020, 40, 1323–1332. [Google Scholar]
- Xie, B.B. Phylogeography Study of Siraitia Grosvenorii. Master’s Thesis, Guangxi Normal University, Guilin, Guangxi, China, 2019. [Google Scholar]
- Zhao, S.; Deng, L.H.; Chen, F. Codon usage bias of chloroplast genome in Kandelia obovate. J. For. Environ. 2020, 40, 534–541. [Google Scholar]
- Ngoc, A.L.; Nurhayat, T.; Alden, S.E.; Nguyen, D.H.; Kendra, P.E. Insecticidal and attractant activities of Magnolia citrata leaf essential oil against two major pests from diptera: Aedes aegypti (culicidae) and Ceratitis capitata (tephritidae). Molecules 2021, 26, 2311. [Google Scholar] [CrossRef]
- Timmis, J.N.; Ayliffe, M.A.; Huang, C.Y.; William, M. Endosymbiotic gene transfer: Organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 2004, 5, 123–135. [Google Scholar] [CrossRef]
- Huang, Y.B.; Wang, J.M.; Li, R.X. Evaluation of cold resistance and physiological mechanism of seven Manglietia species. J. Trop. Subtrop. Bot. 2021, 29, 519–529. [Google Scholar] [CrossRef]
- Sun, B.Y.; Liu, Y.; Xue, Y.Y.; Zhang, Y. The anti-tumor activity validation of Magnolia sargentiana. and Manglietia hookeri. on base of reverse molecular docking. Comput. Appl. Chem. 2019, 36, 337–344. [Google Scholar]
- Zhu, B.; Qian, F.; Wang, X.S.; Liu, Y.L. The phylogeny of magnoliaceae based on chloroplast genome. J. Biol. 2022, 39, 53–58. [Google Scholar] [CrossRef]
- Sima, Y.K.; Lu, S.G.; Han, M.Y.; Ma, H.F.; Xu, T.; Xu, L.; Hao, J.B.; Li, D.; Chen, S.Y. Advances in magnoliaceae systematic research. J. West China For. Sci. 2012, 41, 116–127. [Google Scholar] [CrossRef]
- Hutchinson, J. The Genera of Flowering Plants; Clarendon Press: Oxford, UK, 1964; pp. 44–57. [Google Scholar] [CrossRef]
- Baranova, M.A.; Jeffrey, C.; Wu, Q.G. Leaf Epidermis Features of Magnoliaceae and Their Systematic Significance, with Special Reference to the Genera Parakmeria, Manglietiastrum and Woonyoungia; Science Press: Beijing, China, 2000; pp. 143–152. [Google Scholar]
- Liu, Y.H.; Fan, H.M.; Chen, Z.Y. Proceeding of the International Symposium on the Family Magnoliaceae, Guangzhou, China, 18–22 May 1998; Science Press: Beijing, China, 2000; pp. 3–13. [Google Scholar]
- Chen, D.Q.; Xu, F.X. Observation of pollen morphology of two species from Talauma. Guihaia 2008, 3, 317–319. [Google Scholar] [CrossRef]
- Wang, Y.C.; Zhou, H.Y.; Liu, X.Q. Chloroplast genomic variation in Euonymus maackii Rupr. and its differentiation time in euonymus. Forests 2022, 13, 265. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Zheng, W.; Zhang, H.; Wang, Q.M.; Gao, Y.; Zhang, Z.H.; Sun, Y.X. Complete chloroplast genome sequence of Clivia miniata and its characteristics. Acta Hortic. Sin. 2020, 47, 2439–2450. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq-versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, 6–11. [Google Scholar] [CrossRef]
- Mu, Y.N. Phylogenic and Taxonomic Research of Hypolepisin China Based on the Complete Chloroplast Genome. Ph.D. Thesis, Harbin Normal University, Harbin, China, 2020. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSPv5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Huang, X.; Tan, W.; Li, F.; Liao, R.; Guo, Z.; Shi, T.; Gao, Z. The chloroplast genome of Prunus zhengheensis: Genome comparative and phylogenetic relationships analysis. Gene 2021, 793, 145751. [Google Scholar] [CrossRef]
- Thiel, T.; Michalek, W.; Varshney, R.K.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-Markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.W.; Cui, Y.X.; Wang, Q.; Xu, Z.C.; Wang, Y.; Lin, Y.L.; Song, J.Y.; Yao, H. Identifcation and phylogenetic analysis of fve Crataegus species (Rosaceae) based on complete chloroplast genomes. Planta 2021, 254, 14. [Google Scholar] [CrossRef] [PubMed]
- Han, C.Y.; Ding, R.; Zong, X.Y.; Zhang, L.J.; Chen, X.H.; Qu, B. Structural characterization of Platanthera ussuriensis chloroplast genome and comparative analyses with other species of Orchidaceae. BMC Genom. 2022, 23, 84. [Google Scholar] [CrossRef]
- Li, L.X.; Peng, J.Y.; Wang, D.W.; Duan, A.A. Analysis of chloroplast genome characteristics phylogeny and codon preference in Docynia longiunguis. Chin. J. Biotechnol. 2022, 38, 328–342. [Google Scholar] [CrossRef]
