
Trans- and Within-Generational Developmental Plasticity May Benefit the Prey but Not Its Predator during Heat Waves


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
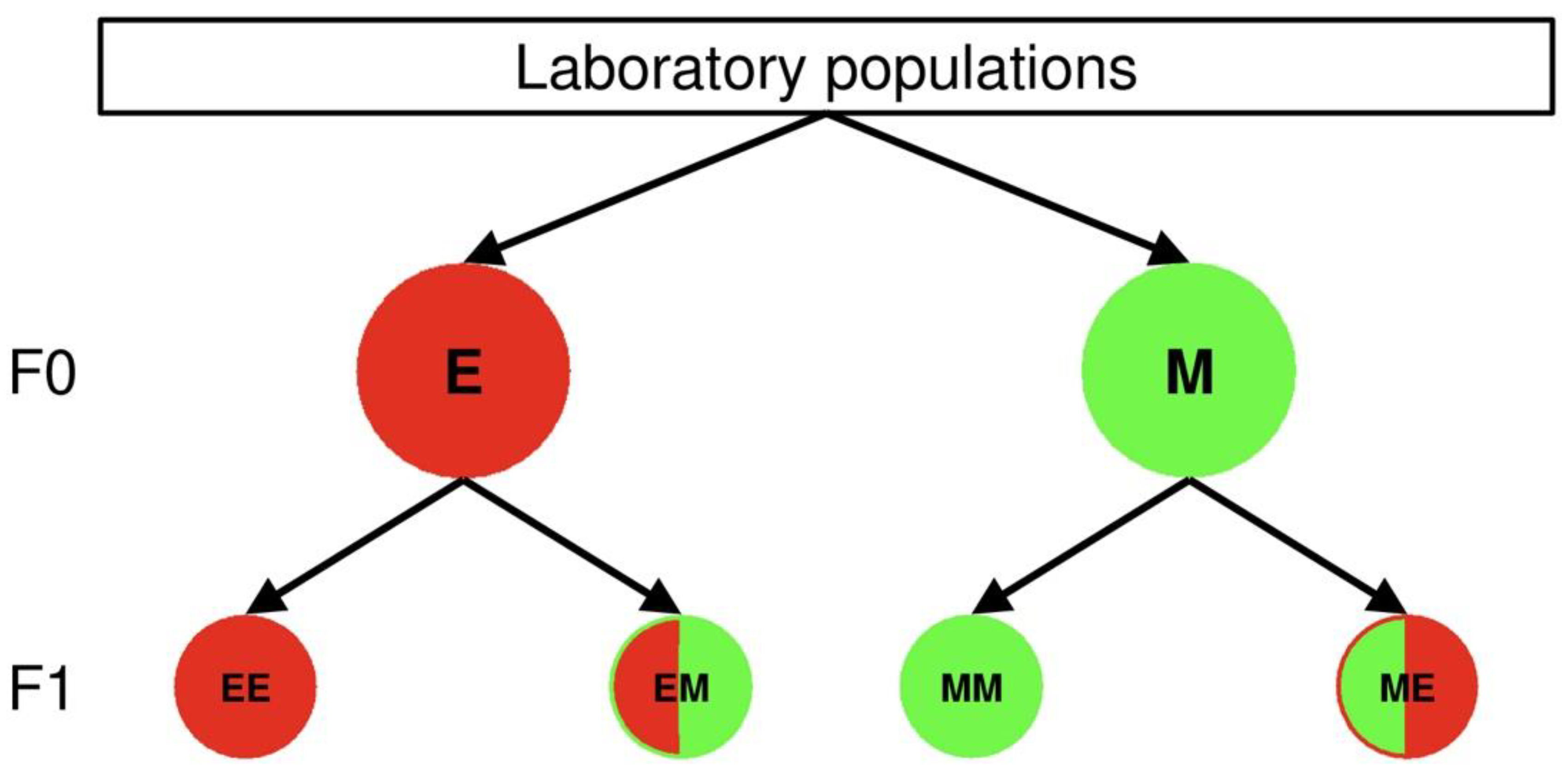
2.1. Mite Origin and Rearing
2.2. Heat Wave Conditions
2.3. Experimental Setup
2.4. Evaluated Traits
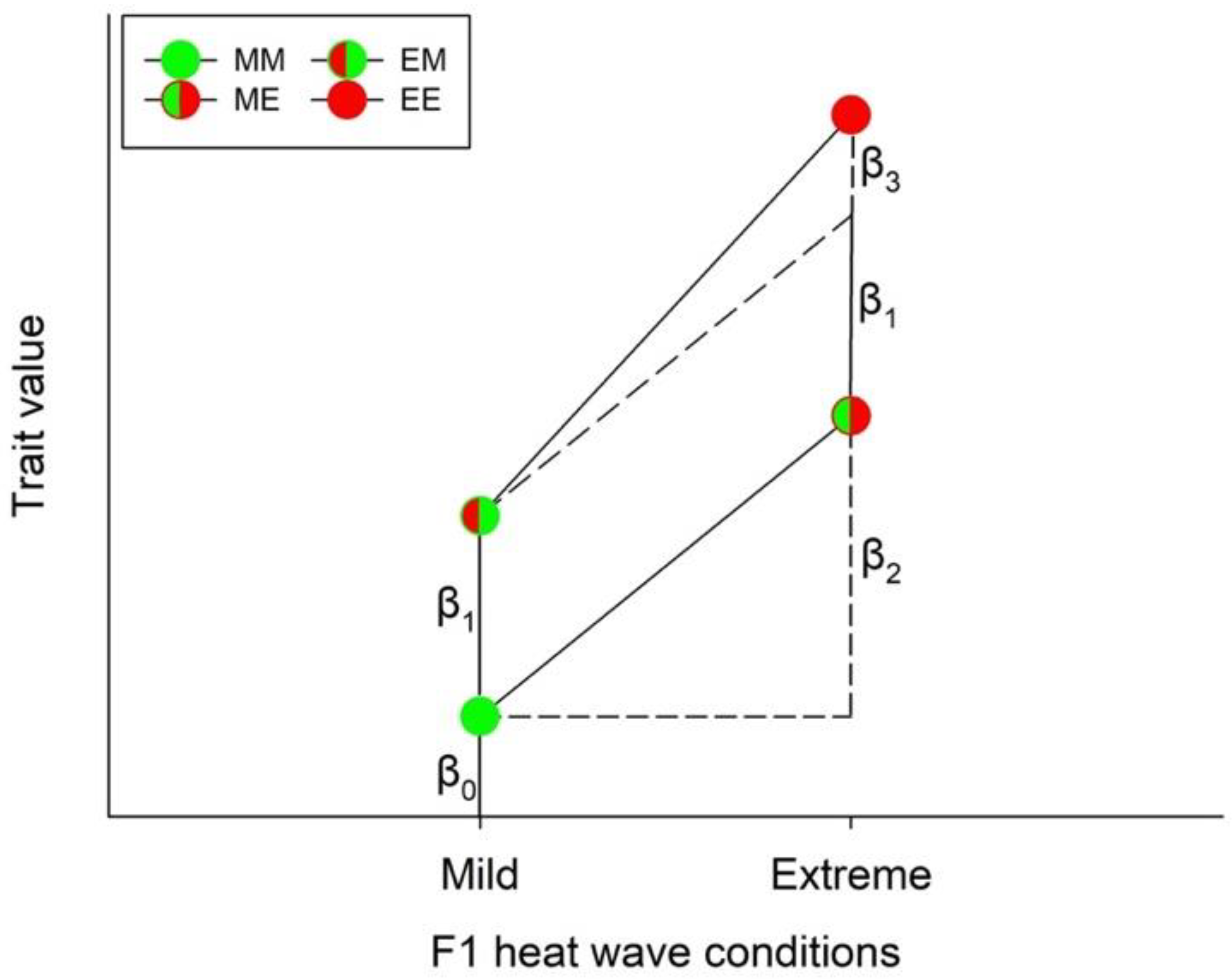
2.5. Statistical Analyses
3. Results
3.1. Heat Wave Effects on Survival and Sex Ratios
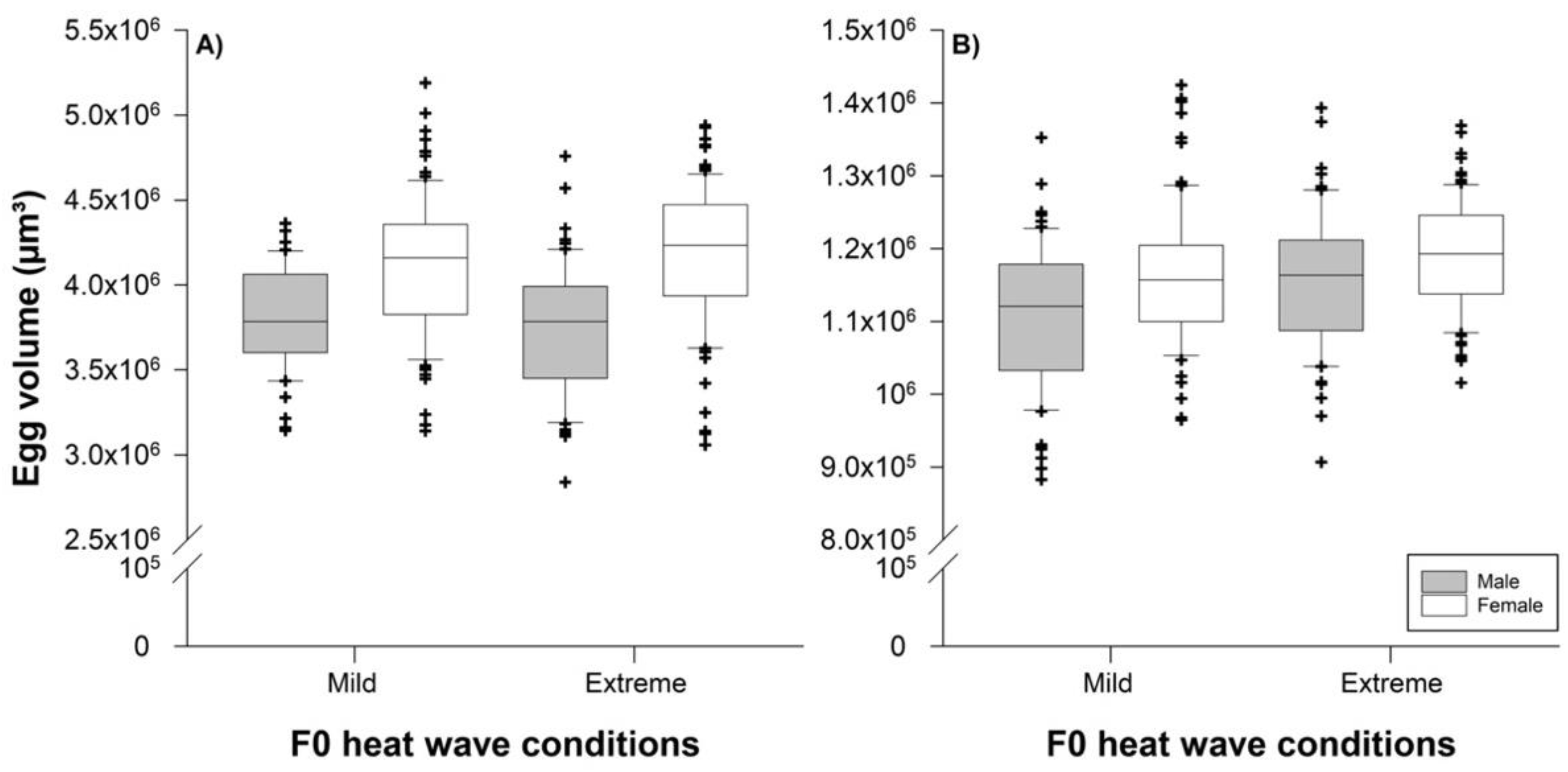
3.2. Effects of Parental Heat Wave Conditions on Offspring Egg Sizes
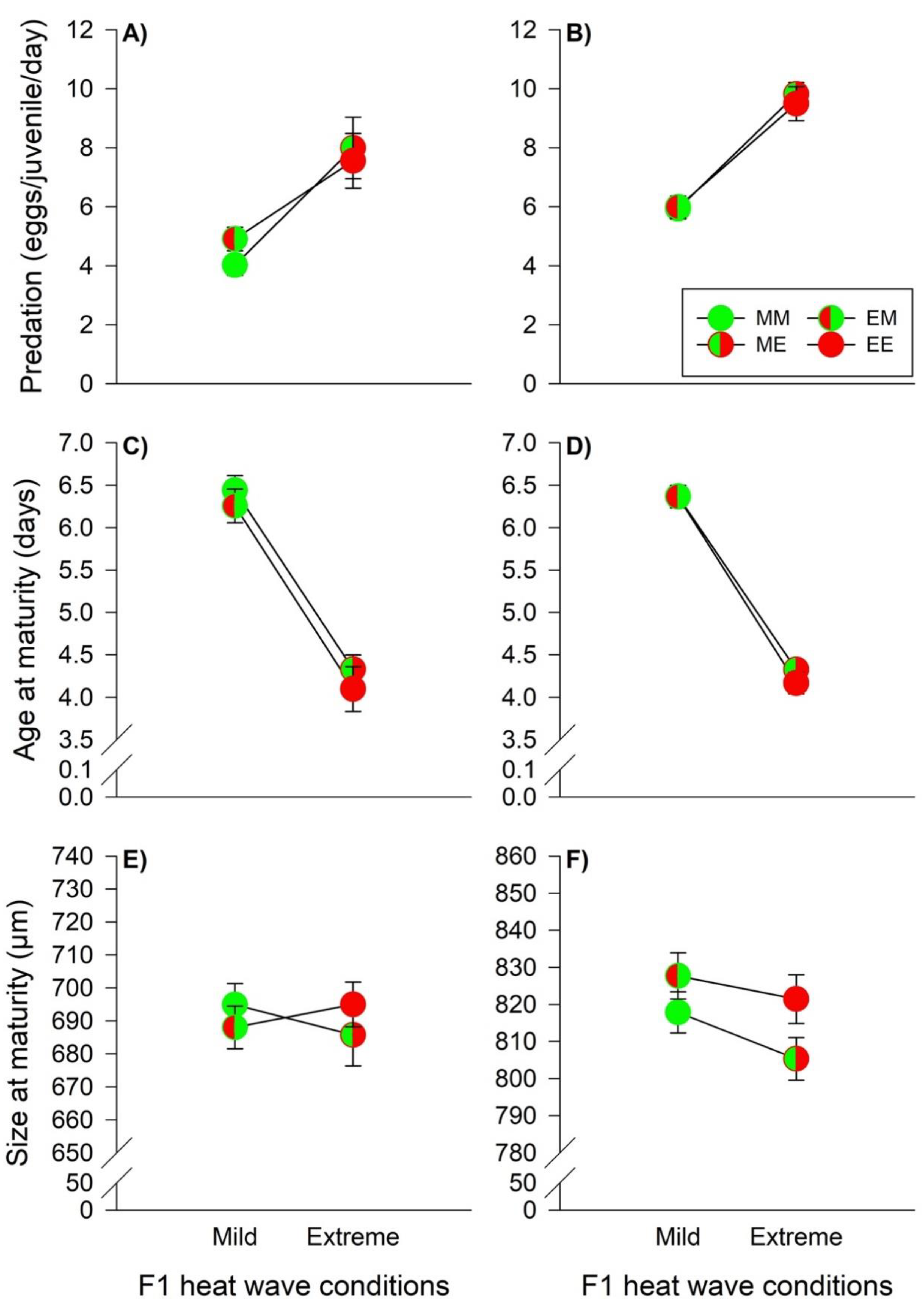
3.3. Heat Wave Effects on Offspring Food Consumption, Age and Size at Maturity
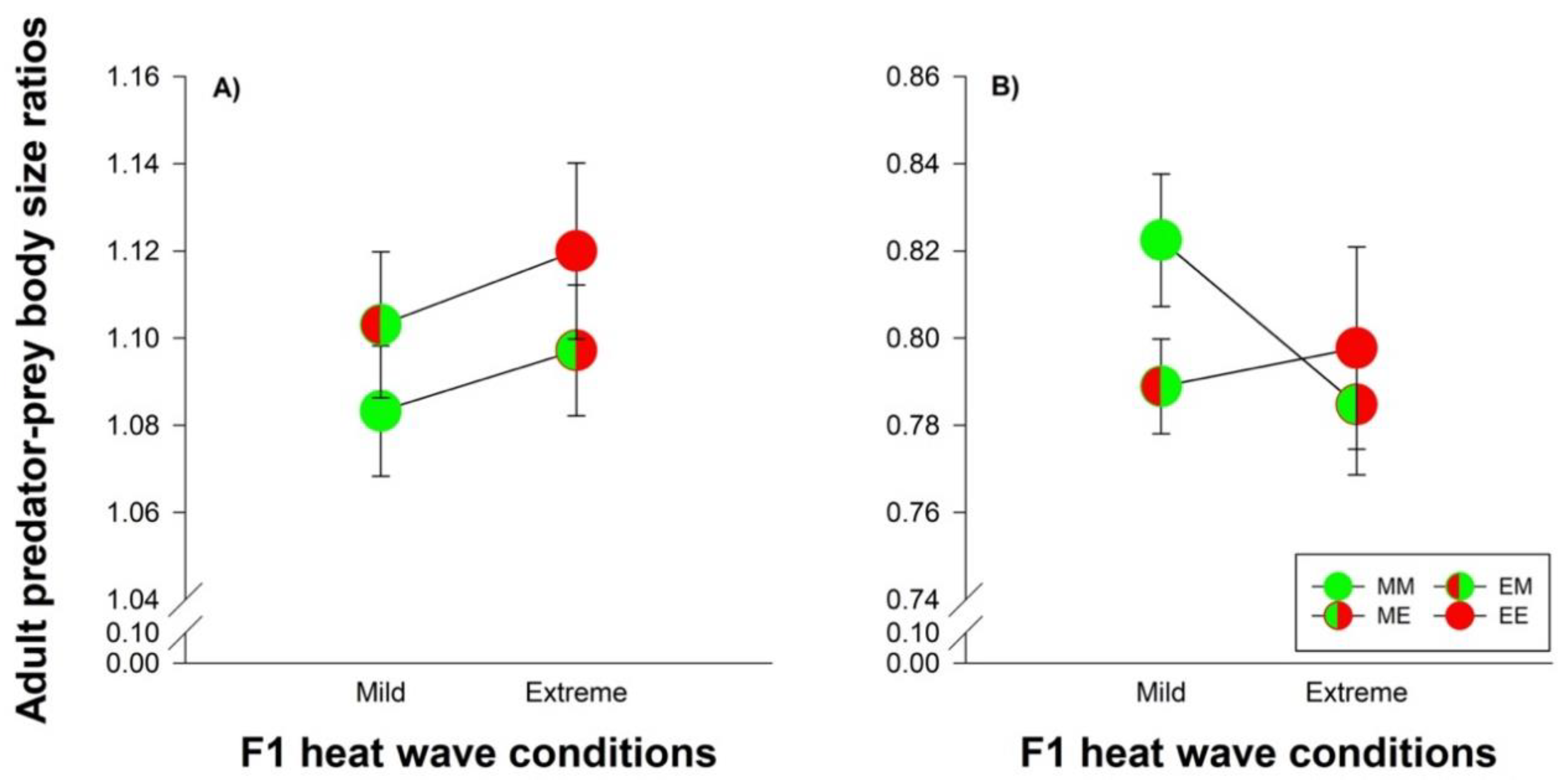
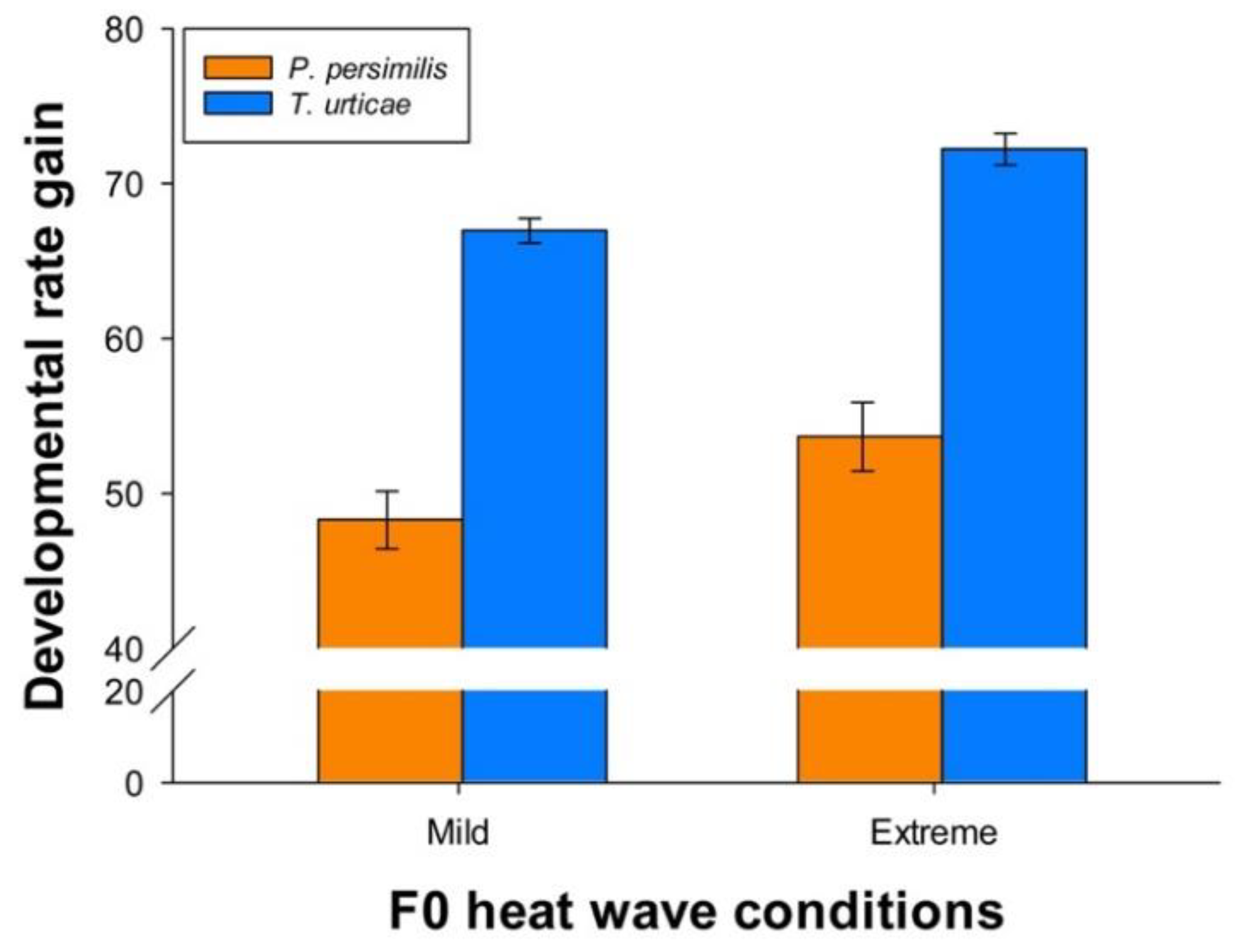
3.4. Species Comparisons: Heat Wave Effects on Predator-Prey Body Size Ratios and Developmental Time
4. Discussion
4.1. Thermal Developmental Plasticity on Age and Size at Maturity: Adaptive or Not?
4.2. Species-and Sex-Specific Responses to Heat Waves
4.3. Potential Experimental Shortcomings
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, M.; Knutti, R.; Arblaster, J.; Dufresne, J.-L.; Fichefet, T.; Friedlingstein, P.; Gao, X.; Gutowski, W.J.; Johns, T.; Krinner, G.; et al. Long-Term Climate Change: Projections, Commitments and Irreversibility. In Climate Change 2013: The Physical Science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Doschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; pp. 1029–1136. [Google Scholar]
- Meehl, G.A.; Tebaldi, C. More intense, more frequent, and longer lasting heat waves in the 21st century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingsolver, J.G.; Diamond, S.E.; Buckley, L.B. Heat stress and the fitness consequences of climate change for terrestrial ectotherms. Funct. Ecol. 2013, 27, 1415–1423. [Google Scholar] [CrossRef]
- Walzer, A.; Formayer, H.; Tixier, M.S. Evidence of trans-generational developmental modifications induced by simulated heat waves in an arthropod. Sci. Rep. 2020, 10, 4098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, N.M.; Crespel, A. Within- and trans-generational environmental adaptation to climate change: Perspectives and new challenges. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Dayananda, B.; Murray, B.R.; Webb, J.K. Hotter nests produce hatchling lizards with lower thermal tolerance. J. Exp. Biol. 2017, 220, 2159–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glass, J.R.; Stahlschmidt, Z.R. Should I stay or should I go? Complex environments influence the developmental plasticity of flight capacity and flight-related trade-offs. Biol. J. Linn. Soc. 2019, 128, 59–69. [Google Scholar] [CrossRef]
- Spinks, R.K.; Munday, P.L.; Donelson, J.M. Developmental effects of heatwave conditions on the early life stages of a coral reef fish. J. Exp. Biol. 2019, 222, jeb202713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunderson, A.R.; Fargevieille, A.; Warner, D.A. Egg incubation temperature does not influence adult heat tolerance in the lizard Anolis sagrei. Biol. Lett. 2020, 16, 20190716. [Google Scholar] [CrossRef]
