
Confocal Characterization of Intestinal Dendritic Cells from Myxines to Teleosts

 ,
,  , ,
, ,  , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
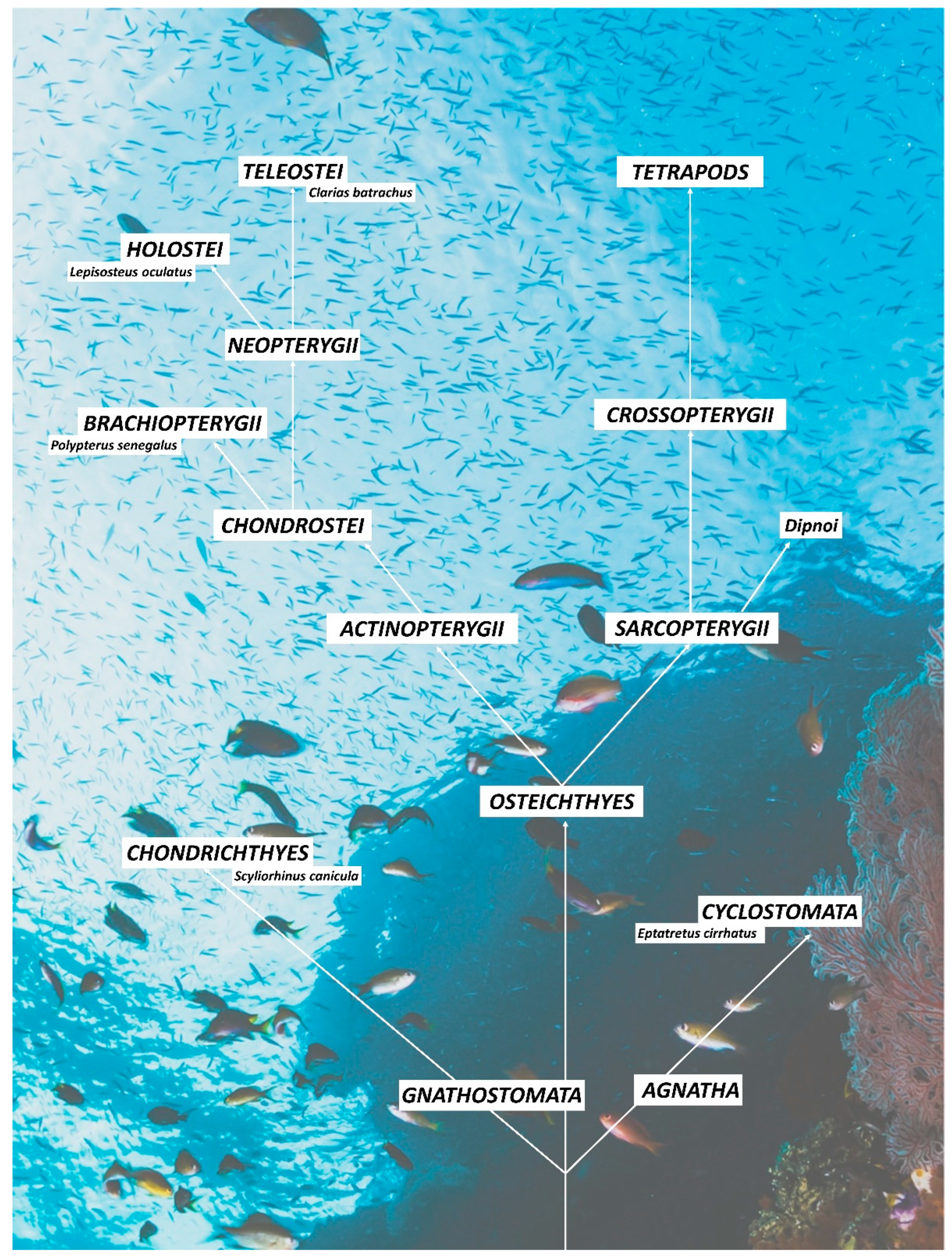
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Tissue Preparation
2.3. Histology and Histochemistry
2.4. Immunofluorescence
2.5. Laser Confocal Immunofluorescence
2.6. Statistical Analysis
2.7. Phylogenetic Conservation of Primary Antibodies
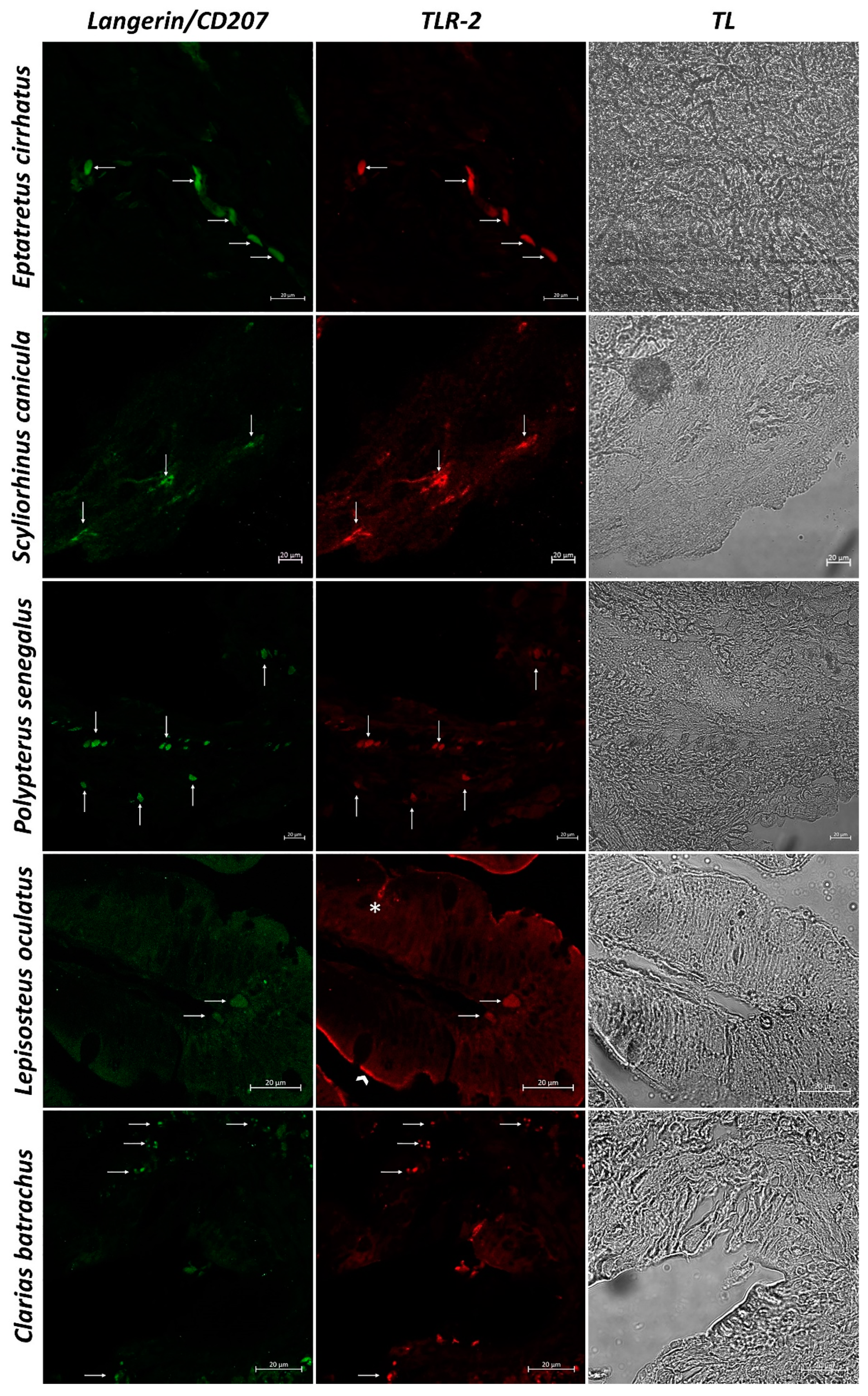
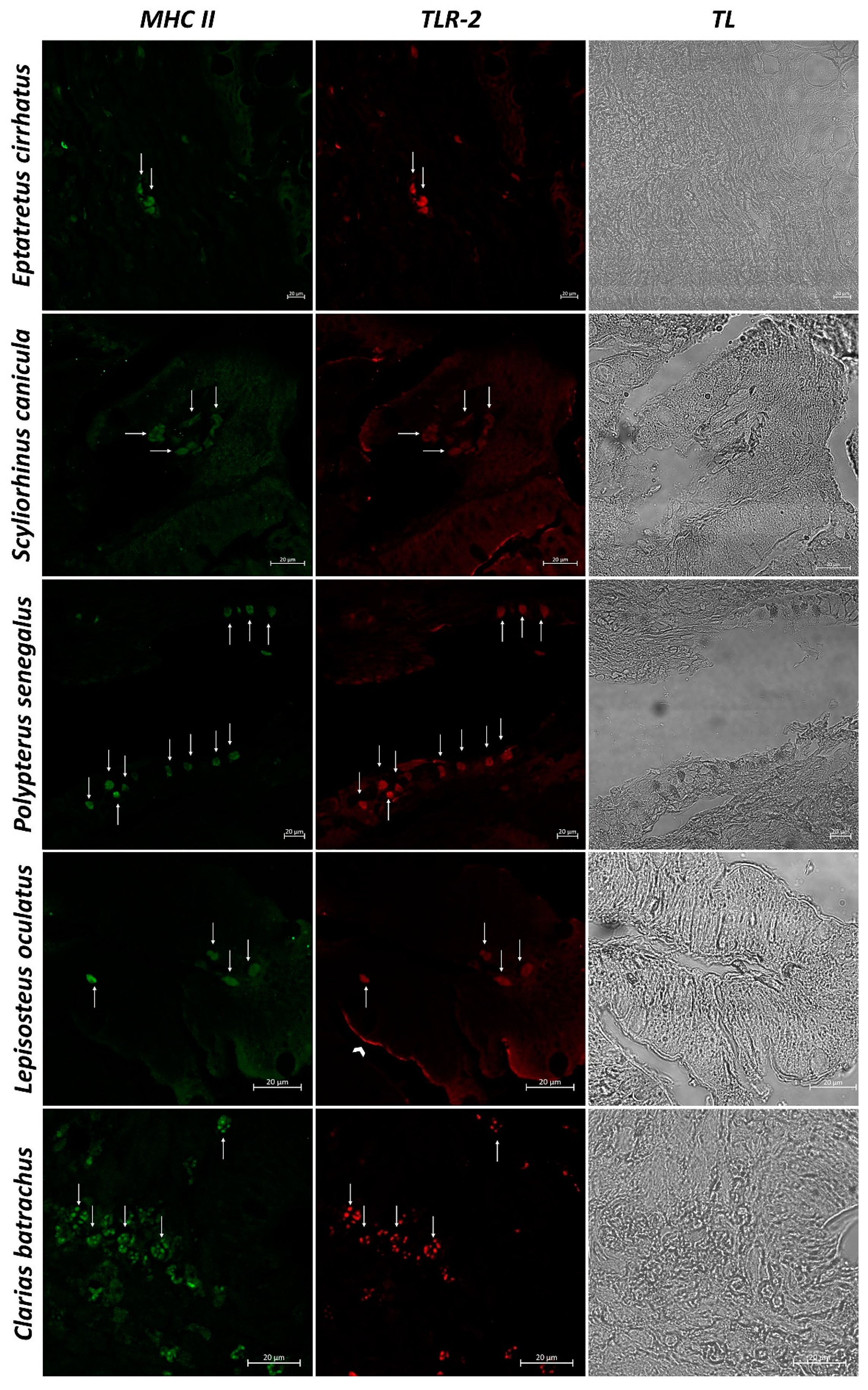
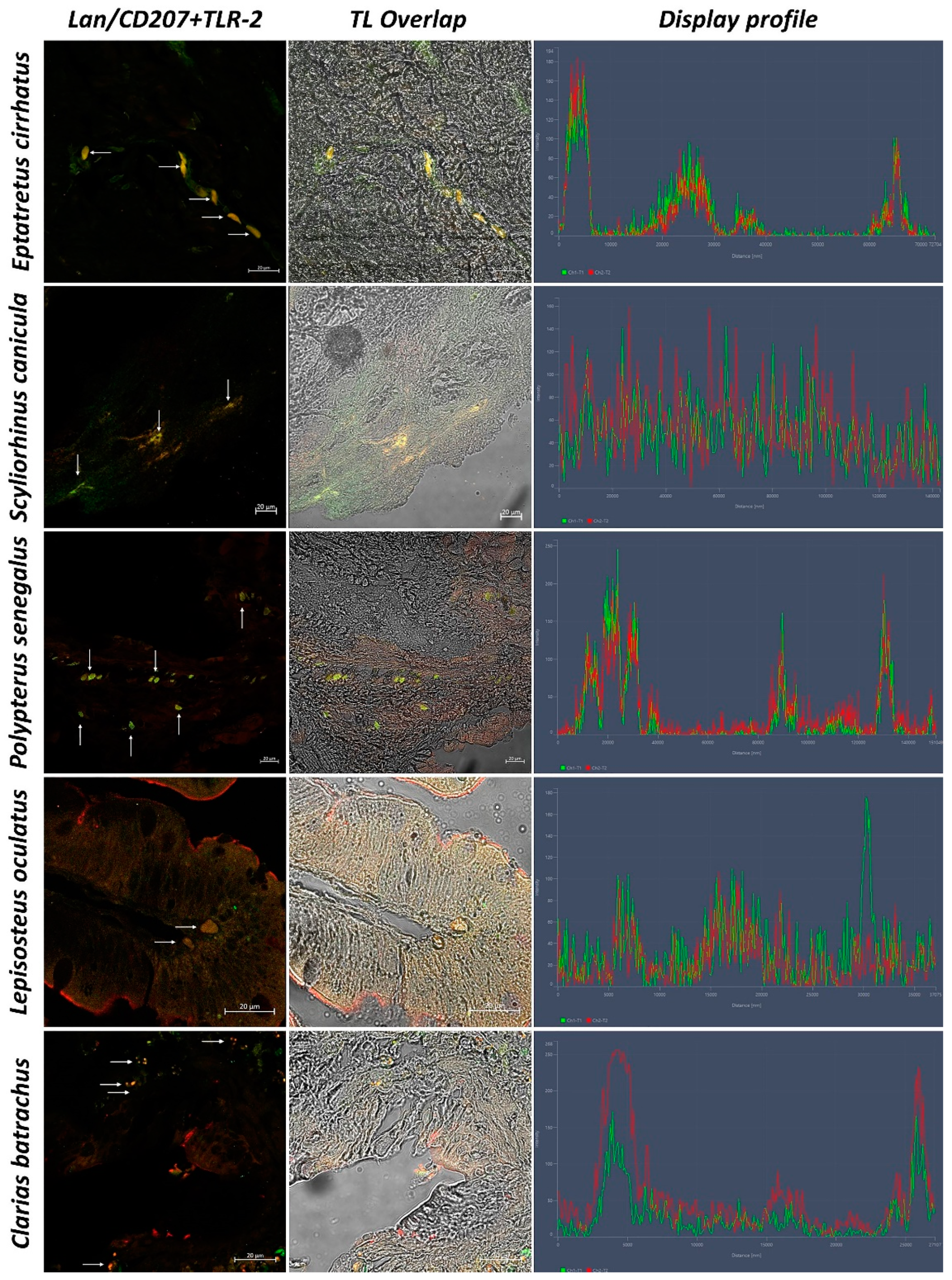
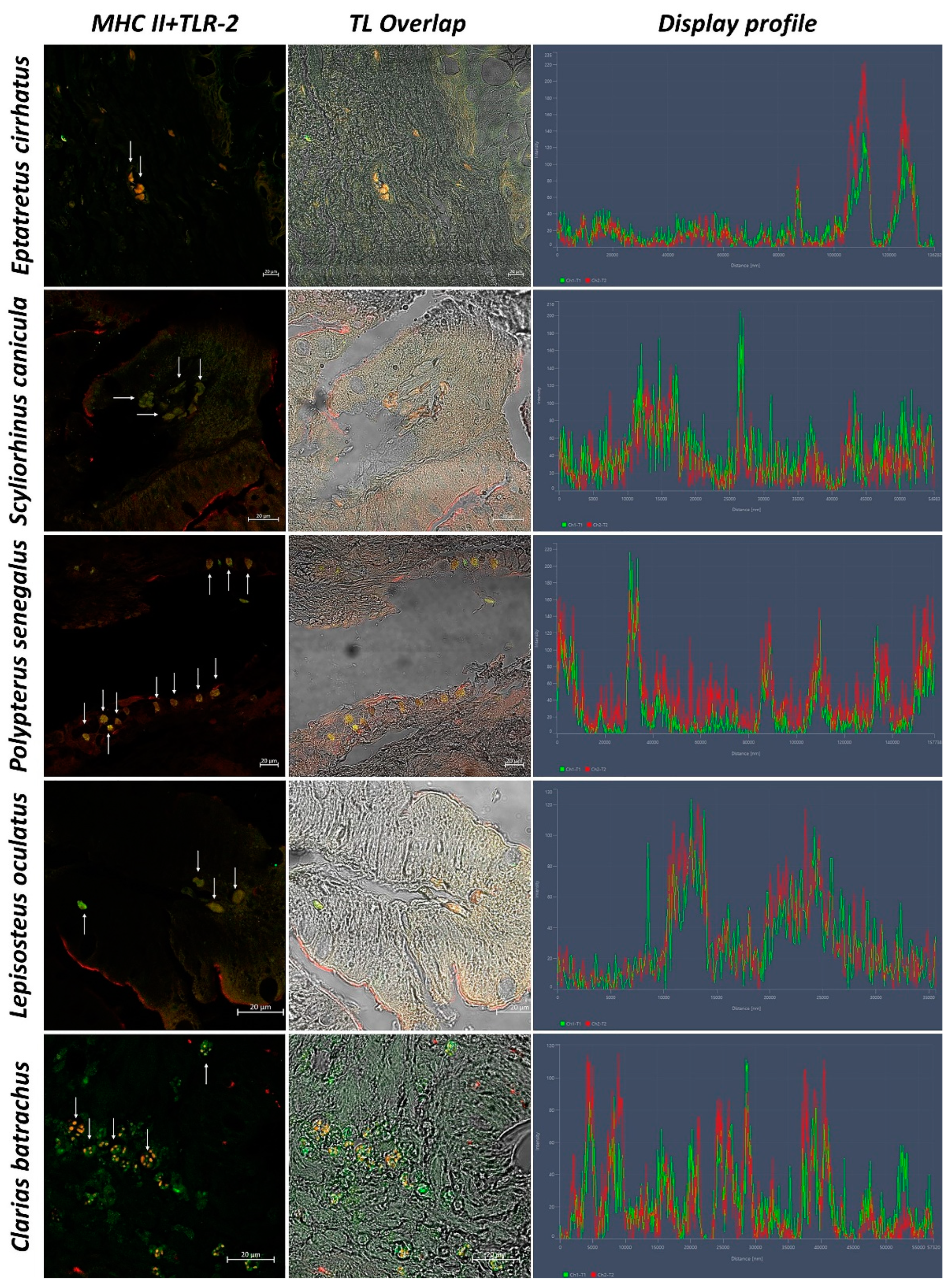
3. Results
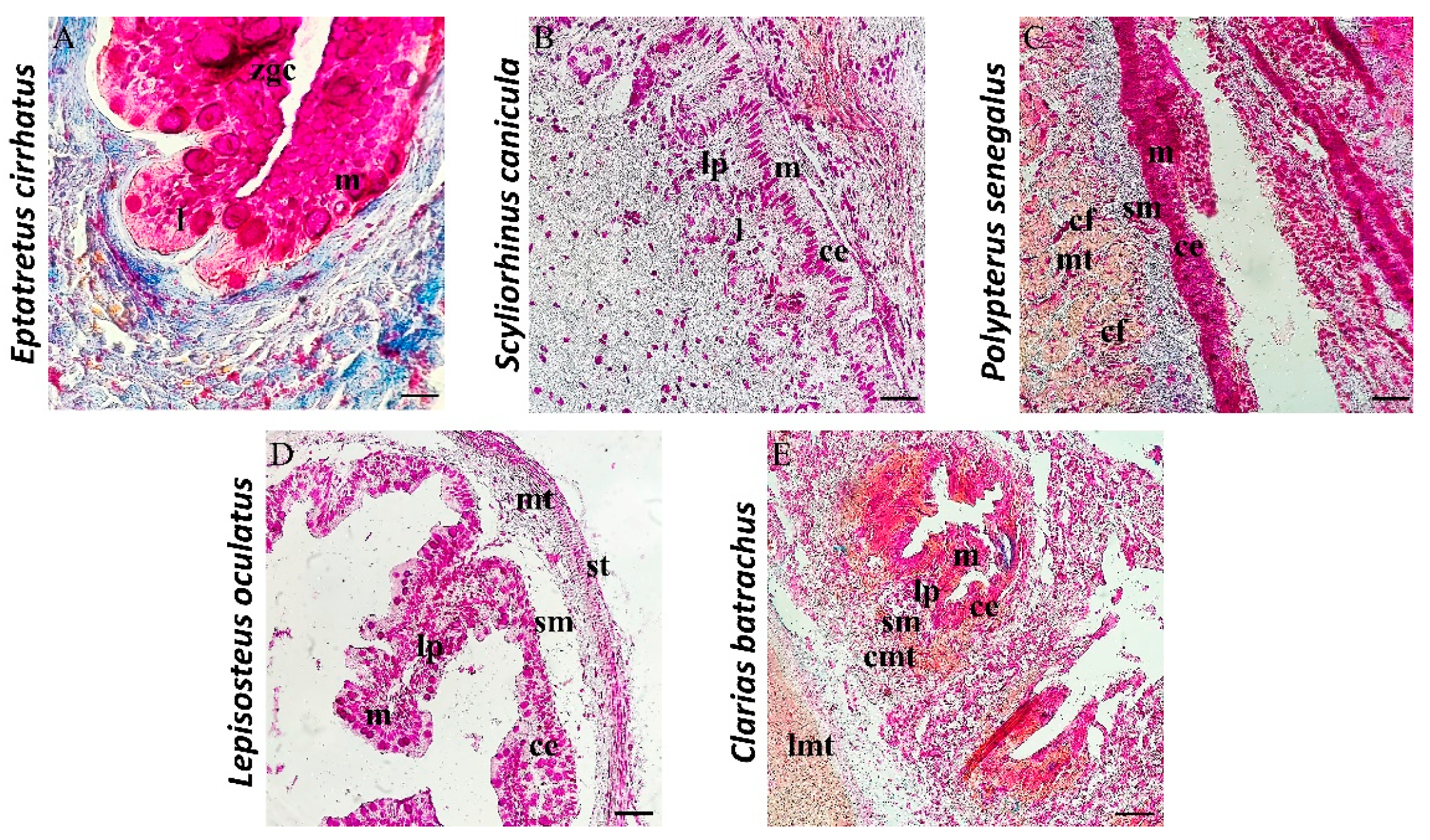
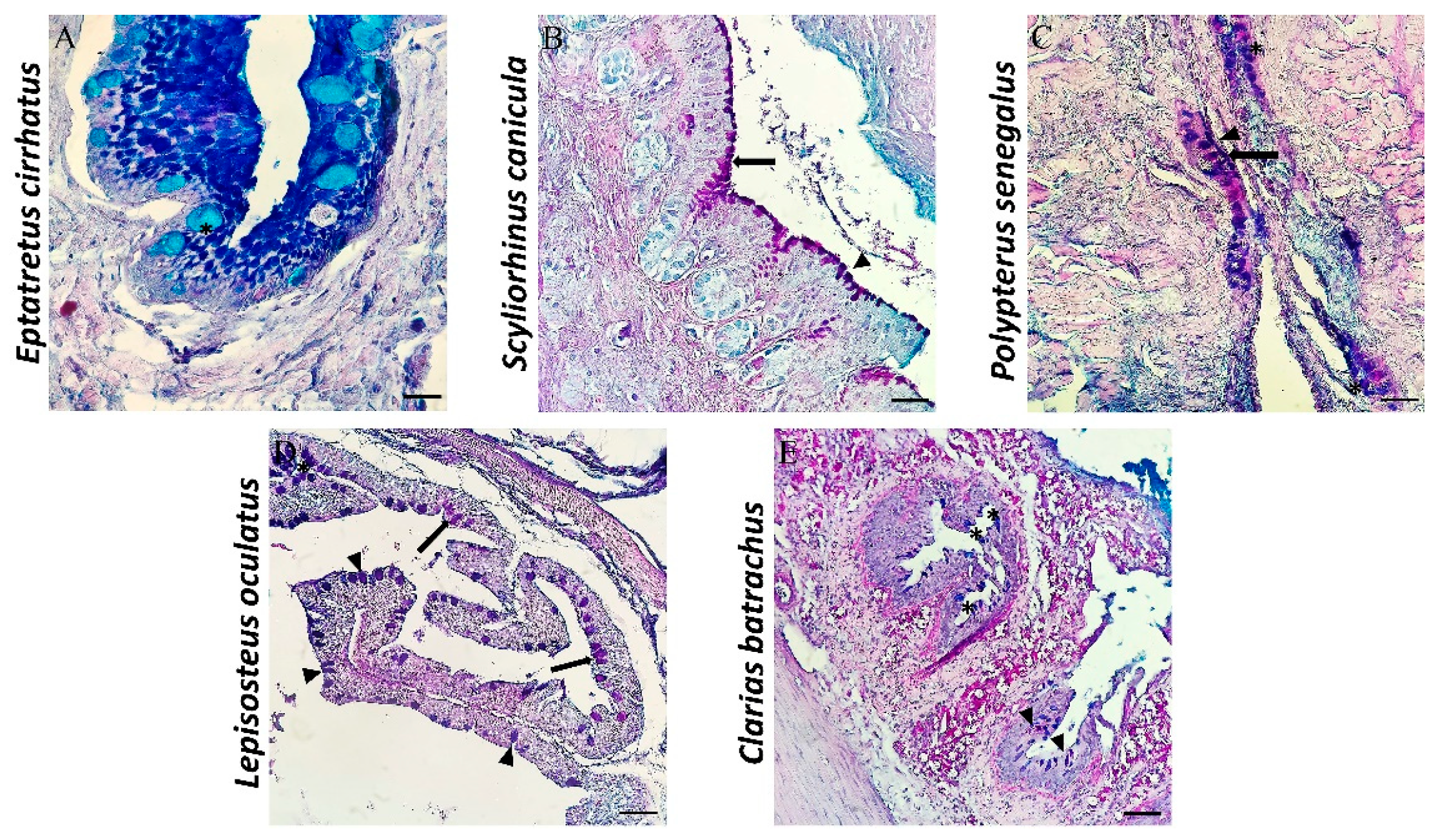
3.1. Histological and Histochemical Description
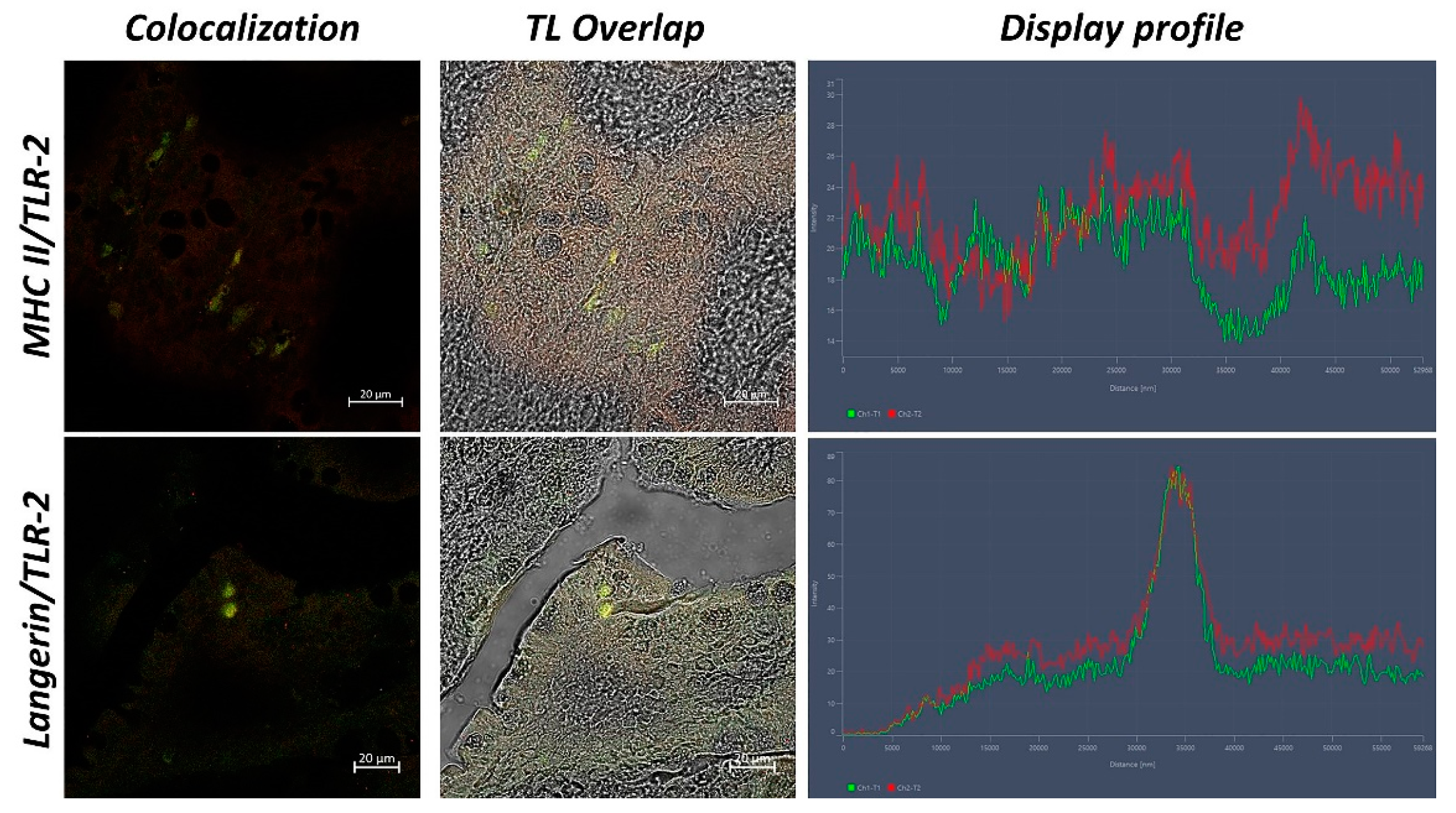
3.2. Confocal Scanning laser Microscopy
3.3. Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, L.; Qin, Z.; Liu, H.; Lin, L.; Ye, J.; Li, J. Recent Advances on Phagocytic B Cells in Teleost Fish. Front. Immunol. 2020, 11, 824. [Google Scholar] [CrossRef]
- Sunyer, J.O. Fishing for Mammalian Paradigms in the Teleost Immune System. Nat. Immunol. 2013, 14, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Mellman, I.; Steinman, R.M. Dendritic Cells. Cell 2001, 106, 255–258. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, C.; Chaves-Pozo, E. Antigen Presentation and Autophagy in Teleost Adaptive Immunity. IJMS 2022, 23, 4899. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Fumia, A.; Miller, A.; Cernigliaro, C.; Zaccone, M.; Salamone, V.; Mastrantonio, E.; Gangemi, S.; Pioggia, G. Immune System and Psychological State of Pregnant Women during COVID-19 Pandemic: Are Micronutrients Able to Support Pregnancy? Nutrients 2022, 14, 2534. [Google Scholar] [CrossRef]
- Okamura, K.; Dijkstra, J.M.; Tsukamoto, K.; Grimholt, U.; Wiegertjes, G.F.; Kondow, A.; Yamaguchi, H.; Hashimoto, K. Discovery of an Ancient MHC Category with Both Class I and Class II Features. Proc. Natl. Acad. Sci. USA 2021, 118, e2108104118. [Google Scholar] [CrossRef]
- Lovy, J.; Savidant, G.P.; Speare, D.J.; Wright, G.M. Langerin/CD207 Positive Dendritic-like Cells in the Haemopoietic Tissues of Salmonids. Fish Shellfish Immunol. 2009, 27, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Johansson, P.; Corripio-Miyar, Y.; Wang, T.; Collet, B.; Secombes, C.J.; Zou, J. Characterisation and Expression Analysis of the Rainbow Trout (Oncorhynchus mykiss) Homologue of the Human Dendritic Cell Marker CD208/Lysosomal Associated Membrane Protein 3. Dev. Comp. Immunol. 2012, 37, 402–413. [Google Scholar] [CrossRef]
- Lugo-Villarino, G.; Balla, K.M.; Stachura, D.L.; Bañuelos, K.; Werneck, M.B.F.; Traver, D. Identification of Dendritic Antigen-Presenting Cells in the Zebrafish. Proc. Natl. Acad. Sci. USA 2010, 107, 15850–15855. [Google Scholar] [CrossRef] [Green Version]