- Guzmán-Díaz, S.; Núñez, F.A.A.; Veltjen, E.; Asselman, P.; Larridon, I.; Samain, M.-S. Comparison of magnoliaceae plastomes: Adding neotropical Magnolia to the discussion. Plants 2022, 11, 448. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.M.; Yuan, L.; Tang, T.L.; Hou, T.; Pan, K.; Wie, N. The complete chloroplast genome sequence of Alpinia oxyphylla Miq. and comparison analysis within the Zingiberaceae family. PLoS ONE 2019, 14, e0218817. [Google Scholar] [CrossRef] [PubMed]
- Santana, A.; Gomes, T.; Nimz, T.; Vieira, L.N.; Guerra, M.P.; Nodari, R.O.; Souza, E.M.; Pedrosa, F.O.; Rogalski, M. The complete plastome of macaw palm [Acrocomia aculeata (Jacq.) lodd. ex mart.] and extensive molecular analyses of the evolution of plastid genes in arecaceae. Planta 2018, 247, 1011–1030. [Google Scholar] [CrossRef]
- Dong, W.; Liu, J.; Yu, J.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef]
- Alzahrani, A.; Albokhari, J.; Yaradua, S.; Abba, A. Comparative analysis of chloroplast genomes of four medicinal capparaceae species: Genome structures, phylogenetic relationships and adaptive evolution. Plants 2021, 10, 1229. [Google Scholar] [CrossRef]
- Saina, J.K.; Li, Z.Z.; Mekbib, Y.; Gichira, A.W.; Liao, Y.Y. Transcriptome sequencing and microsatellite marker discovery in Ailanthus altissima (Mill.) Swingle (Simaroubaceae). Mol. Biol. Rep. 2021, 48, 2007–2023. [Google Scholar] [CrossRef]
- Menezes, A.; Resende-Moreira, L.C.; Buzatti, O.; Nazareno, A.G.; Carlsen, M.; Lobo, F.P.; Kalapothakis, E.; Lovato, M.B. Chloroplast genomes of Byrsonima species (malpighiaceae): Comparative analysis and screening of high divergence sequences. Sci. Rep. 2018, 8, 2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angiosperm Phylogeny Group; Chase, M.W.; Christenhusz, M.J.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar]
- Wang, Y.-B.; Liu, B.-B.; Nie, Z.-L.; Chen, H.-F.; Chen, F.-J.; Figlar, R.B.; Wen, J. Major clades and a revised classification of Magnolia and magnoliaceae based on whole plastid genome sequences via genome skimming. J. Syst. Evol. 2020, 58, 673–695. [Google Scholar] [CrossRef]
- Figlar, R.B. Proleptic branch initiation in michelia and magnolia subgenus Yulania provides basis for combinations in subfamily magnolioideae. In Proceedings of the International Symposium on the Family Magnoliaceae, Guangzhou, China, 18–22 May 1998; Science Press: Beijing, China, 2000; pp. 14–25. [Google Scholar]
- Fu, D.L.; Zhao, D.X.; Sun, J.H.; Xu, J.W.; Wang, J. Component of volatile oils in 10 chinese species of Yulania genus and its phylogenetic implicatior. Sci. Silvae Sin. 2005, 41, 68–74. [Google Scholar]
- Gong, X.; Pang, Y.Z.; Yang, Z.Y. The cross-compatibility of magnoliaceae. Plant Divers. 2001, 23, 339–344. [Google Scholar]
- Dong, S.-S.; Wang, Y.-L.; Xia, N.-H.; Liu, Y.; Liu, M.; Lian, L.; Li, N.; Li, L.-F.; Lang, X.-A.; Gong, Y.-Q.; et al. Plastid and nuclear phylogenomic incongruences and biogeographic implications of Magnolia s.l. (Magnoliaceae). J. Syst. Evol. 2022, 60, 1–15. [Google Scholar] [CrossRef]
- Kim, S. Phylogenetic relationships in family magnoliaceae inferred from ndhF sequences. Am. J. Bot. 2001, 88, 717–728. [Google Scholar] [CrossRef]
- Xie, X.N.; Huang, R.; Li, F.; Tian, E.; Li, C.; Chao, Z. Phylogenetic position of Bupleurum sikangense inferred from the complete chloroplast genome sequence. Gene 2021, 798, 145801. [Google Scholar] [CrossRef]
- Wang, M.; Peng, J.Q.; Cao, J.W.; Cao, F.X.; Li, J.J.; Xue, C.; Wu, Y. Studies on genetic diversity of 48 wild species of magnoliaceae in Yunnan. J. Trop. Subtrop. Bot. 2020, 28, 277–284. [Google Scholar] [CrossRef]
- Xu, F.X.; Wu, Q.G. Pollen morphology of Paramichelia and Tsoongiodedron from China. Acta Bot. Boreali-Occident. Sin. 1995, 6, 3. [Google Scholar]
- Jin, H.; Shi, S.H.; Pan, H.C.; Huang, Y.L.; Zhang, H.D. Phylogenetic relationships between Michelia (Magnoliaceae) and its related genera based on the matk gene sequence. Acta Sci. Nat. Univ. Sunyatseni 1999, 38, 91–97. [Google Scholar] [CrossRef]
- Wu, Z.Y. Sinicae agendae academiae sinicae edita. In Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 2022. [Google Scholar]
- Hu, X.X.; Zheng, W.J. Parakmeria, a new genus of Magnoliaceae of southwestern China. J. Syst. Evol. 1951, 1, 1. Available online: http://journal.ucas.ac.cn/CN/Y1951/V1/I1/1 (accessed on 18 March 2022).