- Tscholl, T.; Nachman, G.; Spangl, B.; Walzer, A. Heat waves affect prey and predators differently via developmental plasticity: Who may benefit most from global warming? Pest Manag. Sci. 2022, 78, 1099–1109. [Google Scholar] [CrossRef]
- Donelson, J.M.; Salinas, S.; Munday, P.L.; Shama, L.N. Transgenerational plasticity and climate change experiments: Where do we go from here? Glob. Chang. Biol. 2018, 24, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Bonduriansky, R.; Crean, A.J.; Day, T. The implications of nongenetic inheritance for evolution in changing environments. Evol. Appl. 2012, 5, 192–201. [Google Scholar] [CrossRef]
- Uller, T. Developmental plasticity and the evolution of parental effects. Trends Ecol. Evol. 2008, 23, 432–438. [Google Scholar] [CrossRef]
- Murren, C.J.; Auld, J.R.; Callahan, H.; Ghalambor, C.K.; Handelsman, C.A.; Heskel, M.A.; Kingsolver, J.G.; Maclean, H.J.; Masel, J.; Maughan, H.; et al. Constraints on the evolution of phenotypic plasticity: Limits and costs of phenotype and plasticity. Heredity (Edinb) 2015, 115, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonka, E.; Oborny, B.; Molnar, I.; Kisdi, E.; Hofbauer, J.; Czaran, T. The adaptive advantage of phenotypic memory in changing environments. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1995, 350, 133–141. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, T.J.; Scheiner, S.M. Phenotypic Plasticity-Functional and Conceptual Approaches; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Shine, R.; Sun, L.X. Attack strategy of an ambush predator: Which attributes of the prey trigger a pit-viper’s strike? Funct. Ecol. 2003, 17, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Nisembaum, L.G.; Besseau, L.; Paulin, C.-H.; Charpantier, A.; Martin, P.; Magnanou, E.; Fuentès, M.; Delgado, M.-J.; Falcón, J. In the heat of the night: Thermo-TRPV channels in the salmonid pineal photoreceptors and modulation of melatonin secretion. Endocrinology 2015, 156, 4629–4638. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, H.; Schmitz, A. Thermoreception: Invertebrates. In Encyclopedia of Animal Behaviour; Breed, M.D., Moore, J., Eds.; Academic Press: Oxford, UK, 2010; pp. 401–408. [Google Scholar]
- Donelan, S.C.; Hellmann, J.K.; Bell, A.M.; Luttbeg, B.; Orrock, J.L.; Sheriff, M.J.; Sih, A. Transgenerational plasticity in human-altered environments. Trends Ecol. Evol. 2020, 35, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.C.; Marshall, D.J. Adaptive parental effects: The importance of estimating environmental predictability and offspring fitness appropriately. Oikos 2014, 123, 769–776. [Google Scholar] [CrossRef]
