
Highly Efficient Use of Infrared Spectroscopy (ATR-FTIR) to Identify Aphid Species


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Measurement
2.2. Statistical Analyses
3. Results
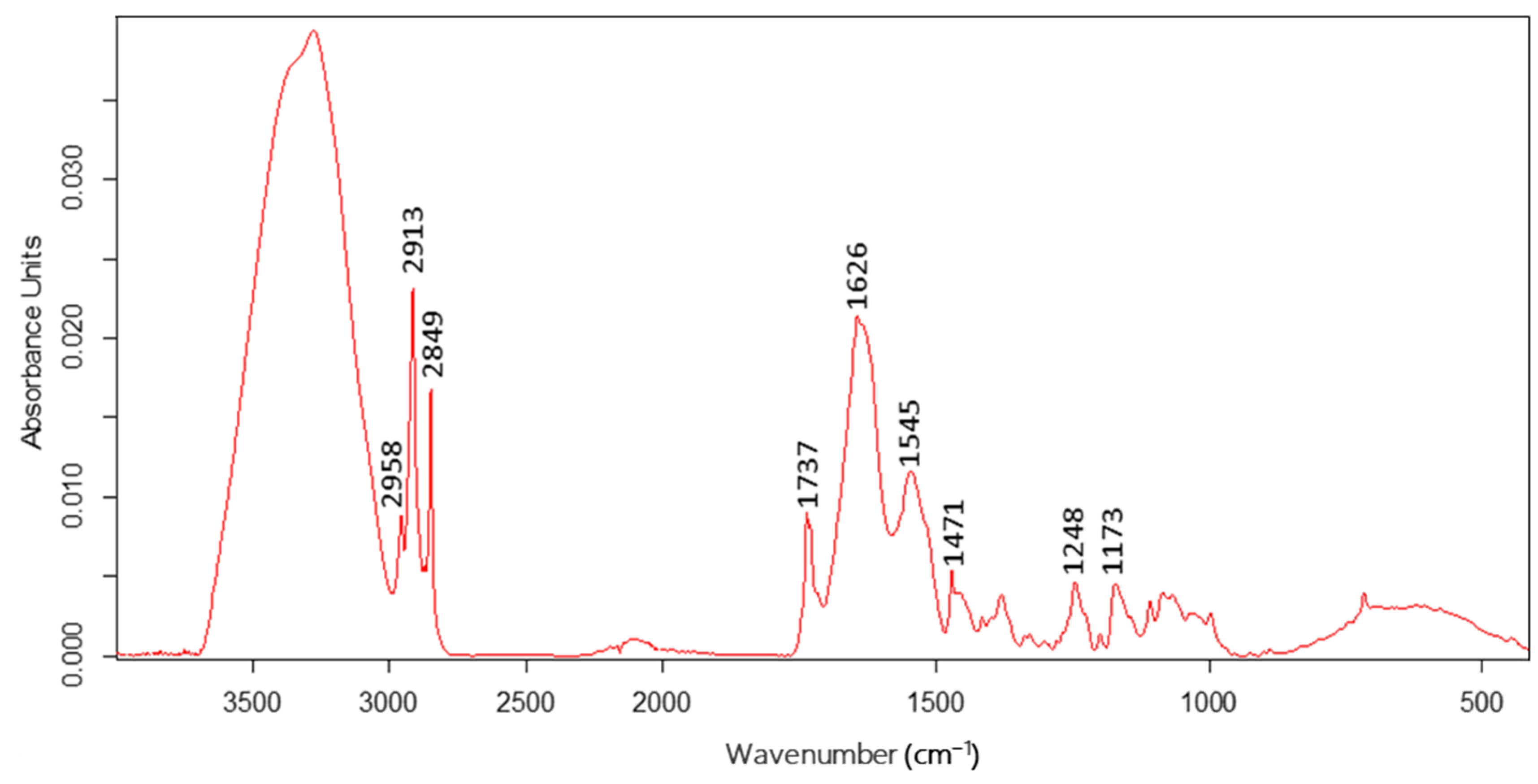
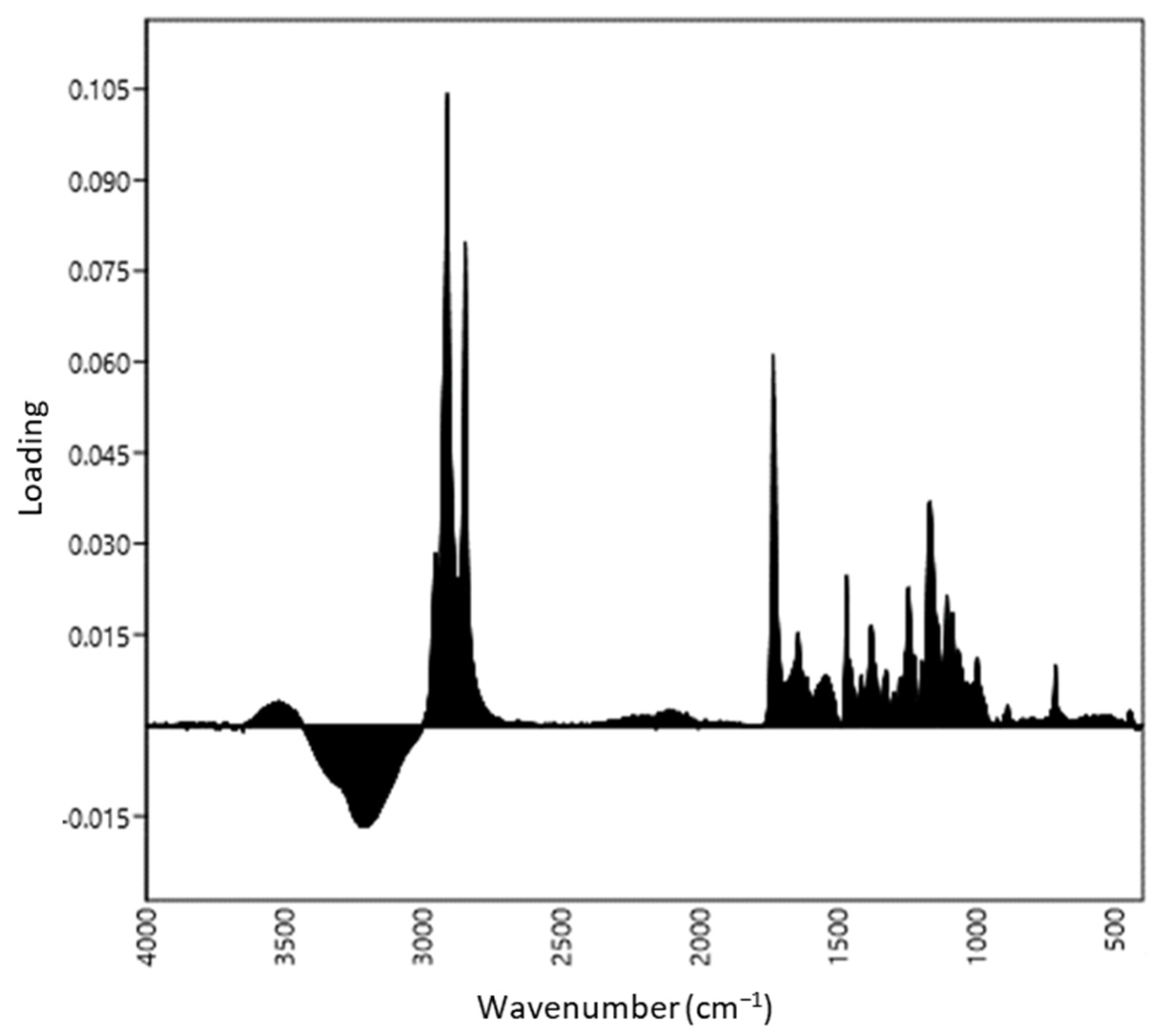
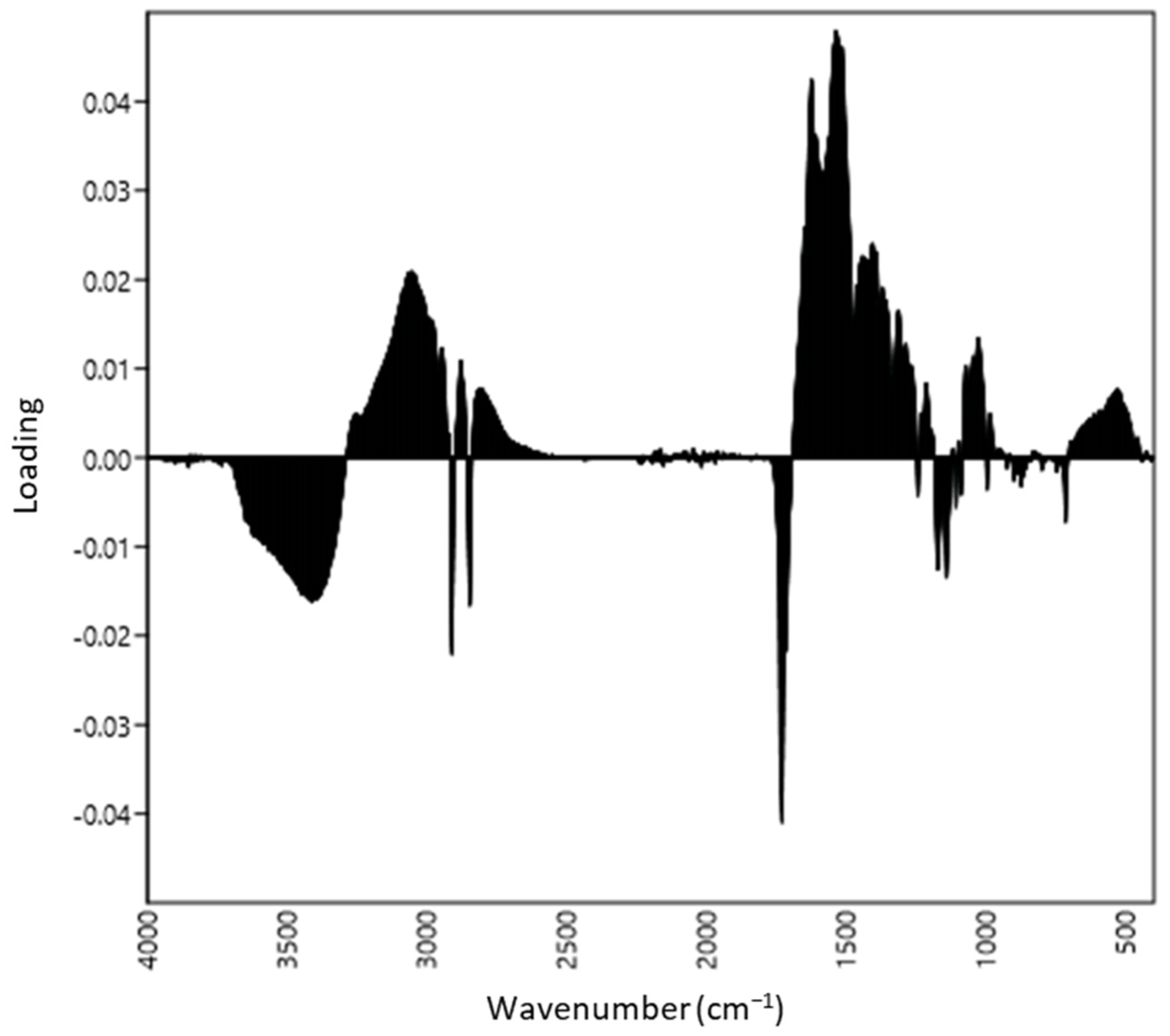
3.1. Distinction of ATR-FTIR Ranges
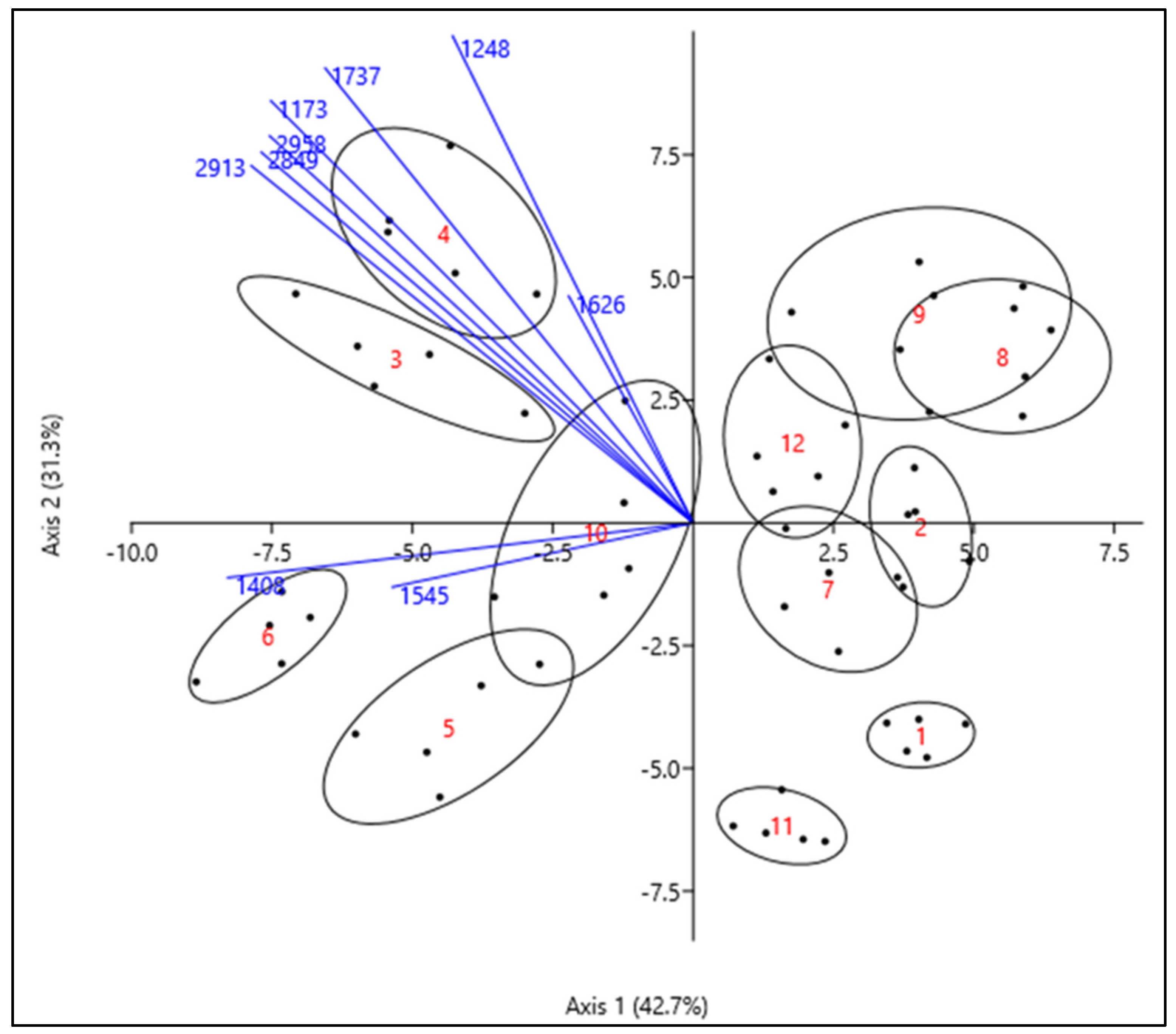
3.2. Species Identification on the Basis of Chemical Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ong, O.T.W.; Kho, E.A.; Esperança, P.M.; Freebairn, C.; Dowell, F.E.; Devine, G.J.; Churcher, T.S. Ability of near-infrared spectroscopy and chemometrics to predict the age of mosquitoes reared under different conditions. Parasite Vectors 2020, 13, 160. [Google Scholar] [CrossRef]