- Lauriano, E.R.; Pergolizzi, S.; Aragona, M.; Montalbano, G.; Guerrera, M.C.; Crupi, R.; Faggio, C.; Capillo, G. Intestinal Immunity of Dogfish Scyliorhinus Canicula Spiral Valve: A Histochemical, Immunohistochemical and Confocal Study. Fish Shellfish Immunol. 2019, 87, 490–498. [Google Scholar] [CrossRef]
- Wang, H.; Chen, X.; Li, S.; Zhou, C.; Xu, L.; Wu, Z.; Chen, X. Identification and Expression Analysis of Langerhans Cells Marker Langerin/CD207 in Grass Carp, Ctenopharyngodon Idella. Gene 2021, 768, 145315. [Google Scholar] [CrossRef]
- Soleto, I.; Granja, A.G.; Simón, R.; Morel, E.; Díaz-Rosales, P.; Tafalla, C. Identification of CD8α+ Dendritic Cells in Rainbow Trout (Oncorhynchus mykiss) Intestine. Fish Shellfish Immunol. 2019, 89, 309–318. [Google Scholar] [CrossRef]
- Persson, E.K.; Scott, C.L.; Mowat, A.M.; Agace, W.W. Dendritic Cell Subsets in the Intestinal Lamina Propria: Ontogeny and Function: HIGHLIGHTS. Eur. J. Immunol. 2013, 43, 3098–3107. [Google Scholar] [CrossRef] [Green Version]
- Alesci, A.; Fumia, A.; Miller, A.; Calabrò, C.; Santini, A.; Cicero, N.; Lo Cascio, P. Spirulina Promotes Macrophages Aggregation in Zebrafish (Danio rerio) Liver. Nat. Prod. Res. 2022, 1–7. [Google Scholar] [CrossRef]
- Estensoro, I.; Mulero, I.; Redondo, M.J.; Alvarez-Pellitero, P.; Mulero, V.; Sitja-Bobadilla, A. Modulation of Leukocytic Populations of Gilthead Sea Bream (Sparus aurata) by the Intestinal Parasite Enteromyxum leei (Myxozoa: Myxosporea). Parasitology 2014, 141, 425–440. [Google Scholar] [CrossRef] [Green Version]
- Volarevic, V.; Markovic, B.S.; Jankovic, M.G.; Djokovic, B.; Jovicic, N.; Harrell, C.R.; Fellabaum, C.; Djonov, V.; Arsenijevic, N.; Lukic, M.L. Galectin 3 Protects from Cisplatin-Induced Acute Kidney Injury by Promoting TLR-2-Dependent Activation of IDO1/Kynurenine Pathway in Renal DCs. Theranostics 2019, 9, 5976–6001. [Google Scholar] [CrossRef]
- Dahiya, P.; Hussain, M.A.; Mazumder, S. MtROS Induced via TLR-2-SOCE Signaling Plays Proapoptotic and Bactericidal Role in Mycobacterium Fortuitum-Infected Head Kidney Macrophages of Clarias Gariepinus. Front. Immunol. 2021, 12, 748758. [Google Scholar] [CrossRef]
- Bassity, E.; Clark, T.G. Functional Identification of Dendritic Cells in the Teleost Model, Rainbow Trout (Oncorhynchus mykiss). PLoS ONE 2012, 7, e33196. [Google Scholar] [CrossRef] [Green Version]
- Aghaallaei, N.; Bajoghli, B.; Schwarz, H.; Schorpp, M.; Boehm, T. Characterization of Mononuclear Phagocytic Cells in Medaka Fish Transgenic for a Cxcr3a:Gfp Reporter. Proc. Natl. Acad. Sci. USA 2010, 107, 18079–18084. [Google Scholar] [CrossRef] [Green Version]
- Haugland, G.T.; Jordal, A.-E.O.; Wergeland, H.I. Characterization of Small, Mononuclear Blood Cells from Salmon Having High Phagocytic Capacity and Ability to Differentiate into Dendritic like Cells. PLoS ONE 2012, 7, e49260. [Google Scholar] [CrossRef] [Green Version]