- Vazquez, D.P.; Gianoli, E.; Morris, W.F.; Bozinovic, F. Ecological and evolutionary impacts of changing climatic variability. Biol. Rev. Camb. Philos. Soc. 2017, 92, 22–42. [Google Scholar] [CrossRef] [Green Version]
- Jeffs, C.T.; Leather, S.R. Effects of extreme, fluctuating temperature events on life history traits of the grain aphid, Sitobion avenae. Entomol. Exp. Appl. 2014, 150, 240–249. [Google Scholar] [CrossRef]
- Xing, K.; Sun, D.; Zhao, F. Within- and trans-generational life history responses to diurnal temperature amplitudes of the pupal stage in the diamondback moth. Environ. Entomol. 2021, 50, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Dinh, K.V.; Doan, K.L.U.; Doan, N.X.; Pham, H.Q.; Le, T.H.O.; Le, M.-H.; Vu, M.T.T.; Dahms, H.-U.; Truong, K.N. Parental exposures increase the vulnerability of copepod offspring to copper and a simulated marine heatwave. Environ. Pollut. 2021, 287, 117603. [Google Scholar] [CrossRef]
- Uthicke, S.; Patel, F.; Petrik, C.; Watson, S.A.; Karelitz, S.E.; Lamare, M.D. Cross-generational response of a tropical sea urchin to global change and a selection event in a 43-month mesocosm study. Glob. Chang. Biol. 2021, 27, 3448–3462. [Google Scholar] [CrossRef] [PubMed]
- Tariel, J.; Plénet, S.; Luquet, É. Transgenerational plasticity in the context of predator-prey interactions. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef]
- Gibert, J.P.; Delong, J.P. Temperature alters food web body-size structure. Biol. Lett. 2014, 10, 20140473. [Google Scholar] [CrossRef] [PubMed]
- Emmerson, M.C.; Raffaelli, D. Predator-prey body size, interaction strength and the stability of a real food web. J. Anim. Ecol. 2004, 73, 399–409. [Google Scholar] [CrossRef]
- Knapp, M.; van Houten, Y.M.; van Baal, E.; Groot, T. Use of predatory mites in commercial biocontrol: Current status and future prospects. Acarologia 2018, 58, 72–82. [Google Scholar] [CrossRef]
- Walzer, A.; Schausberger, P. Sex-specific developmental plasticity of generalist and specialist predatory mites (Acari: Phytoseiidae) in response to food stress. Biol. J. Linn. Soc. 2011, 102, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Walzer, A.; Schausberger, P. Intra- and trans-generational costs of reduced female body size caused by food limitation early in life in mites. PLoS ONE 2013, 8, e79089. [Google Scholar] [CrossRef]
- Walzer, A.; Schausberger, P. Food stress causes sex-specific maternal effects in mites. J. Exp. Biol. 2015, 218, 2603–2609. [Google Scholar] [CrossRef] [Green Version]
- Walzer, A.; Schausberger, P. Phenotypic plasticity in anti-intraguild predator strategies: Mite larvae adjust their behaviours according to vulnerability and predation risk. Exp. Appl. Acarol. 2013, 60, 95–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.Y.; Zhang, Z.Q. Development, lifespan and reproduction of spider mites exposed to predator-induced stress across generations. Biogerontology 2019, 20, 871–882. [Google Scholar] [CrossRef]