- Mwanga, E.P.; Mapua, S.A.; Siria, D.J.; Ngowo, H.S.; Nangacha, F.; Mgando, J.; Baldini, F.; Jiménez, M.G.; Ferguson, H.M.; Wynne, K.; et al. Using mid-infrared spectroscopy and supervised machine-learning to identify vertebrate blood meals in the malaria vector, Anopheles arabiensis. Malar. J. 2019, 18, 187. [Google Scholar] [CrossRef]
- González Jiménez, M.; Babayan, S.A.; Khazaeli, P.; Doyle, M.; Walton, F.; Reedy, E.; Glew, T.; Viana, M.; Ranford-Cartwright, L.; Niang, A.; et al. Prediction of mosquito species and population age structure using mid-infrared spectroscopy and supervised machine learning. Wellcome Open Res. 2019, 4, 76. [Google Scholar] [CrossRef]
- Baker, M.J.; Trevisan, J.; Bassan, P.; Bhargava, R.; Butler, H.J.; Dorling, K.M.; Fielden, P.R.; Fogarty, S.W.; Fullwood, N.J.; Heys, K.A.; et al. Using Fourier transform IR spectroscopy to analyze biological materials. Nat. Protoc. 2014, 9, 1771–1791. [Google Scholar] [CrossRef]