- Lauriano, E.R.; Faggio, C.; Capillo, G.; Spanò, N.; Kuciel, M.; Aragona, M.; Pergolizzi, S. Immunohistochemical Characterization of Epidermal Dendritic-like Cells in Giant Mudskipper, Periophthalmodon Schlosseri. Fish Shellfish Immunol. 2018, 74, 380–385. [Google Scholar] [CrossRef]
- Zhu, Y.; Giles, S.; Young, G.C.; Hu, Y.; Bazzi, M.; Ahlberg, P.E.; Zhu, M.; Lu, J. Endocast and Bony Labyrinth of a Devonian “Placoderm” Challenges Stem Gnathostome Phylogeny. Curr. Biol. 2021, 31, 1112–1118.e4. [Google Scholar] [CrossRef] [PubMed]
- Zaccone, D.; Icardo, J.M.; Kuciel, M.; Alesci, A.; Pergolizzi, S.; Satora, L.; Lauriano, E.R.; Zaccone, G. Polymorphous Granular Cells in the Lung of the Primitive Fish, the Bichir P Olypterus Senegalus. Acta Zool. 2017, 98, 13–19. [Google Scholar] [CrossRef]
- Kent, G.C. Anatomia Comparata dei Vertebrati; Piccin: Padova, Italy, 1997; ISBN 978-88-299-1305-3. [Google Scholar]
- Liem, K.F.; Bemis, W.E.; Grande, L. Anatomia Comparata dei Vertebrati: Una Visione Funzionale ed Evolutiva; Edises: Napoli, Italy, 2012; ISBN 978-88-7959-694-7. [Google Scholar]
- Giavini, E.; Menegola, E. Manuale di Anatomia Comparata; Edises: Napoli, Italy, 2010; ISBN 978-88-7959-550-6. [Google Scholar]
- Monforte, M.T.; Smeriglio, A.; Germanò, M.P.; Pergolizzi, S.; Circosta, C.; Galati, E.M. Evaluation of Antioxidant, Antiinflammatory, and Gastroprotective Properties of Rubus fruticosus L. Fruit Juice: Biological Properties of R. fruticosus L. Fruit Juice. Phytother. Res. 2018, 32, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Pergolizzi, S.; Rizzo, G.; Favaloro, A.; Alesci, A.; Pallio, S.; Melita, G.; Cutroneo, G.; Lauriano, E.R. Expression of VAChT and 5-HT in Ulcerative Colitis Dendritic Cells. Acta Histochem. 2021, 123, 151715. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Pergolizzi, S.; Gangemi, J.; Kuciel, M.; Capillo, G.; Aragona, M.; Faggio, C. Immunohistochemical Colocalization of G Protein Alpha Subunits and 5-HT in the Rectal Gland of the Cartilaginous Fish Scyliorhinus canicula. Microsc. Res. Tech. 2017, 80, 1018–1027. [Google Scholar] [CrossRef]
- Pergolizzi, S.; Alesci, A.; Centofanti, A.; Aragona, M.; Pallio, S.; Magaudda, L.; Cutroneo, G.; Lauriano, E.R. Role of Serotonin in the Maintenance of Inflammatory State in Crohn’s Disease. Biomedicines 2022, 10, 765. [Google Scholar] [CrossRef]
- Icardo, J.M.; Colvee, E.; Lauriano, E.R.; Capillo, G.; Guerrera, M.C.; Zaccone, G. The Structure of the Gas Bladder of the Spotted Gar, Lepisosteus Oculatus: The Gas Bladder of Lepisosteus oculatus. J. Morphol. 2015, 276, 90–101. [Google Scholar] [CrossRef]
- Zaccone, G.; Lauriano, E.R.; Capillo, G.; Kuciel, M. Air- Breathing in Fish: Air- Breathing Organs and Control of Respiration. Acta Histochem. 2018, 120, 630–641. [Google Scholar] [CrossRef]
- Zaccone, G.; Fudge, D.S.; Winegard, T.M.; Capillo, G.; Kuciel, M.; Funakoshi, K.; Lauriano, E.R. Confocal Imaging and Phylogenetic Considerations of the Subcutaneous Neurons in the Atlantic Hagfish Myxine glutinosa. Acta Zool. 2015, 96, 209–217. [Google Scholar] [CrossRef]