- Seiter, M.; Schausberger, P. Maternal intraguild predation risk affects offspring anti-predator behavior and learning in mites. Sci. Rep. 2015, 5, 15046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freinschlag, J.; Schausberger, P. Predation risk-mediated maternal effects in the two-spotted spider mite, Tetranychus urticae. Exp. Appl. Acarol. 2016, 69, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Vangansbeke, D.; De Schrijver, L.; Spranghers, T.; Audenaert, J.; Verhoeven, R.; Nguyen, D.T.; Gobin, B.; Tirry, L.; De Clercq, P. Alternating temperatures affect life table parameters of Phytoseiulus persimilis, Neoseiulus californicus (Acari: Phytoseiidae) and their prey Tetranychus urticae (Acari: Tetranychidae). Exp. Appl. Acarol. 2013, 61, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Vangansbeke, D.; Audenaert, J.; Nguyen, D.T.; Verhoeven, R.; Gobin, B.; Tirry, L.; De Clercq, P. Diurnal temperature variations affect development of a herbivorous arthropod pest and its predators. PLoS ONE 2015, 10, e0124898. [Google Scholar] [CrossRef] [Green Version]
- Margolies, D.C.; Wrensch, D.L. Temperature-induced changes in spider mite fitness offsetting effects of development time, fecundity, and sex ratio. Entomol. Exp. Appl. 1996, 78, 111–118. [Google Scholar] [CrossRef]
- Ubara, M.; Osakabe, M. Suspension of egg hatching caused by high humidity and submergence in spider mites. Environ. Entomol. 2015, 44, 1210–1219. [Google Scholar] [CrossRef]
- Le Hesran, S.; Groot, T.; Knapp, M.; Bukovinszky, T.; Forestier, T.; Dicke, M. Phenotypic variation in egg survival in the predatory mite Phytoseiulus persimilis under dry conditions. Biol. Control 2019, 130, 88–94. [Google Scholar] [CrossRef]
- Le Hesran, S.; Groot, T.; Knapp, M.; Bukovinszky, T.; Nugroho, J.E.; Beretta, G.; Dicke, M. Maternal effect determines drought resistance of eggs in the predatory mite Phytoseiulus persimilis. Oecologia 2020, 192, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Hosmer, D.W.; Lemeshow, S. Goodness of fit tests for the multiple logistic regression model. Commun. Stat. Theory Methods 1980, 9, 1043–1069. [Google Scholar] [CrossRef]
- Colquhoun, D. Lectures on Biostatistics. An Introduction to Statistics with Applications in Biology and Medicine; Clarendon Press: Oxford, UK, 1971; p. 425. [Google Scholar]
- Vucic-Pestic, O.; Ehnes, R.B.; Rall, B.C.; Brose, U. Warming up the system: Higher predator feeding rates but lower energetic efficiencies. Glob. Chang. Biol. 2011, 17, 1301–1310. [Google Scholar] [CrossRef]
- Rosenblatt, A.E.; Smith-Ramesh, L.M.; Schmitz, O.J. Interactive effects of multiple climate change variables on food web dynamics: Modeling the effects of changing temperature, CO2, and water availability on a tri-trophic food web. Food Webs 2017, 13, 98–108. [Google Scholar] [CrossRef]
- Caswell, H. Life history theory and the equilibrium status of populations. Am. Nat. 1982, 120, 317–339. [Google Scholar] [CrossRef]