- Talari, A.C.S.; Garcia Martinez, M.A.; Movasaghi, Z.; Rehman, S.; Ur Rehman, I. Advances in Fourier transform infrared (FTIR) spectroscopy of biological tissues. Appl. Spectrosc. Rev. 2017, 52, 456–506. [Google Scholar] [CrossRef]
- Oliveira, J.S.; Baia, T.C.; Gama, R.A.; Lima, K.M.G. Development of a novel nondestructive method based on spectral fingerprint for determination of abused drug in insects: An alternative entomotoxicology approach. Microchem. J. 2014, 115, 39–46. [Google Scholar] [CrossRef]
- Baia, T.C.; Gama, R.A.; Lima, L.A.S.; Lima, K.M.G. FTIR microspectroscopy coupled with variable selection methods for the identification of flunitrazepam in necrophagous flies. Anal. Methods 2016, 8, 968–972. [Google Scholar] [CrossRef]
- Fischnaller, S.; Dowell, F.E.; Lusser, A.; Schlick-Steiner, B.C.; Steiner, F.M. Nondestructive species identification of Drosophila obscura and D. subobscura (Diptera) using near-infrared spectroscopy. Fly 2012, 6, 284–289. [Google Scholar] [CrossRef]
- Barbosa, T.M.; de Lima, L.A.; Dos Santos, M.C.D.; Vasconcelos, S.D.; Gama, R.A.; Lima, K.M.G. A novel use of infra-red spectroscopy (NIRS and ATR-FTIR) coupled with variable selection algorithms for the identification of insect species (Diptera: Sarcophagidae) of medico-legal relevance. Acta Trop. 2018, 185, 1–12. [Google Scholar] [CrossRef]
- Dowell, F.E.; Parker, A.G.; Benedict, M.Q.; Robinson, A.S.; Broce, A.B.; Wirtz, R.A. Sex separation of tsetse fly pupae using near-infrared spectroscopy. Bull. Entomol. Res. 2005, 95, 249–257. [Google Scholar] [CrossRef]
- Dowell, F.E.; Throne, J.E.; Wang, D.; Baker, J.E. Identifying stored-grain insects using near-infrared spectroscopy. J. Econ. Entomol. 1999, 92, 165–169. [Google Scholar] [CrossRef]
- Jia, F.; Maghirang, E.; Dowell, F.; Abel, C.; Ramaswamy, S. Differentiating tobacco budworm and corn earworm using near-infrared spectroscopy. J. Econ. Entomol. 2007, 100, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Siegwart, M.; Bouvier, F.; Maugin, S.; Lecomte, A.; Lavigne, C. Differentiating oriental fruit moth and codling moth (Lepidoptera: Tortricidae) larvae using near-infrared spectroscopy. J. Econ. Entomol. 2015, 108, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Antonialli-Junior, W.F.; Lima, S.M.; Andrade, L.H.C.; Súarez, Y.R. Comparative study of the cuticular hydrocarbon in queens, workers and males of Ectatomma vizottoi (Hymenoptera, Formicidae) by Fourier transform-infrared photoacoustic spectroscopy. Genet. Mol. Res. 2007, 6, 492–499. Available online: https://www.geneticsmr.com/year2007/vol6-3/pdf/gmr0324.pdf (accessed on 6 May 2022).
