
New Insights into the Diversity of Branchiomeric Muscle Development: Genetic Programs and Differentiation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
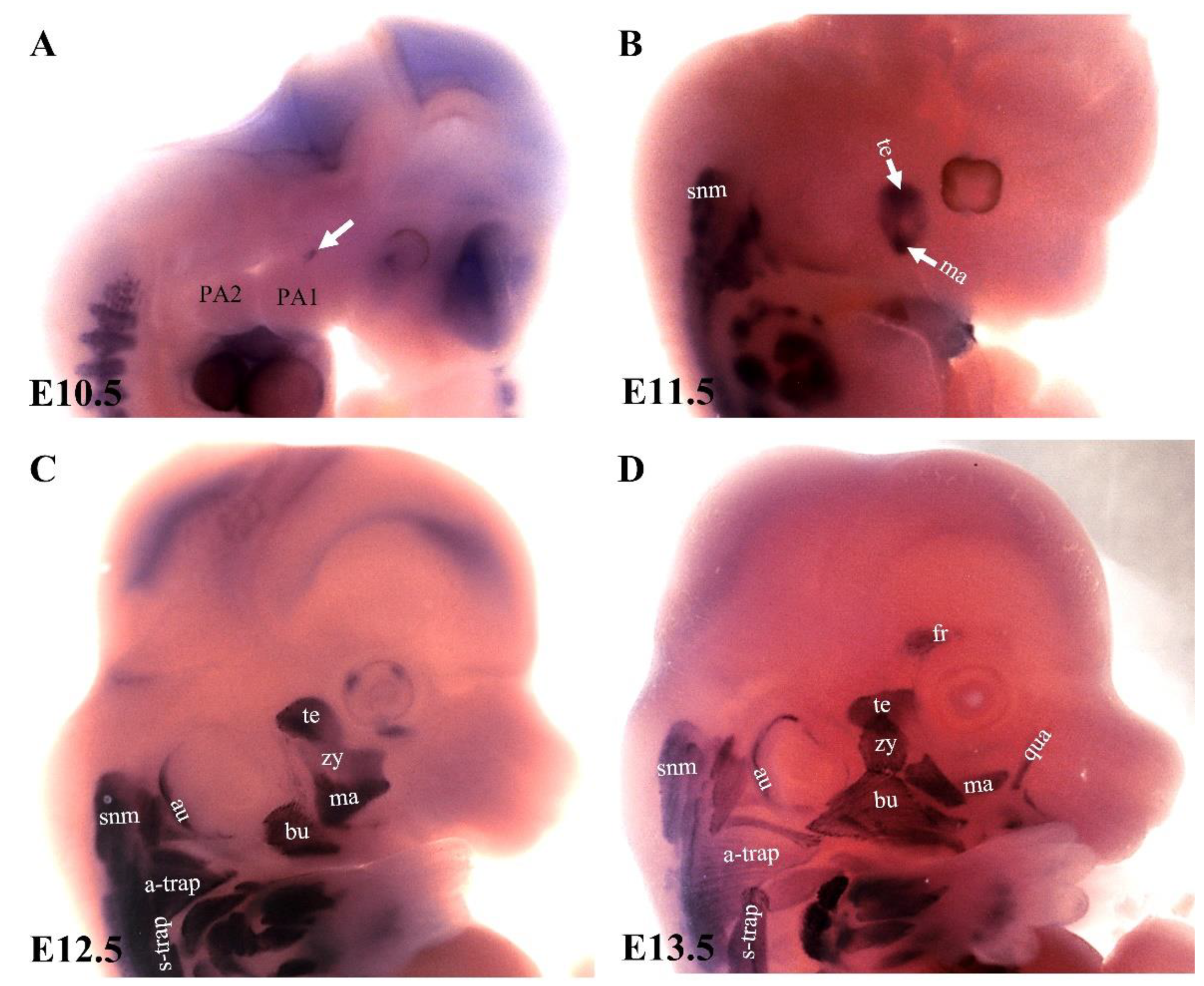{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
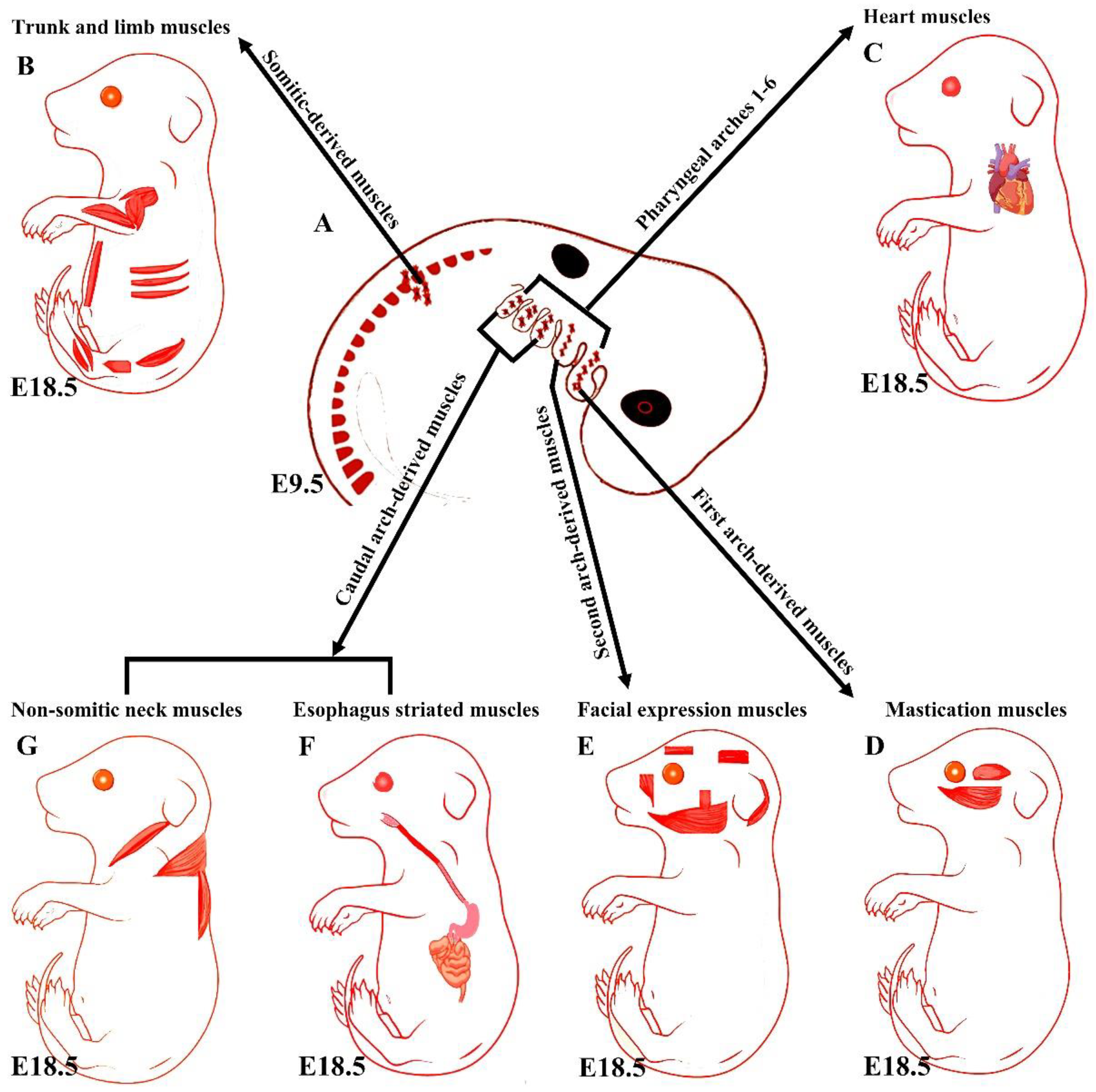
2. An Overview of the Early Trunk and Limb Muscle Development
3. An Overview of Early Branchiomeric Muscle Development
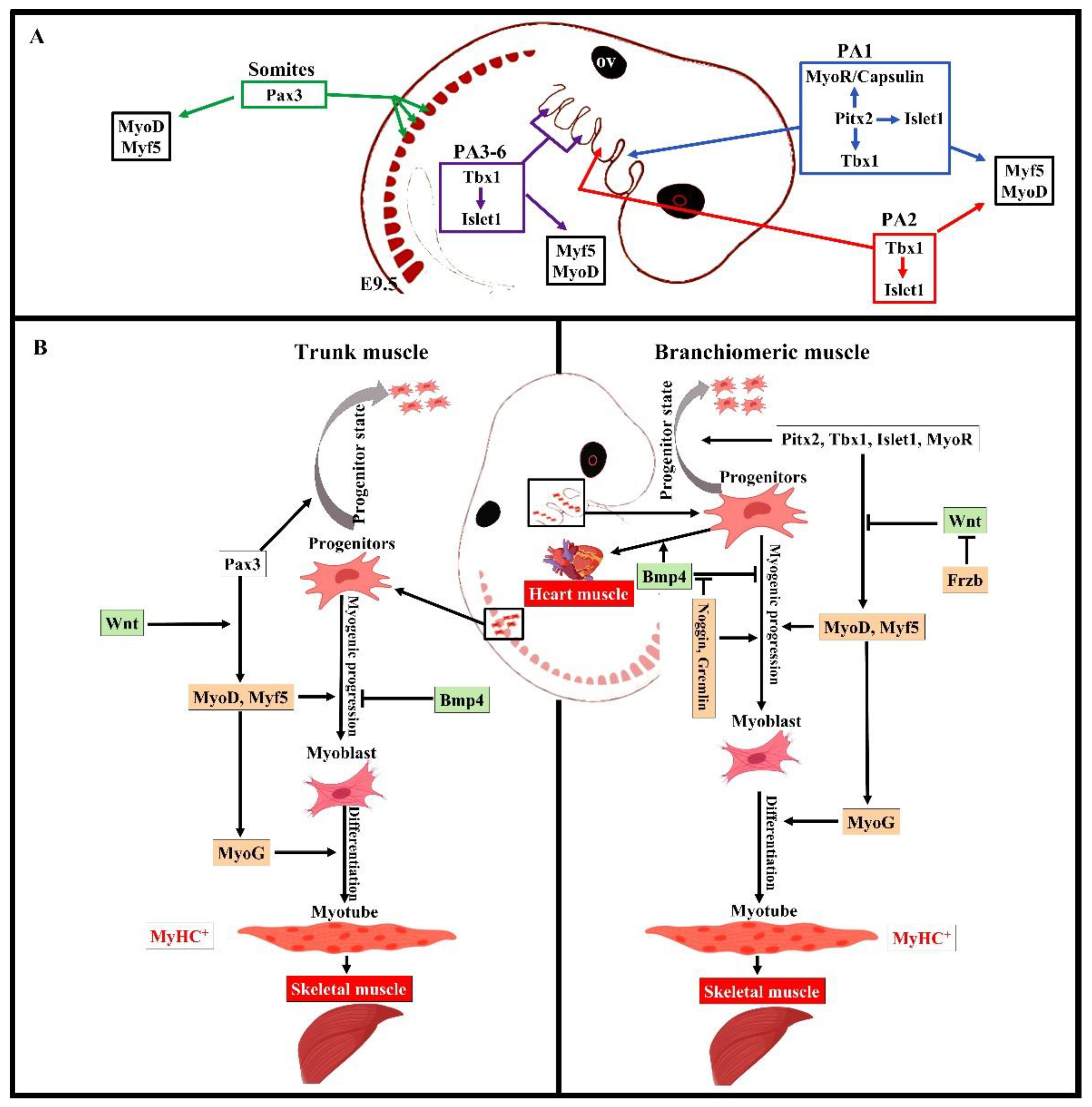
4. Distinct Genetic Programs in Branchiomeric Muscles
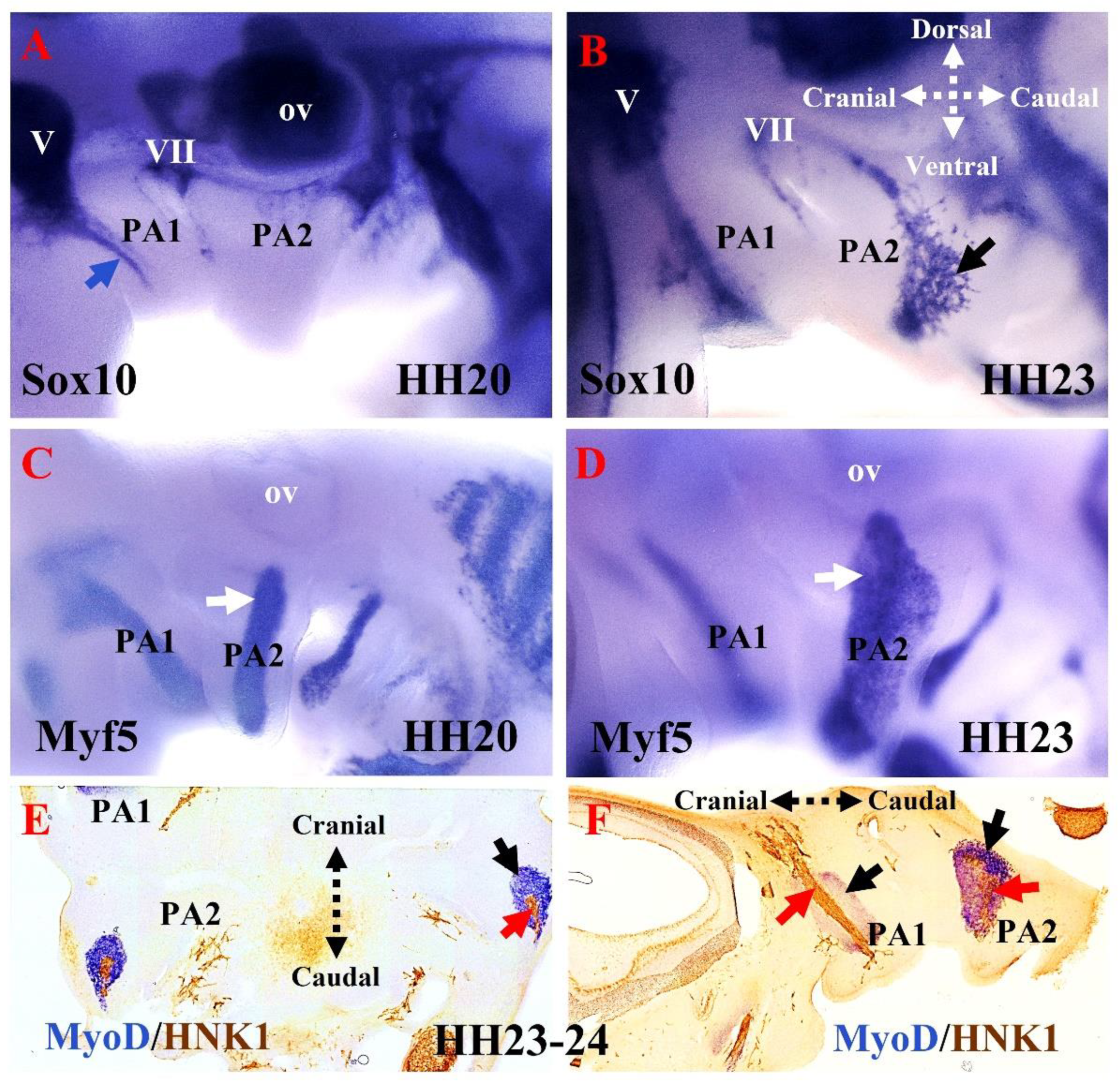
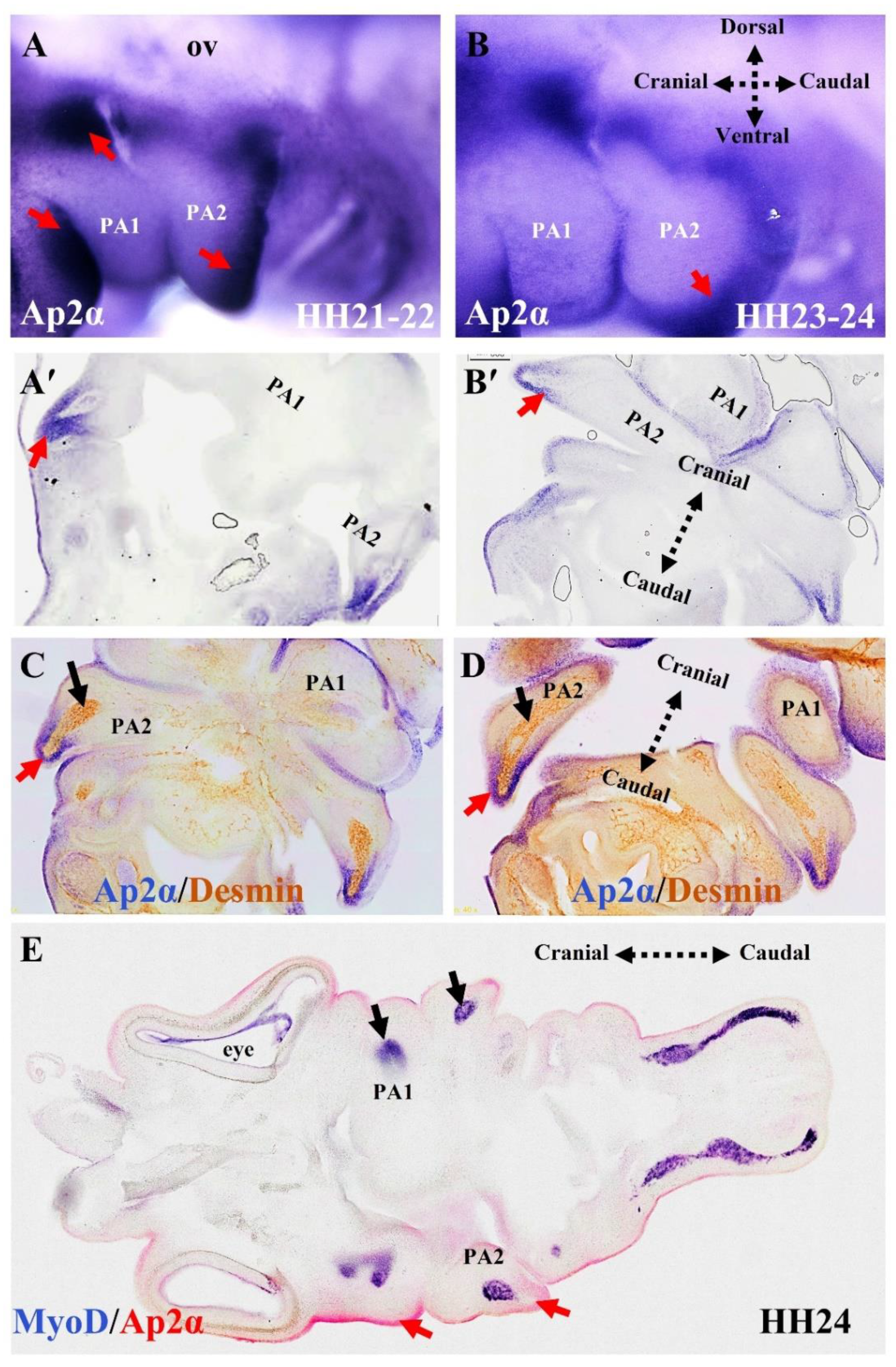
5. The Relationship between Branchiomeric Muscle Progenitors and Neural Crest Cells in Chicken Embryos
6. Branchiomeric Muscles Are Heterogeneous in Terms of the Onset of Their Myogenic Differentiation
7. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lescroart, F.; Kelly, R.G.; Le Garrec, J.F.; Nicolas, J.F.; Meilhac, S.M.; Buckingham, M. Clonal analysis reveals common lineage relationships between head muscles and second heart field derivatives in the mouse embryo. Development 2010, 137, 3269–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, J.M.; Hawrot, K.; Sharpe, C.; Noble, A.; Wood, W.M.; Jorge, E.C.; Goldhamer, D.J.; Kardon, G.; Dietrich, S. The emergence of Pax7-expressing muscle stem cells during vertebrate head muscle development. Front. Aging Neurosci. 2015, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckingham, M.; Vincent, S.D. Distinct and dynamic myogenic populations in the vertebrate embryo. Curr. Opin. Genet. Dev. 2009, 19, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Sambasivan, R.; Kuratani, S.; Tajbakhsh, S. An eye on the head: The development and evolution of craniofacial muscles. Development 2011, 138, 2401–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckingham, M.; Rigby, P.W.J. Gene Regulatory Networks and Transcriptional Mechanisms that Control Myogenesis. Dev. Cell 2014, 28, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heude, E.; Tesarova, M.; Sefton, E.M.; Jullian, E.; Adachi, N.; Grimaldi, A.; Zikmund, T.; Kaiser, J.; Kardon, G.; Kelly, R.G.; et al. Unique morphogenetic signatures define mammalian neck muscles and associated connective tissues. eLife 2018, 7, e40179. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.R.; Bassel-Duby, R.; Hawkins, A.; Chang, P.; Valdez, R.; Wu, H.; Gan, L.; Shelton, J.M.; Richardson, J.A.; Olson, E.N. Control of facial muscle development by MyoR and capsulin. Science 2002, 298, 2378–2381. [Google Scholar] [CrossRef]