- Sabelis, M.W. Reproduction. In Spider Mites-Their Biology, Natural Enemies and Control-Volume 1B; Helle, W., Sabelis, M.W., Eds.; World Crop Pests; Elsevier Science Publishers B.V.: Amsterdam, The Netherlands, 1985; pp. 73–82. [Google Scholar]
- Atkinson, D. Temperature and organism size-a biological law for ectotherms? Adv. Ecol. Res. 1994, 25. [Google Scholar]
- Walters, R.J.; Hassall, M. The temperature-size rule in ectotherms: May a general explanation exist after all? Am. Nat. 2006, 167, 510–523. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Massie, K.R.; Ragland, G.J.; Smith, M.H. Rapid population divergence in thermal reaction norms for an invading species: Breaking the temperature-size rule. J. Evol. Biol. 2007, 20, 892–900. [Google Scholar] [CrossRef]
- Fu, D.M.; He, H.M.; Zou, C.; Xiao, H.J.; Xue, F.S. Life-history responses of the rice stem borer Chilo suppressalis to temperature change: Breaking the temperature-size rule. J. Therm. Biol. 2016, 61, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-P.; Chou, L.-S. Temperature effects on life history traits of two sympatric branchiopods from an ephemeral wetland. PLoS ONE 2017, 12, e0179449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Mishra, G.; Omkar. Plasticity in reproductive output and development in response to thermal variation in ladybird beetle, Menochilus sexmaculatus. J. Therm. Biol. 2018, 71, 180–188. [Google Scholar] [CrossRef]
- Xia, Q.W.; Chen, C.; Tang, J.J.; He, H.M.; Xue, F.S. A reverse temperature-size rule associated with a negative relationship between larval development time and pupal weight in a tropical population of Ostrinia furnacalis. Physiol. Entomol. 2019, 44, 209–214. [Google Scholar] [CrossRef]
- Aguilar-Alberola, J.A.; Mesquita-Joanes, F. Breaking the temperature-size rule: Thermal effects on growth, development and fecundity of a crustacean from temporary waters. J. Therm. Biol. 2014, 42, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Barrett, K.; Helms, B.S.; Samoray, S.T.; Guyer, C. Growth patterns of a stream vertebrate differ between urban and forested catchments. Freshw. Biol. 2010, 55, 1628–1635. [Google Scholar] [CrossRef]
- Angilletta, M.J., Jr. Thermal Adaptation-a Theoretical and Empirical Synthesis; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Gardner, J.L.; Heinsohn, R.; Joseph, L. Shifting latitudinal clines in avian body size correlate with global warming in Australian passerines. Proc. R. Soc. B Biol. Sci. 2009, 276, 3845–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teplitsky, C.; Millien, V. Climate warming and Bergmann’s rule through time: Is there any evidence? Evol. Appl. 2014, 7, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Baar, Y.; Friedman, A.L.L.; Meiri, S.; Scharf, I. Little effect of climate change on body size of herbivorous beetles. Insect Sci. 2018, 25, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Siepielski, A.M.; Morrissey, M.B.; Carlson, S.M.; Francis, C.D.; Kingsolver, J.G.; Whitney, K.D.; Kruuk, L.E.B. No evidence that warmer temperatures are associated with selection for smaller body sizes. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191332. [Google Scholar] [CrossRef]