- Antonialli-Junior, W.F.; Súarez, Y.R.; Izida, T.; Andrade, L.H.C.; Lima, S.M. Intra-and interspecifc variation of cuticular hydrocarbon composition in two Ectatomma species (Hymenoptera: Formicidae) based on Fourier transform infrared photoacoustic spectroscopy. Genet. Mol. Res. 2008, 7, 559–566. [Google Scholar] [CrossRef]
- Klarica, J.; Bittner, L.; Pallua, J.; Pezzei, C.; Huck-Pezzei, V.; Dowell, F.; Schied, J.; Bonn, G.K.; Huck, C.; Schlick-Steiner, B.C.; et al. Near-infrared imaging spectroscopy as a tool to discriminate two cryptic Tetramorium ant species. J. Chem. Ecol. 2011, 37, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Keszthelyi, S.; Pónya, Z.; Csóka, Á.; Bázár, G.; Morschhauser, T.; Donkó, T. Non-destructive imaging and spectroscopic techniques to investigate the hidden-lifestyle arthropod pests: A review. J. Plant Dis. Prot. 2020, 127, 283–295. [Google Scholar] [CrossRef]
- Dharmaraj, S.; Jamaludin, A.S.; Razak, H.M.; Valliappan, R.; Ahmad, N.A.; Harn, G.L.; Ismail, Z. The classification of Phyllanthus niruri Linn. according to location by infrared spectroscopy. Vib. Spectrosc. 2006, 41, 68–72. [Google Scholar] [CrossRef]
- Aldrich, B.T.; Maghirang, E.B.; Dowell, F.E.; Kambhampati, S. Identification of termite species and subspecies of the genus Zootermopsis using near-infrared reflectance spectroscopy. J. Insect Sci. 2007, 7, 18. [Google Scholar] [CrossRef]
- Perez-Mendoza, J.; Dowell, F.E.; Broce, A.B.; Throne, J.E.; Wirtz, R.A.; Xie, F.; Fabrick, J.A.; Baker, J.E. Chronological age-grading of house flies by using near-infrared spectroscopy. J. Med. Entomol. 2002, 39, 499–508. [Google Scholar] [CrossRef]
- Ami, D.; Natalello, A.; Zullini, A.; Doglia, S.M. Fourier transform infrared microspectroscopy as a new tool for nematode studies. FEBS Lett. 2004, 576, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.L.; Hands, J.R.; Fullwood, L.M.; Smith, J.A.; Baker, M.J. Rapid discrimination of maggots utilising ATR-FTIR spectroscopy. Forensic Sci. Int. 2015, 249, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J. Near-infrared spectroscopy (NIRS) for taxonomic entomology: A brief review. J. Appl. Entomol. 2020, 144, 241–250. [Google Scholar] [CrossRef]
- Wang, Y.; Nansen, C.; Zhang, Y. Integrative insect taxonomy based on morphology, mitochondrial DNA, and hyperspectral reflectance profiling. Zool. J. Linn. Soc. 2016, 177, 378–394. [Google Scholar] [CrossRef]
- Durak, R.; Depciuch, J.; Kapusta, I.; Kisała, J.; Durak, T. Changes in chemical composition and accumulation of cryoprotectants as the adaptation of anholocyclic aphid Cinara tujafilina to overwintering. Int. J. Mol. Sci. 2021, 22, 511. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: https://paleo.carleton.ca/2001_1/past/past.pdf (accessed on 30 October 2014).
- Natural History Museum, University of Oslo. Available online: https://www.nhm.uio.no/english/research/resources/past/index.html (accessed on 25 August 2021).
- Warren, J.A.; Ratnasekera, T.D.P.; Campbell, D.A.; Anderson, G.S. Initial investigations of spectral measurements to estimate the time within stages of Protophormia terraenovae (Robineau-Desvoidy) (Diptera: Calliphoridae). Forensic Sci. Int. 2017, 278, 205–216. [Google Scholar] [CrossRef]
- Warren, J.A.; Ratnasekera, T.D.P.; Campbell, D.A.; Anderson, G.S. Spectral signatures of immature Lucilia sericata (Meigen) (Diptera: Calliphoridae). Insects 2017, 8, 34. [Google Scholar] [CrossRef]
- Rodríguez-Fernández, J.I.; de Carvalho, C.J.; Pasquini, C.; de Lima, M.G.; Moura, M.O.; Arízaga, G.G.C. Barcoding without DNA? Species identification using near infrared spectroscopy. Zootaxa 2011, 2933, 46–54. [Google Scholar] [CrossRef]
- Sinclair, B.J. Linking energetics and overwintering in temperate insects. J. Therm. Biol. 2015, 54, 5–11. [Google Scholar] [CrossRef]
- Durak, R.; Durak, T. Metabolic response of aphid Cinara tujafilina to cold stress. Biology 2021, 10, 1288. [Google Scholar] [CrossRef]
- Kaneko, F.; Katagiri, C.; Sazaki, G.; Nagashima, K. ATR FTIR spectroscopic study on insect body surface lipids rich in methylene-interrupted diene. J. Phys. Chem. B 2018, 122, 12322–12330. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, F.; Katagiri, C.; Nagashima, K.; Sazaki, G.; Ikemoto, Y. Cuticular lipid topology on insect body surfaces studied by synchrotron radiation FTIR ATR microspectroscopy. J. Phys. Chem. B 2021, 125, 9757–9767. [Google Scholar] [CrossRef]
- Kot, I.; Kmiec, K. Study on intensity of infestation, biology and harmfulness of woolly beech aphid (Phyllaphis fagi L.) on Fagus sylvatica (L.). Acta Sci. Pol. Hortorum Cultus 2012, 11, 3–11. Available online: https://czasopisma.up.lublin.pl/index.php/asphc/article/view/2738 (accessed on 8 August 2019).