- Wu, Z.Y. The areal-types of Chinese genera of seed plants. Plant Divers. 1991, 13, 1. Available online: https://journal.kib.ac.cn/EN/Y1991/V13/IS4/1 (accessed on 27 March 2022).
- Xiong, Q.; Zhang, M.; Yu, D.P. Cloning and phylogenetic analysis of b-class mads-box gene in Parakmeria omeiensis. Mol. Plant Breed. 2018, 16, 5540–5548. [Google Scholar] [CrossRef]
- Sima, Y.K.; Wang, J.; Cao, L.M.; Wang, B.Y.; Wang, Y.H. Prefoliation features of the magnoliaceae and their systematic significance. J. Yunnan Univ. 2001, 23, 71–78. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Protection Grade | Sampling Area | Longitude/Latitude |
|---|---|---|---|---|
| Manglietia | Manglietia crassipes Y.L.Law | - | Guangxi, China | 109°50′ E/23°40′ N |
| Manglietia grandis Hu & W.C.Cheng | II | Yunnan, China | 104°33′ E/22°48′ N | |
| Manglietia hookeri Cubitt & W.W.Sm. | - | Yunnan, China | 99°55′ E/21°10′ N | |
| Manglietia ventii N.V.Tiep. | II | Yunnan, China | 102°10′ E/24°23′ N | |
| Yulania | Yulania kobus (DC.) Spach | - | Yunnan, China | 102°10′ E/24°23′ N |
| Yulania soulangeana (Soul.-Bod.) D.L.Fu | - | Yunnan, China | 102°10′ E/24°23′ N |
| Species | Manglietia crassipes | Manglietia grandis | Manglietia hookeri | Manglietia ventii | Yulania kobus | Yulania soulangeana |
|---|---|---|---|---|---|---|
| Total length (bp) | 159,901 | 160,008 | 159,905 | 159,950 | 159,778 | 159,778 |
| LSC length (bp) | 87,959 | 88,534 | 87,973 | 88,008 | 87,840 | 87,753 |
| SSC length (bp) | 18,800 | 18,803 | 18,776 | 18,800 | 18,734 | 18,734 |
| IR length (bp) | 26,571 | 26,207 | 26,578 | 26,571 | 26,602 | 26,602 |
| Overall GC content (%) | 39.3 | 39.3 | 39.3 | 39.3 | 39.3 | 39.3 |
| Total gene number | 131 | 131 | 131 | 131 | 131 | 131 |
| GenBank accession | MW415418 | MW415419 | MW415420 | MW415421 | MW415416 | MW415417 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Tian, J.; Xu, L.; Zhao, X.; Song, Y.; Wang, D. Comparative Chloroplast Genomes of Six Magnoliaceae Species Provide New Insights into Intergeneric Relationships and Phylogeny. Biology 2022, 11, 1279. https://doi.org/10.3390/biology11091279
Yang L, Tian J, Xu L, Zhao X, Song Y, Wang D. Comparative Chloroplast Genomes of Six Magnoliaceae Species Provide New Insights into Intergeneric Relationships and Phylogeny. Biology. 2022; 11(9):1279. https://doi.org/10.3390/biology11091279
Chicago/Turabian StyleYang, Lin, Jinhong Tian, Liu Xu, Xueli Zhao, Yuyang Song, and Dawei Wang. 2022. "Comparative Chloroplast Genomes of Six Magnoliaceae Species Provide New Insights into Intergeneric Relationships and Phylogeny" Biology 11, no. 9: 1279. https://doi.org/10.3390/biology11091279