- Grainger, T.N.; Levine, J.M. Rapid evolution of life-history traits in response to warming, predation and competition: A meta-analysis. Ecol. Lett. 2022, 25, 541–554. [Google Scholar] [CrossRef]
- Baudier, K.M.; Mudd, A.E.; Erickson, S.C.; O’Donnell, S. Microhabitat and body size effects on heat tolerance: Implications for responses to climate change (army ants: Formicidae, Ecitoninae). J. Anim. Ecol. 2015, 84, 1322–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendt, C.F.; Verble-Pearson, R. Critical thermal maxima and body size positively correlate in red imported fire ants, Solenopsis invicta. Southwest. Nat. 2016, 61, 79–83. [Google Scholar] [CrossRef]
- Nervo, B.; Roggero, A.; Isaia, M.; Chamberlain, D.; Rolando, A.; Palestrini, C. Integrating thermal tolerance, water balance and morphology: An experimental study on dung beetles. J. Therm. Biol. 2021, 101, 103093. [Google Scholar] [CrossRef] [PubMed]
- Esperk, T.; Kjaersgaard, A.; Walters, R.J.; Berger, D.; Blanckenhorn, W.U. Plastic and evolutionary responses to heat stress in a temperate dung fly: Negative correlation between basal and induced heat tolerance? J. Evol. Biol. 2016, 29, 900–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, A.R.; Diamond, S.E.; Martin, R.A. Evidence for the evolution of thermal tolerance, but not desiccation tolerance, in response to hotter, drier city conditions in a cosmopolitan, terrestrial isopod. Evol. Appl. 2021, 14, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Angilletta, M.J., Jr.; Dunham, A.E. The temperature-size rule in ectotherms: Simple evolutionary explanations may not be general. Am. Nat. 2003, 162, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Fox, C.W.; Czesak, M.E. Evolutionary ecology of progeny size in arthropods. Annu. Rev. Entomol. 2000, 45, 341–369. [Google Scholar] [CrossRef] [Green Version]
- Rall, B.C.; Vucic-Pestic, O.; Ehnes, R.B.; Emmerson, M.; Brose, U. Temperature, predator-prey interaction strength and population stability. Glob. Chang. Biol. 2010, 16, 2145–2157. [Google Scholar] [CrossRef]
- Stillwell, R.C.; Blanckenhorn, W.U.; Teder, T.; Davidowitz, G.; Fox, C.W. Sex differences in phenotypic plasticity affect variation in sexual size dimorphism in insects: From physiology to evolution. Annu. Rev. Entomol. 2010, 55, 227–245. [Google Scholar] [CrossRef] [Green Version]
- Stearns, S.C.; Kawecki, T.J. Fitness sensitivity and the canalization of life-history traits. Evolution 1994, 48, 1438–1450. [Google Scholar] [CrossRef]
- Schmalhausen, I.I. Factors of Evolution: The Theory of Stabilizing Selection; Blakiston: Philadelphia, PA, USA, 1949. [Google Scholar]