- Kuciel, M.; Rita Lauriano, E.; Silvestri, G.; Żuwała, K.; Pergolizzi, S.; Zaccone, D. The Structural Organization and Immunohistochemistry of G-Protein Alpha Subunits in the Olfactory System of the Air-Breathing Mudskipper, Periophthalmus Barbarus (Linnaeus, 1766) (Gobiidae, Oxudercinae). Acta Histochem. 2014, 116, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.R.; Żuwała, K.; Kuciel, M.; Budzik, K.A.; Capillo, G.; Alesci, A.; Pergolizzi, S.; Dugo, G.; Zaccone, G. Confocal Immunohistochemistry of the Dermal Glands and Evolutionary Considerations in the Caecilian, Typhlonectes natans (Amphibia: Gymnophiona). Acta Zool. 2016, 97, 154–164. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- D’Iglio, C.; Natale, S.; Albano, M.; Savoca, S.; Famulari, S.; Gervasi, C.; Lanteri, G.; Panarello, G.; Spanò, N.; Capillo, G. Otolith Analyses Highlight Morpho-Functional Differences of Three Species of Mullet (Mugilidae) from Transitional Water. Sustainability 2021, 14, 398. [Google Scholar] [CrossRef]
- Bartl, S. What Sharks Can Tell Us about the Evolution of MHC Genes. Immunol. Rev. 1998, 166, 317–331. [Google Scholar] [CrossRef]
- Grimholt, U. MHC and Evolution in Teleosts. Biology 2016, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Dijkstra, J.M.; Yamaguchi, T.; Grimholt, U. Conservation of Sequence Motifs Suggests That the Nonclassical MHC Class I Lineages CD1/PROCR and UT Were Established before the Emergence of Tetrapod Species. Immunogenetics 2018, 70, 459–476. [Google Scholar] [CrossRef]
- Flajnik, M.; Ohta, Y.; Namikawa-Yomada, C.; Nonaka, M. Insight into the Primordial MHC from Studies in Ectothermic Vertebrates. Immunol. Rev. 1999, 167, 59–67. [Google Scholar] [CrossRef]
- Nonaka, M.; Nonaka, M.I. The Evolution of Major Histocompatibility Complex in Teleosts. In The Evolution of the Immune System; Elsevier: Amsterdam, The Netherlands, 2016; pp. 331–349. ISBN 978-0-12-801975-7. [Google Scholar]
- Dijkstra, J.M.; Yamaguchi, T. Ancient Features of the MHC Class II Presentation Pathway, and a Model for the Possible Origin of MHC Molecules. Immunogenetics 2019, 71, 233–249. [Google Scholar] [CrossRef]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish. Front. Immunol. 2019, 10, 2292. [Google Scholar] [CrossRef] [Green Version]
- Dooley, H. Chondrichthyes: The Immune System of Cartilaginous Fishes. In Advances in Comparative Immunology; Cooper, E.L., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 659–685. ISBN 978-3-319-76767-3. [Google Scholar]
- Stosik, M.; Tokarz-Deptuła, B.; Deptuła, W. Major Histocompatibility Complex in Osteichthyes. J. Vet. Res. 2020, 64, 127–136. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Pergolizzi, S.; Capillo, G.; Kuciel, M.; Alesci, A.; Faggio, C. Immunohistochemical Characterization of Toll-like Receptor 2 in Gut Epithelial Cells and Macrophages of Goldfish Carassius Auratus Fed with a High-Cholesterol Diet. Fish Shellfish Immunol. 2016, 59, 250–255. [Google Scholar] [CrossRef]