- Shih, H.P.; Gross, M.K.; Kioussi, C. Cranial muscle defects of Pitx2 mutants result from specification defects in the first branchial arch. Proc. Natl. Acad. Sci. USA 2007, 104, 5907–5912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noden, D.M.; Francis-West, P. The differentiation and morphogenesis of craniofacial muscles. Dev. Dyn. 2006, 235, 1194–1218. [Google Scholar] [CrossRef]
- Noden, D.M.; Marcucio, R.; Borycki, A.G.; Emerson, C.P., Jr. Differentiation of avian craniofacial muscles: I. Patterns of early regulatory gene expression and myosin heavy chain synthesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1999, 216, 96–112. [Google Scholar] [CrossRef]
- Tzahor, E. Head Muscle Development. In Vertebrate Myogenesis: Stem Cells and Precursors; Brand-Saberi, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 123–142. [Google Scholar]
- Buckingham, M.; Relaix, F. PAX3 and PAX7 as upstream regulators of myogenesis. Semin. Cell Dev. Biol. 2015, 44, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Ordahl, C.P. Early stages of chick somite development. Anat. Embryol. 1995, 191, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Shih, H.P.; Gross, M.K.; Kioussi, C. Muscle development: Forming the head and trunk muscles. Acta Histochem. 2008, 110, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Brent, A.E.; Schweitzer, R.; Tabin, C.J. A somitic compartment of tendon progenitors. Cell 2003, 113, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Parker, M.H.; Seale, P.; Rudnicki, M.A. Looking back to the embryo: Defining transcriptional networks in adult myogenesis. Nat. Rev. Genet. 2003, 4, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Pu, Q.; Abduelmula, A.; Masyuk, M.; Theiss, C.; Swandulla, D.; Hans, M.; Patel, K.; Brand-Saberi, B.; Huang, R.J. The dermomyotome ventrolateral lip is essential for the hypaxial myotome formation. BMC Dev. Biol. 2013, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, S.; Schubert, F.R.; Lumsden, A. Control of dorsoventral pattern in the chick paraxial mesoderm. Development 1997, 124, 3895–3908. [Google Scholar] [CrossRef]
- Hollway, G.; Currie, P. Vertebrate myotome development. Birth Defects Res. Part C Embryo Today Rev. 2005, 75, 172–179. [Google Scholar] [CrossRef]
- Ordahl, C.P.; Berdougo, E.; Venters, S.J.; Denetclaw, W. The dermomyotome dorsomedial lip drives growth and morphogenesis of both the primary myotome and dermomyotome epithelium. Development 2001, 128, 1731–1744. [Google Scholar] [CrossRef]