- Enigl, M.; Schausberger, P. Mate choice in the predaceous mite Phytoseiulus persimilis: Evidence of self-referent phenotype matching? Entomol. Exp. Appl. 2004, 112, 21–28. [Google Scholar] [CrossRef]
- Walzer, A.; Schausberger, P. Interdependent effects of male and female body size plasticity on mating behaviour of predatory mites. Anim. Behav. 2015, 100, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Walzer, A.; Schausberger, P. Canalization of body size matters for lifetime reproductive success of male predatory mites (Acari: Phytoseiidae). Biol. J. Linn. Soc. 2014, 111, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Schausberger, P.; Walzer, A.; Murata, Y.; Osakabe, M. Low level of polyandry constrains phenotypic plasticity of male body size in mites. PLoS ONE 2017, 12, e0188924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moya-Laraño, J.; Halaj, J.; Wise, D.H. Climbing to reach females: Romeo should be small. Evolution 2002, 56, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Brandt, Y.; Andrade, M.C.B. What is the matter with the gravity hypothesis? Funct. Ecol. 2007, 21, 1182–1183. [Google Scholar] [CrossRef]
- García-Navas, V.; Noguerales, V.; Cordero, P.J.; Ortego, J. Ecological drivers of body size evolution and sexual size dimorphism in short-horned grasshoppers (Orthoptera: Acrididae). J. Evol. Biol. 2017, 30, 1592–1608. [Google Scholar] [CrossRef] [PubMed]
- Kielland, O.N.; Bech, C.; Einum, S. No evidence for thermal transgenerational plasticity in metabolism when minimizing the potential for confounding effects. Can. J. Zool. 2017, 284, 20162494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uller, T.; Nakagawa, S.; English, S. Weak evidence for anticipatory parental effects in plants and animals. J. Evol. Biol. 2013, 26, 2161–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, S.C.; Marshall, D.J. Temperature-induced maternal effects and environmental predictability. J. Exp. Biol. 2011, 214, 2329–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.-Y.; Xing, K.; Zhao, F. Complex delayed and transgenerational effects driven by the interaction of heat and insecticide in the maternal generation of the wheat aphid, Sitobion avenae. Pest Manag. Sci. 2021, 77, 4453–4461. [Google Scholar] [CrossRef]
- Faassen, E.J.; García-Altares, M.; Mendes e Mello, M.; Lürling, M. Trans generational effects of the neurotoxin BMAA on the aquatic grazer Daphnia magna. Aquat. Toxicol. 2015, 168, 98–107. [Google Scholar] [CrossRef]
- Grostal, P.; Dicke, M. Direct and indirect cues of predation risk influence behavior and reproduction of prey: A case for acarine interactions. Behav. Ecol. 1999, 10, 422–427. [Google Scholar] [CrossRef]



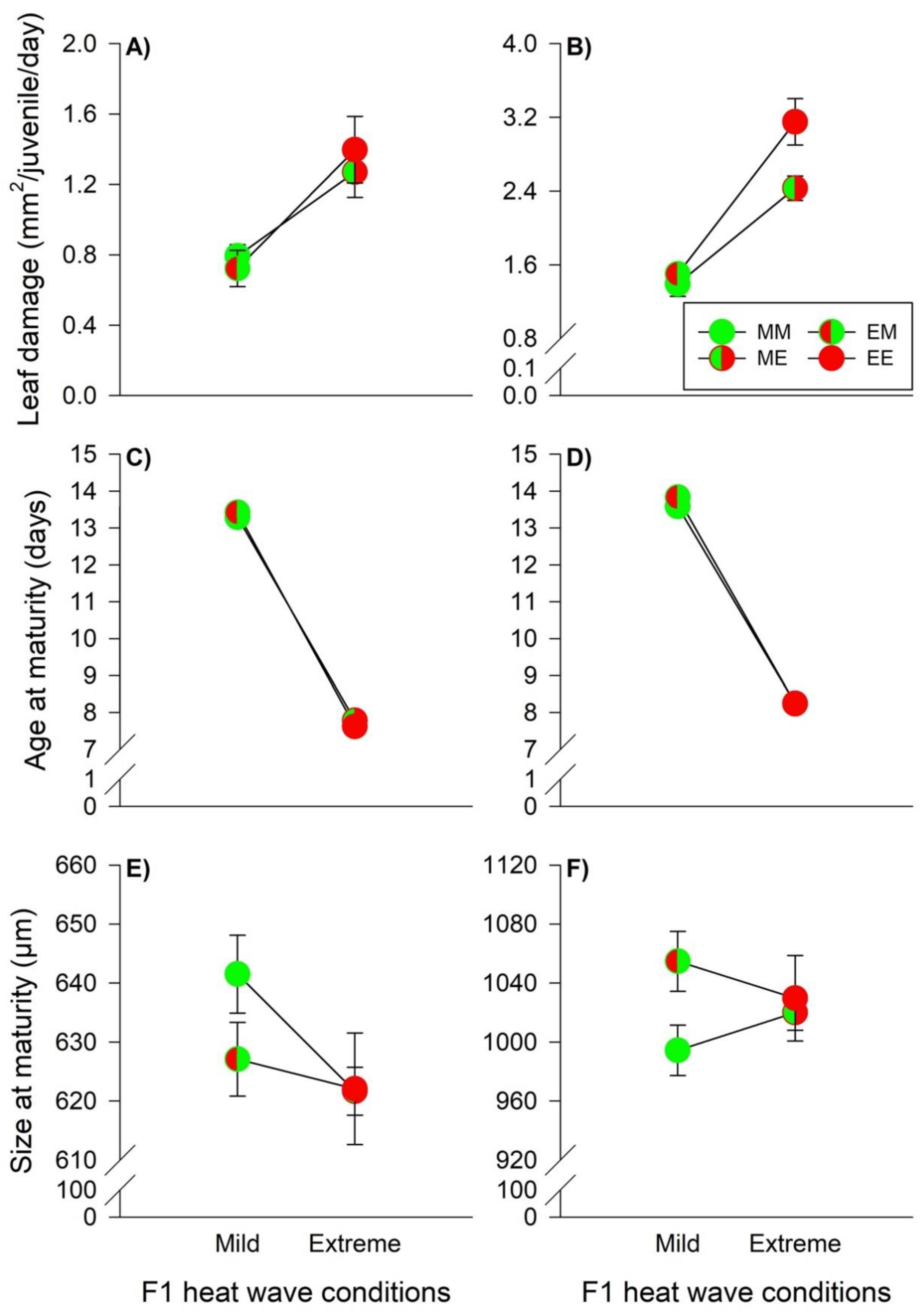
 , EM =
, EM =  , ME =
, ME =  , EE =
, EE =  ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as their parents. Vertical lines show the 95% confidence limits.