- Marino, A.; Pergolizzi, S.; Lauriano, E.R.; Santoro, G.; Spataro, F.; Cimino, F.; Speciale, A.; Nostro, A.; Bisignano, G. TLR2 Activation in Corneal Stromal Cells by Staphylococcus aureus -Induced Keratitis. APMIS 2015, 123, 163–168. [Google Scholar] [CrossRef]
- Marino, A.; Santoro, G.; Spataro, F.; Lauriano, E.R.; Pergolizzi, S.; Cimino, F.; Speciale, A.; Nostro, A.; Bisignano, G.; Dugo, G. Resveratrol Role in Staphylococcus aureus -Induced Corneal Inflammation. Pathog. Dis. 2013, 68, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Alesci, A.; Pergolizzi, S.; Fumia, A.; Calabrò, C.; Lo Cascio, P.; Lauriano, E.R. Mast Cells in Goldfish (Carassius auratus) Gut: Immunohistochemical Characterization. Acta Zool. 2022, azo.12417. [Google Scholar] [CrossRef]
- Alesci, A.; Lauriano, E.R.; Aragona, M.; Capillo, G.; Pergolizzi, S. Marking Vertebrates Langerhans Cells, from Fish to Mammals. Acta Histochem. 2020, 122, 151622. [Google Scholar] [CrossRef]
- Alesci, A.; Fumia, A.; Lo Cascio, P.; Miller, A.; Cicero, N. Immunostimulant and Antidepressant Effect of Natural Compounds in the Management of COVID-19 Symptoms. J. Am. Coll. Nutr. 2021, 1–15. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Lo Cascio, P.; Capillo, G.; Lauriano, E.R. Localization of Vasoactive Intestinal Peptide and Toll-like Receptor 2 Immunoreactive Cells in Endostyle of Urochordate Styela plicata (Lesueur, 1823). Microsc. Res. Tech. 2022, jemt.24119. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Aragona, M.; Alesci, A.; Lo Cascio, P.; Pergolizzi, S. Toll-like Receptor 2 and α-Smooth Muscle Actin Expressed in the Tunica of a Urochordate, Styela Plicata. Tissue Cell 2021, 71, 101584. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Capillo, G.; Cascio, P.L.; Lauriano, E.R. Rodlet Cells in Kidney of Goldfish (Carassius auratus, Linnaeus 1758): A Light and Confocal Microscopy Study. Acta Histochem. 2022, 124, 151876. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Silvestri, G.; Kuciel, M.; Żuwała, K.; Zaccone, D.; Palombieri, D.; Alesci, A.; Pergolizzi, S. Immunohistochemical Localization of Toll-like Receptor 2 in Skin Langerhans’ Cells of Striped Dolphin (Stenella coeruleoalba). Tissue Cell 2014, 46, 113–121. [Google Scholar] [CrossRef]
- Anandhakumar, C.; Lavanya, V.; Pradheepa, G.; Tirumurugaan, K.G.; Dhinakar Raj, G.; Raja, A.; Pazhanivel, N.; Balachandran, C. Expression Profile of Toll-like Receptor 2 MRNA in Selected Tissues of Shark (Chiloscyllium sp.). Fish Shellfish Immunol. 2012, 33, 1174–1182. [Google Scholar] [CrossRef]
- Jault, C.; Pichon, L.; Chluba, J. Toll-like Receptor Gene Family and TIR-Domain Adapters in Danio Rerio. Mol. Immunol. 2004, 40, 759–771. [Google Scholar] [CrossRef]