- Vasyutina, E.; Stebler, J.; Brand-Saberi, B.; Schulz, S.; Raz, E.; Birchmeier, C. CXCR4 and Gab1 cooperate to control the development of migrating muscle progenitor cells. Genes Dev. 2005, 19, 2187–2198. [Google Scholar] [CrossRef] [Green Version]
- Lours-Calet, C.; Alvares, L.E.; El-Hanfy, A.S.; Gandesha, S.; Walters, E.H.; Sobreira, D.R.; Wotton, K.R.; Jorge, E.C.; Lawson, J.A.; Lewis, A.K. Evolutionarily conserved morphogenetic movements at the vertebrate head–trunk interface coordinate the transport and assembly of hypopharyngeal structures. Dev. Biol. 2014, 390, 231–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, N.; Pascual-Anaya, J.; Hirai, T.; Higuchi, S.; Kuroda, S.; Kuratani, S. Stepwise participation of HGF/MET signaling in the development of migratory muscle precursors during vertebrate evolution. Zool. Lett. 2018, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sefton, E.M.; Gallardo, M.; Kardon, G. Developmental origin and morphogenesis of the diaphragm, an essential mammalian muscle. Dev. Biol. 2018, 440, 64–73. [Google Scholar] [CrossRef]
- Comai, G.; Heude, E.; Mella, S.; Paisant, S.; Pala, F.; Gallardo, M.; Langa, F.; Kardon, G.; Gopalakrishnan, S.; Tajbakhsh, S. A distinct cardiopharyngeal mesoderm genetic hierarchy establishes antero-posterior patterning of esophagus striated muscle. eLife 2019, 8, e47460. [Google Scholar] [CrossRef] [PubMed]
- Lescroart, F.; Dumas, C.E.; Adachi, N.; Kelly, R.G. Emergence of heart and branchiomeric muscles in cardiopharyngeal mesoderm. Exp. Cell Res. 2022, 410, 112931. [Google Scholar] [CrossRef]
- Yahya, I.; Boing, M.; Pu, Q.; Puchert, M.; Oedemis, V.; Engele, J.; Brand-Saberi, B.; Morosan-Puopolo, G. Cxcr4 and Sdf-1 are critically involved in the formation of facial and non-somitic neck muscles. Sci. Rep. 2020, 10, 5049. [Google Scholar] [CrossRef] [Green Version]
- Yahya, I.; Morosan-Puopolo, G.; Brand-Saberi, B. The CXCR4/SDF-1 Axis in the Development of Facial Expression and Non-somitic Neck Muscles. Front. Cell Dev. Biol. 2020, 8, 615264. [Google Scholar] [CrossRef]
- Tzahor, E.; Evans, S.M. Pharyngeal mesoderm development during embryogenesis: Implications for both heart and head myogenesis. Cardiovasc. Res. 2011, 91, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Diogo, R.; Kelly, R.G.; Christiaen, L.; Levine, M.; Ziermann, J.M.; Molnar, J.L.; Noden, D.M.; Tzahor, E. A new heart for a new head in vertebrate cardiopharyngeal evolution. Nature 2015, 520, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Lescroart, F.; Hamou, W.; Francou, A.; Theveniau-Ruissy, M.; Kelly, R.G.; Buckingham, M. Clonal analysis reveals a common origin between nonsomite-derived neck muscles and heart myocardium. Proc. Natl. Acad. Sci. USA 2015, 112, 1446–1451. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, D.; Chen, K.; Jubran, A.; Ramirez, A.; Astrof, S. Endothelium in the pharyngeal arches 3, 4 and 6 is derived from the second heart field. Dev. Biol. 2017, 421, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Yahya, I.; Al Haj, A.; Brand-Saberi, B.; Morosan-Puopolo, G. Chicken Second Branchial Arch Progenitor Cells Contribute to Heart Musculature in vitro and in vivo. Cells Tissues Organs 2020, 209, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.G.; Brown, N.A.; Buckingham, M.E. The arterial pole of the mouse heart forms from Fgf10-expressing cells in pharyngeal mesoderm. Dev. Cell 2001, 1, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, S.; Comai, G.; Sambasivan, R.; Francou, A.; Kelly, R.G.; Tajbakhsh, S. A Cranial Mesoderm Origin for Esophagus Striated Muscles. Dev. Cell 2015, 34, 694–704. [Google Scholar] [CrossRef]
- Adachi, N.; Bilio, M.; Baldini, A.; Kelly, R.G. Cardiopharyngeal mesoderm origins of musculoskeletal and connective tissues in the mammalian pharynx. Development 2020, 147, dev185256. [Google Scholar] [CrossRef]
- Bothe, I.; Tenin, G.; Oseni, A.; Dietrich, S. Dynamic control of head mesoderm patterning. Development 2011, 138, 2807–2821. [Google Scholar] [CrossRef] [Green Version]
- Harel, I.; Nathan, E.; Tirosh-Finkel, L.; Zigdon, H.; Guimaraes-Camboa, N.; Evans, S.M.; Tzahor, E. Distinct origins and genetic programs of head muscle satellite cells. Dev. Cell 2009, 16, 822–832. [Google Scholar] [CrossRef] [Green Version]
- Sambasivan, R.; Gayraud-Morel, B.; Dumas, G.; Cimper, C.; Paisant, S.; Kelly, R.G.; Tajbakhsh, S. Distinct regulatory cascades govern extraocular and pharyngeal arch muscle progenitor cell fates. Dev. Cell 2009, 16, 810–821. [Google Scholar] [CrossRef] [Green Version]
- Gage, P.J.; Suh, H.; Camper, S.A. Dosage requirement of Pitx2 for development of multiple organs. Development 1999, 126, 4643–4651. [Google Scholar] [CrossRef]
- Kitamura, K.; Miura, H.; Miyagawa-Tomita, S.; Yanazawa, M.; Katoh-Fukui, Y.; Suzuki, R.; Ohuchi, H.; Suehiro, A.; Motegi, Y.; Nakahara, Y. Mouse Pitx2 deficiency leads to anomalies of the ventral body wall, heart, extra-and periocular mesoderm and right pulmonary isomerism. Development 1999, 126, 5749–5758. [Google Scholar] [CrossRef]