- Ammar, E.D.; Alessandro, R.T.; Hall, D.G. Ultrastructural and chemical studies on waxy secretions and wax-producing structures on the integument of the woolly oak aphid Stegophylla brevirostris Quednau (Hemiptera: Aphididae). J. Microsc. Ultrastruct. 2013, 1, 43–50. [Google Scholar] [CrossRef]
- Gorb, S.N. Functional surfaces in biology. Little structures with big effects. SSBM 2009, 1, 384. [Google Scholar] [CrossRef]
- Durak, R.; Węgrzyn, E.; Leniowski, K. When a little means a lot—Slight daily cleaning is crucial for obligatory ant-tended aphids. Ethol. Ecol. Evol. 2014, 28, 20–29. [Google Scholar] [CrossRef]
- Durak, R.; Dampc, J.; Kula-Maximenko, M.; Mołoń, M.; Durak, T. Changes in antioxidative, oxidoreductive and detoxification enzymes during development of aphids and temperature increase. Antioxidants 2021, 10, 1181. [Google Scholar] [CrossRef]
- Saeidi, F.; Mikani, A.; Moharramipour, S. Thermal tolerance variations and physiological adjustments in a winter active and a summer active aphid species. J. Therm. Biol. 2021, 98, 102950. [Google Scholar] [CrossRef]
- Durak, T.; Depciuch, J. Effect of plant sample preparation and measuring methods on ATR-FTIR spectra results. Environ. Exp. Bot. 2020, 169, 103915. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ATR-FTIR Range (cm−1) | Peak at Corresponding Wavenumber (cm−1) | Proposed Definition of the Spectral Assignments |
|---|---|---|
| 2950–2960 | 2958 | C-H stretching, asymmetric vibrations: (CH3) mainly from lipids |
| 2910–2930 | 2913 | C-H stretching, asymmetric vibrations: (CH2) mainly from lipids |
| 2835–2860 | 2849 | C-H stretching, symmetric vibrations: (CH2) mainly from lipids |
| 1720–1750 | 1737 | C=O stretching, symmetric vibrations: mainly from lipids |
| 1620–1640 | 1626 | C=O stretching vibrations of amide I: mainly from proteins |
| 1500–1560 | 1545 | C=N stretching vibrations of amide II and N-H bending vibrations: mainly from proteins |
| 1390–1420 | 1408 | C-H bending, asymmetric vibrations: (CH3) mainly from lipids |
| 1230–1260 | 1248 | O-H and C-H bending vibrations: mainly from carbohydrates |
| 1145–1180 | 1173 | C-O stretching vibrations: (C-OH, C-O-C) mainly from carbohydrates |
| ATR-FTIR Peak at Corresponding Wavenumber (cm−1) | 2958 | 2913 | 2849 | 1737 | 1626 | 1545 | 1408 | 1248 | 1173 |
|---|---|---|---|---|---|---|---|---|---|
| LDA 1 | −0.58 *** | −0.58 *** | −0.56 *** | −0.47 *** | −0.12 | −0.43 *** | −0.69 *** | −0.31 * | −0.52 *** |
| LDA 2 | 0.55 *** | 0.50 *** | 0.52 *** | 0.64 *** | 0.32 * | −0.09 | −0.08 | 0.72 *** | 0.64 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durak, R.; Ciak, B.; Durak, T. Highly Efficient Use of Infrared Spectroscopy (ATR-FTIR) to Identify Aphid Species. Biology 2022, 11, 1232. https://doi.org/10.3390/biology11081232
Durak R, Ciak B, Durak T. Highly Efficient Use of Infrared Spectroscopy (ATR-FTIR) to Identify Aphid Species. Biology. 2022; 11(8):1232. https://doi.org/10.3390/biology11081232
Chicago/Turabian StyleDurak, Roma, Beata Ciak, and Tomasz Durak. 2022. "Highly Efficient Use of Infrared Spectroscopy (ATR-FTIR) to Identify Aphid Species" Biology 11, no. 8: 1232. https://doi.org/10.3390/biology11081232