- Oshiumi, H.; Matsumoto, M.; Funami, K.; Akazawa, T.; Seya, T. TICAM-1, an Adaptor Molecule That Participates in Toll-like Receptor 3–Mediated Interferon-β Induction. Nat. Immunol. 2003, 4, 161–167. [Google Scholar] [CrossRef]
- Mayer, W.J.; Irschick, U.M.; Moser, P.; Wurm, M.; Huemer, H.P.; Romani, N.; Irschick, E.U. Characterization of Antigen-Presenting Cells in Fresh and Cultured Human Corneas Using Novel Dendritic Cell Markers. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4459–4467. [Google Scholar] [CrossRef]
- Morrison, R.N.; Koppang, E.O.; Hordvik, I.; Nowak, B.F. MHC Class II+ Cells in the Gills of Atlantic Salmon (Salmo salar L.) Affected by Amoebic Gill Disease. Vet. Immunol. Immunopathol. 2006, 109, 297–303. [Google Scholar] [CrossRef]
- Olsen, M.M.; Kania, P.W.; Heinecke, R.D.; Skjoedt, K.; Rasmussen, K.J.; Buchmann, K. Cellular and Humoral Factors Involved in the Response of Rainbow Trout Gills to Ichthyophthirius Multifiliis Infections: Molecular and Immunohistochemical Studies. Fish Shellfish Immunol. 2011, 30, 859–869. [Google Scholar] [CrossRef]
- Kordon, A.O.; Scott, M.A.; Ibrahim, I.; Abdelhamed, H.; Ahmed, H.; Baumgartner, W.; Karsi, A.; Pinchuk, L.M. Identification of Langerhans-like Cells in the Immunocompetent Tissues of Channel Catfish, Ictalurus Punctatus. Fish Shellfish Immunol. 2016, 58, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Zhou, Q.; Zhao, C.; Lin, G.; Xu, J.; Wen, Z. An Ectoderm-Derived Myeloid-like Cell Population Functions as Antigen Transporters for Langerhans Cells in Zebrafish Epidermis. Dev. Cell 2019, 49, 605–617.e5. [Google Scholar] [CrossRef]
- Rast, J.P.; Buckley, K.M. Lamprey Immunity Is Far from Primitive. Proc. Natl. Acad. Sci. USA 2013, 110, 5746–5747. [Google Scholar] [CrossRef] [Green Version]
- Boehm, T.; Hirano, M.; Holland, S.J.; Das, S.; Schorpp, M.; Cooper, M.D. Evolution of Alternative Adaptive Immune Systems in Vertebrates. Annu. Rev. Immunol. 2018, 36, 19–42. [Google Scholar] [CrossRef]
- Mandujano-Tinoco, E.A.; Sultan, E.; Ottolenghi, A.; Gershoni-Yahalom, O.; Rosental, B. Evolution of Cellular Immunity Effector Cells; Perspective on Cytotoxic and Phagocytic Cellular Lineages. Cells 2021, 10, 1853. [Google Scholar] [CrossRef]
- Netea, M.G.; Schlitzer, A.; Placek, K.; Joosten, L.A.B.; Schultze, J.L. Innate and Adaptive Immune Memory: An Evolutionary Continuum in the Host’s Response to Pathogens. Cell Host Microbe 2019, 25, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Puhr, S.; Lee, J.; Zvezdova, E.; Zhou, Y.J.; Liu, K. Dendritic Cell Development—History, Advances, and Open Questions. Semin. Immunol. 2015, 27, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Flajnik, M.F. A Cold-Blooded View of Adaptive Immunity. Nat. Rev. Immunol. 2018, 18, 438–453. [Google Scholar] [CrossRef]
- Bacci, S.; Romagnoli, P. The Role of Dendritic Cells in Vertebrates: A Review. Int. Biol. Rev. 2017, 1, 1–9. [Google Scholar] [CrossRef]