- Lu, M.-F.; Pressman, C.; Dyer, R.; Johnson, R.L.; Martin, J.F. Function of Rieger syndrome gene in left–right asymmetry and craniofacial development. Nature 1999, 401, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.G.; Jerome-Majewska, L.A.; Papaioannou, V.E. The del22q11.2 candidate gene Tbx1 regulates branchiomeric myogenesis. Hum. Mol. Genet. 2004, 13, 2829–2840. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Sun, X.; Liu, W.; Ai, D.; Klysik, E.; Lu, M.-F.; Hadley, J.; Antoni, L.; Chen, L.; Baldini, A. Pitx2 promotes development of splanchnic mesoderm-derived branchiomeric muscle. Development 2006, 133, 4891–4899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.; Dilworth, F.J. Differential modulation of cell cycle progression distinguishes members of the myogenic regulatory factor family of transcription factors. FEBS J. 2013, 280, 3991–4003. [Google Scholar] [CrossRef]
- Evans, D.J.; Noden, D.M. Spatial relations between avian craniofacial neural crest and paraxial mesoderm cells. Dev. Dyn. 2006, 235, 1310–1325. [Google Scholar] [CrossRef]
- Rinon, A.; Lazar, S.; Marshall, H.; Buchmann-Moller, S.; Neufeld, A.; Elhanany-Tamir, H.; Taketo, M.M.; Sommer, L.; Krumlauf, R.; Tzahor, E. Cranial neural crest cells regulate head muscle patterning and differentiation during vertebrate embryogenesis. Development 2007, 134, 3065–3075. [Google Scholar] [CrossRef] [Green Version]
- Tirosh-Finkel, L.; Zeisel, A.; Brodt-Ivenshitz, M.; Shamai, A.; Yao, Z.; Seger, R.; Domany, E.; Tzahor, E. BMP-mediated inhibition of FGF signaling promotes cardiomyocyte differentiation of anterior heart field progenitors. Development 2010, 137, 2989–3000. [Google Scholar] [CrossRef] [Green Version]
- Tzahor, E.; Kempf, H.; Mootoosamy, R.C.; Poon, A.C.; Abzhanov, A.; Tabin, C.J.; Dietrich, S.; Lassar, A.B. Antagonists of Wnt and BMP signaling promote the formation of vertebrate head muscle. Genes Dev. 2003, 17, 3087–3099. [Google Scholar] [CrossRef] [Green Version]
- Blentic, A.; Tandon, P.; Payton, S.; Walshe, J.; Carney, T.; Kelsh, R.N.; Mason, I.; Graham, A. The emergence of ectomesenchyme. Dev. Dyn. 2008, 237, 592–601. [Google Scholar] [CrossRef] [Green Version]
- Graham, A. The neural crest. Curr. Biol. 2003, 13, R381–R384. [Google Scholar] [CrossRef] [Green Version]
- Giovannone, D.; Ortega, B.; Reyes, M.; El-Ghali, N.; Rabadi, M.; Sao, S.; de Bellard, M.E. Chicken trunk neural crest migration visualized with HNK1. Acta Histochem. 2015, 117, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minoux, M.; Rijli, F.M. Molecular mechanisms of cranial neural crest cell migration and patterning in craniofacial development. Development 2010, 137, 2605–2621. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Asai, R.; Maruyama, K.; Kurihara, Y.; Nakanishi, T.; Kurihara, H.; Miyagawa-Tomita, S. Postotic and preotic cranial neural crest cells differently contribute to thyroid development. Dev. Biol. 2016, 409, 72–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escot, S.; Blavet, C.; Hartle, S.; Duband, J.L.; Fournier-Thibault, C. Misregulation of SDF1-CXCR4 signaling impairs early cardiac neural crest cell migration leading to conotruncal defects. Circ. Res. 2013, 113, 505–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirby, M.L.; Gale, T.F.; Stewart, D.E. Neural crest cells contribute to normal aorticopulmonary septation. Science 1983, 220, 1059–1061. [Google Scholar] [CrossRef]
- Siismets, E.M.; Hatch, N.E. Cranial neural crest cells and their role in the pathogenesis of craniofacial anomalies and coronal craniosynostosis. J. Dev. Biol. 2020, 8, 18. [Google Scholar] [CrossRef]
- Zalc, A.; Sinha, R.; Gulati, G.S.; Wesche, D.J.; Daszczuk, P.; Swigut, T.; Weissman, I.L.; Wysocka, J. Reactivation of the pluripotency program precedes formation of the cranial neural crest. Science 2021, 371, eabb4776. [Google Scholar] [CrossRef]
- Betancur, P.; Bronner-Fraser, M.; Sauka-Spengler, T. Genomic code for Sox10 activation reveals a key regulatory enhancer for cranial neural crest. Proc. Natl. Acad. Sci. USA 2010, 107, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Grenier, J.; Teillet, M.A.; Grifone, R.; Kelly, R.G.; Duprez, D. Relationship between Neural Crest Cells and Cranial Mesoderm during Head Muscle Development. PLoS ONE 2009, 4, e4381. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Saint-Jeannet, J.-P. Induction of the neural crest and the opportunities of life on the edge. Dev. Biol. 2004, 275, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yahya, I.; Böing, M.; Brand-Saberi, B.; Morosan-Puopolo, G. How to distinguish between different cell lineages sharing common markers using combinations of double in-situ-hybridization and immunostaining in avian embryos: CXCR4-positive mesodermal and neural crest-derived cells. Histochem. Cell Biol. 2021, 155, 145–155. [Google Scholar] [CrossRef]
- Pusch, C.; Hustert, E.; Pfeifer, D.; Südbeck, P.; Kist, R.; Roe, B.; Wang, Z.; Balling, R.; Blin, N.; Scherer, G. The SOX10/Sox10 gene from human and mouse: Sequence, expression, and transactivation by the encoded HMG domain transcription factor. Hum. Genet. 1998, 103, 115–123. [Google Scholar] [CrossRef]
- Cheng, Y.-C.; Cheung, M.; Abu-Elmagd, M.M.; Orme, A.; Scotting, P.J. Chick sox10, a transcription factor expressed in both early neural crest cells and central nervous system. Dev. Brain Res. 2000, 121, 233–241. [Google Scholar] [CrossRef]