, EM = , ME = , EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as their parents. Vertical lines show the 95% confidence limits.
). The lines connecting two circles refer to offspring experiencing the same heat wave condition as their parents. Vertical lines show the 95% confidence limits.
, EM = , ME = , EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as their parents. Vertical lines show the 95% confidence limits. , EM = , ME = , EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as parents. Vertical lines show 95% confidence limits.
, EM = , ME = , EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as parents. Vertical lines show 95% confidence limits.
, EM = , ME = , EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as parents. Vertical lines show 95% confidence limits.
, EM = , ME = , EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as parents. Vertical lines show 95% confidence limits. , EM = , ME = and EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as their parents. Vertical lines show the 95% confidence limits.
, EM = , ME = and EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as their parents. Vertical lines show the 95% confidence limits.
, EM = , ME = and EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as their parents. Vertical lines show the 95% confidence limits.
, EM = , ME = and EE = ). The lines connecting two circles refer to offspring experiencing the same heat wave condition as their parents. Vertical lines show the 95% confidence limits.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Source of Variation | Phytoseiulus persimilis | Tetranychus urticae | ||||
|---|---|---|---|---|---|---|---|
| Wald’s χ2 | df | p | Wald’s χ2 | df | p | ||
| Survival | F0 | 2.076 | 1 | 0.1498 | 0.436 | 1 | 0.5090 |
| F1 | 2.770 | 1 | 0.0960 | 2.945 | 1 | 0.0862 | |
| F0*F1 | 0.316 | 1 | 0.5742 | 0.253 | 1 | 0.6149 | |
| Goodness-of fit | 0.318 | 2 | 0.8532 | 0.254 | 2 | 0.8808 | |
| Sex ratio | F0 | 0.034 | 1 | 0.8531 | 0.028 | 1 | 0.8663 |
| F1 | 2.272 | 1 | 0.1317 | 0.028 | 1 | 0.8663 | |
| F0*F1 | 0.012 | 1 | 0.9143 | 2.217 | 1 | 0.1365 | |
| Goodness-of-fit | 0.012 | 2 | 0.9942 | 2.223 | 2 | 0.3291 | |
| Factor | Source of Variation | P. persimilis Males | P. persimilis Females | ||||||
|---|---|---|---|---|---|---|---|---|---|
| t | df | p | t | df | p | ||||
| Age at maturity | F0 | −0.1848 | −1.34 | 109 | 0.1816 | −0.0040 | −0.05 | 176 | 0.9638 |
| F1 | −2.1126 | −13.68 | 109 | <0.0001 | −2.0436 | −22.86 | 176 | <0.0001 | |
| F0*F1 | −0.0447 | −0.21 | 109 | 0.8317 | −0.1543 | −1.26 | 176 | 0.2085 | |
| Size at maturity | F0 | −6.8912 | −1.44 | 109 | 0.1525 | 9.8281 | 2.21 | 176 | 0.0281 |
| F1 | −9.2020 | −1.71 | 109 | 0.0896 | −13.1326 | −2.92 | 176 | 0.0040 | |
| F0*F1 | 16.1598 | 2.21 | 109 | 0.0290 | 6.8832 | 1.12 | 176 | 0.2653 | |
| Food consumption | F0 | 0.8843 | 1.99 | 109 | 0.0489 | 0.0604 | 0.19 | 176 | 0.8503 |
| F1 | 3.9666 | 7.95 | 109 | <0.0001 | 3.8859 | 11.98 | 176 | <0.0001 | |
| F0*F1 | −1.3177 | −1.94 | 109 | 0.0544 | −0.3847 | −0.87 | 176 | 0.3869 | |
| Factor | Source of Variation | T. urticae Males | T. urticae Females | ||||||
|---|---|---|---|---|---|---|---|---|---|
| t | df | p | t | df | p | ||||
| Age at maturity | F0 | 0.1310 | 1.19 | 134 | 0.2366 | 0.2464 | 2.30 | 162 | 0.0225 |
| F1 | −5.5048 | −53.05 | 134 | <0.0001 | −5.3386 | −52.40 | 162 | <0.0001 | |
| F0*F1 | −0.2929 | −1.95 | 134 | 0.0529 | −0.2569 | −1.77 | 162 | 0.0788 | |
| Size at maturity | F0 | −14.4298 | −2.72 | 134 | 0.0073 | 60.3511 | 4.02 | 162 | <0.0001 |
| F1 | −19.8578 | −3.98 | 134 | <0.0001 | 25.6546 | 1.79 | 162 | 0.0747 | |
| F0*F1 | 14.8522 | 2.06 | 134 | 0.0414 | −50.6480 | −2.49 | 162 | 0.0137 | |
| Food consumption | F0 | −0.0708 | −0.68 | 134 | 0.4954 | 0.1062 | 0.87 | 162 | 0.3861 |
| F1 | 0.4773 | 4.89 | 134 | <0.0001 | 1.0373 | 8.92 | 162 | <0.0001 | |
| F0*F1 | 0.1983 | 1.41 | 134 | 0.1620 | 0.6146 | 3.72 | 162 | 0.0003 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walzer, A.; Nachman, G.; Spangl, B.; Stijak, M.; Tscholl, T. Trans- and Within-Generational Developmental Plasticity May Benefit the Prey but Not Its Predator during Heat Waves. Biology 2022, 11, 1123. https://doi.org/10.3390/biology11081123
Walzer A, Nachman G, Spangl B, Stijak M, Tscholl T. Trans- and Within-Generational Developmental Plasticity May Benefit the Prey but Not Its Predator during Heat Waves. Biology. 2022; 11(8):1123. https://doi.org/10.3390/biology11081123
Chicago/Turabian StyleWalzer, Andreas, Gösta Nachman, Bernhard Spangl, Miroslava Stijak, and Thomas Tscholl. 2022. "Trans- and Within-Generational Developmental Plasticity May Benefit the Prey but Not Its Predator during Heat Waves" Biology 11, no. 8: 1123. https://doi.org/10.3390/biology11081123