- Yuan, S.; Tao, X.; Huang, S.; Chen, S.; Xu, A. Comparative Immune Systems in Animals. Annu. Rev. Anim. Biosci. 2014, 2, 235–258. [Google Scholar] [CrossRef]
- Bao, H.; Liu, Y.; Qin, J.; Xu, C.; Hei, N.; Jaber, J.R.; Chen, Q. An Immunohistochemical Study of S-100 Protein in the Intestinal Tract of Chinese Soft-Shelled Turtle, Pelodiscus Sinensis. Res. Vet. Sci. 2011, 91, e16–e24. [Google Scholar] [CrossRef]
- Granja, A.G.; Leal, E.; Pignatelli, J.; Castro, R.; Abós, B.; Kato, G.; Fischer, U.; Tafalla, C. Identification of Teleost Skin CD8α+ Dendritic-like Cells, Representing a Potential Common Ancestor for Mammalian Cross-Presenting Dendritic Cells. J. Immunol. 2015, 195, 1825–1837. [Google Scholar] [CrossRef] [Green Version]
- Alesci, A.; Pergolizzi, S.; Lo Cascio, P.; Fumia, A.; Lauriano, E.R. Neuronal Regeneration: Vertebrates Comparative Overview and New Perspectives for Neurodegenerative Diseases. Acta Zool. 2021, 103, 129–140. [Google Scholar] [CrossRef]
- Phrompanya, P.; Saenphet, K.; Saenphet, S. Comparative Histochemical Study of the Gastrointestinal Tracts of the Nile Tilapia (Oreochromis niloticus) and the Hybrid Catfish (Clarias batrachus x Clarias gariepinus). Acta Histochem. 2019, 121, 261–267. [Google Scholar] [CrossRef]
- Weinrauch, A.; Edwards, S.; Goss, G. Anatomy of the Pacific Hagfish (Epatatretus stoutii). In Hagfish Biology; CRC Press: Boca Raton, FL, USA, 2015; pp. 1–39. ISBN 978-1-4822-3345-2. [Google Scholar]
- Magid, A.A. The Epithelium of the Gastro-Intestinal Tract of Polypterus Senegalus (Pisces: Brachiopterygii). J. Morphol. 1975, 146, 447–456. [Google Scholar] [CrossRef]
- Verdile, N.; Pasquariello, R.; Scolari, M.; Scirè, G.; Brevini, T.A.L.; Gandolfi, F. A Detailed Study of Rainbow Trout (Onchorhynchus mykiss) Intestine Revealed That Digestive and Absorptive Functions Are Not Linearly Distributed along Its Length. Animals 2020, 10, 745. [Google Scholar] [CrossRef]
- Yadav, A.K.; Srivastava, P.P.; Chowdhary, S.; Lakra, W.S.; Shrivastava, P.; Dayal, R. Histological Alterations in the Intestine of Threatened Asian Catfish, Clarias Batrachus Fed with Different Types of Fats through Semi-Purified Diets. Adv. Appl. Sci. Res. 2014, 5, 35–41. [Google Scholar]
- Alessio, A.; Pergolizzi, S.; Gervasi, T.; Aragona, M.; Lo Cascio, P.; Cicero, N.; Lauriano, E.R. Biological Effect of Astaxanthin on Alcohol-Induced Gut Damage in Carassius auratus Used as Experimental Model. Nat. Prod. Res. 2021, 35, 5737–5743. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, H.; Zhao, C.; Zhang, H. Evolutionary History of the Toll-Like Receptor Gene Family across Vertebrates. Genome Biol. Evol. 2020, 12, 3615–3634. [Google Scholar] [CrossRef]
- Rumfelt, L.L.; Mckinney, E.C.; Taylor, E.; Flajnik, M.F. The Development of Primary and Secondary Lymphoid Tissues in the Nurse Shark Ginglymostoma cirratum: B-Cell Zones Precede Dendritic Cell Immigration and T-Cell Zone Formation during Ontogeny of the Spleen: Shark Lymphoid Organ Development. Scand. J. Immunol. 2002, 56, 130–148. [Google Scholar] [CrossRef]