- Buac, K.; Watkins-Chow, D.E.; Loftus, S.K.; Larson, D.M.; Incao, A.; Gibney, G.; Pavan, W.J. A Sox10 expression screen identifies an amino acid essential for Erbb3 function. PLoS Genet. 2008, 4, e1000177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabian, P.; Tseng, K.-C.; Thiruppathy, M.; Arata, C.; Chen, H.-J.; Smeeton, J.; Nelson, N.; Crump, J.G. Lifelong single-cell profiling of cranial neural crest diversification in zebrafish. Nat. Commun. 2022, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.S.; Charney, R.M.; García-Castro, M.I. Specification and formation of the neural crest: Perspectives on lineage segregation. Genesis 2019, 57, e23276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betters, E.; Charney, R.M.; Garcia-Castro, M.I. Early specification and development of rabbit neural crest cells. Dev. Biol. 2018, 444 (Suppl. S1), S181–S192. [Google Scholar] [CrossRef]
- Yagi, H.; Yanagisawa, M.; Suzuki, Y.; Nakatani, Y.; Ariga, T.; Kato, K.; Robert, K.Y. HNK-1 epitope-carrying tenascin-C spliced variant regulates the proliferation of mouse embryonic neural stem cells. J. Biol. Chem. 2010, 285, 37293–37301. [Google Scholar] [CrossRef] [Green Version]
- Yahya, I.; Böing, M.; Hockman, D.; Brand-Saberi, B.; Morosan-Puopolo, G. The Emergence of Embryonic Myosin Heavy Chain during Branchiomeric Muscle Development. Life 2022, 12, 785. [Google Scholar] [CrossRef]
- Agarwal, M.; Sharma, A.; Kumar, P.; Kumar, A.; Bharadwaj, A.; Saini, M.; Kardon, G.; Mathew, S.J. Myosin heavy chain-embryonic regulates skeletal muscle differentiation during mammalian development. Development 2020, 147, dev184507. [Google Scholar] [CrossRef]
- Schubert, F.R.; Singh, A.J.; Afoyalan, O.; Kioussi, C.; Dietrich, S. To roll the eyes and snap a bite–function, development and evolution of craniofacial muscles. Semin. Cell Dev. Biol. 2018, 91, 31–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitzmann, M.; Fernandez, A. Crosstalk between cell cycle regulators and the myogenic factor MyoD in skeletal myoblasts. Cell. Mol. Life Sci. CMLS 2001, 58, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Burke, A.C.; Nowicki, J. A new view of patterning domains in the vertebrate mesoderm. Dev. Cell 2003, 4, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Amthor, H.; Christ, B.; Patel, K. A molecular mechanism enabling continuous embryonic muscle growth-a balance between proliferation and differentiation. Development 1999, 126, 1041–1053. [Google Scholar] [CrossRef] [PubMed]