- Dong, F.; Song, X.; Xing, J.; Tang, X.; Sheng, X.; Chi, H.; Zhan, W. Immunological Characteristics of Dendritic Cells Marker CD83 in Flounder (Paralichthys olivaceus). Fish Shellfish Immunol. Rep. 2021, 2, 100030. [Google Scholar] [CrossRef]
- Sharma, B.B. Dash Toll-like Receptors (TLR) in Fish. Available online: https://www.researchgate.net/publication/324257921_TOLL-LIKE_RECEPTORS_TLR_IN_FISH (accessed on 16 April 2022).
- Lewis, K.L.; Del Cid, N.; Traver, D. Perspectives on Antigen Presenting Cells in Zebrafish. Dev. Comp. Immunol. 2014, 46, 63–73. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Supplier | Dilution | Animal Source |
|---|---|---|---|
| Langerin/CD207 | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | 1:250 | Mouse |
| MHC II | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | 1:250 | Mouse |
| TLR-2 | Active Motif, La Hulpe, Belgium; Europe | 1:125 | Rabbit |
| Alexa Fluor 488 donkey anti-mouse IgG FITC conjugated | Molecular Probes, Invitrogen | 1:300 | Mouse |
| Alexa Fluor 594 donkey anti-rabbit IgG TRITC conjugated | Molecular Probes, Invitrogen | 1:300 | Rabbit |
| Lan/CD207 | TLR-2 | Lan/CD207 + TLR-2 | MHC II | TLR-2 | MHC II + TLR-2 | |
|---|---|---|---|---|---|---|
| Eptatretus cirrhatus | 116.30 ± 18.17 * | 152.70 ± 33.59 * | 107.71 ± 13.80 * | 90.34 ± 9.65 ** | 146.20 ± 11.63 ** | 76.41 ± 9.39 * |
| Scyliorhinus canicula | 145.29 ± 11.37 * | 175.17 ± 19.18 ** | 138.56 ± 27.56 ** | 168.12 ± 7.78 ** | 176.40 ± 11.57 * | 152.17 ± 11.48 ** |
| Polypterus senegalus | 203.80 ± 14.08 * | 231.10 ± 24.06 * | 193.43 ± 10.68 * | 220.75 ± 7.46 * | 251.01 ± 8.97 * | 205.72 ± 12.46 * |
| Lepisosteus oculatus | 172.72 ± 20.72 ** | 222.43 ± 17.99 * | 166.11 ± 14.44 ** | 217.58 ± 11.56 * | 238.29 ± 10.86 * | 212.10 ± 10.18 ** |
| Clarias batrachus | 226.08 ± 16.85 * | 249.98 ± 18.93 * | 215.38 ± 11.72 * | 237.59 ± 22.36 * | 261.43 ± 10.40 * | 221.67 ± 9.42 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alesci, A.; Capillo, G.; Fumia, A.; Messina, E.; Albano, M.; Aragona, M.; Lo Cascio, P.; Spanò, N.; Pergolizzi, S.; Lauriano, E.R. Confocal Characterization of Intestinal Dendritic Cells from Myxines to Teleosts. Biology 2022, 11, 1045. https://doi.org/10.3390/biology11071045
Alesci A, Capillo G, Fumia A, Messina E, Albano M, Aragona M, Lo Cascio P, Spanò N, Pergolizzi S, Lauriano ER. Confocal Characterization of Intestinal Dendritic Cells from Myxines to Teleosts. Biology. 2022; 11(7):1045. https://doi.org/10.3390/biology11071045
Chicago/Turabian StyleAlesci, Alessio, Gioele Capillo, Angelo Fumia, Emmanuele Messina, Marco Albano, Marialuisa Aragona, Patrizia Lo Cascio, Nunziacarla Spanò, Simona Pergolizzi, and Eugenia Rita Lauriano. 2022. "Confocal Characterization of Intestinal Dendritic Cells from Myxines to Teleosts" Biology 11, no. 7: 1045. https://doi.org/10.3390/biology11071045