- McMahon, J.A.; Takada, S.; Zimmerman, L.B.; Fan, C.-M.; Harland, R.M.; McMahon, A.P. Noggin-mediated antagonism of BMP signaling is required for growth and patterning of the neural tube and somite. Genes Dev. 1998, 12, 1438–1452. [Google Scholar] [CrossRef] [Green Version]
- Tirosh-Finkel, L.; Elhanany, H.; Rinon, A.; Tzahor, E. Mesoderm progenitor cells of common origin contribute to the head musculature and the cardiac outflow tract. Development 2006, 133, 1943–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Scheven, G.; Alvares, L.E.; Mootoosamy, R.C.; Dietrich, S. Neural tube derived signals and Fgf8 act antagonistically to specify eye versus mandibular arch muscles. Development 2006, 133, 2731–2745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penn, B.H.; Bergstrom, D.A.; Dilworth, F.J.; Bengal, E.; Tapscott, S.J. A MyoD-generated feed-forward circuit temporally patterns gene expression during skeletal muscle differentiation. Genes Dev. 2004, 18, 2348–2353. [Google Scholar] [CrossRef] [Green Version]
- Beylkin, D.H.; Allen, D.L.; Leinwand, L.A. MyoD, Myf5, and the calcineurin pathway activate the developmental myosin heavy chain genes. Dev. Biol. 2006, 294, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Weiss, A.; McDonough, D.; Wertman, B.; Acakpo-Satchivi, L.; Montgomery, K.; Kucherlapati, R.; Leinwand, L.; Krauter, K. Organization of human and mouse skeletal myosin heavy chain gene clusters is highly conserved. Proc. Natl. Acad. Sci. USA 1999, 96, 2958–2963. [Google Scholar] [CrossRef] [Green Version]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.; Leinwand, L.A.; Reggiani, C. Developmental myosins: Expression patterns and functional significance. Skelet Muscle 2015, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yahya, I.; Hockman, D.; Brand-Saberi, B.; Morosan-Puopolo, G. New Insights into the Diversity of Branchiomeric Muscle Development: Genetic Programs and Differentiation. Biology 2022, 11, 1245. https://doi.org/10.3390/biology11081245
Yahya I, Hockman D, Brand-Saberi B, Morosan-Puopolo G. New Insights into the Diversity of Branchiomeric Muscle Development: Genetic Programs and Differentiation. Biology. 2022; 11(8):1245. https://doi.org/10.3390/biology11081245
Chicago/Turabian StyleYahya, Imadeldin, Dorit Hockman, Beate Brand-Saberi, and Gabriela Morosan-Puopolo. 2022. "New Insights into the Diversity of Branchiomeric Muscle Development: Genetic Programs and Differentiation" Biology 11, no. 8: 1245. https://doi.org/10.3390/biology11081245