
B-Type Natriuretic Peptide (BNP) Revisited—Is BNP Still a Biomarker for Heart Failure in the Angiotensin Receptor/Neprilysin Inhibitor Era?



Abstract
:Simple Summary
Abstract
1. Introduction
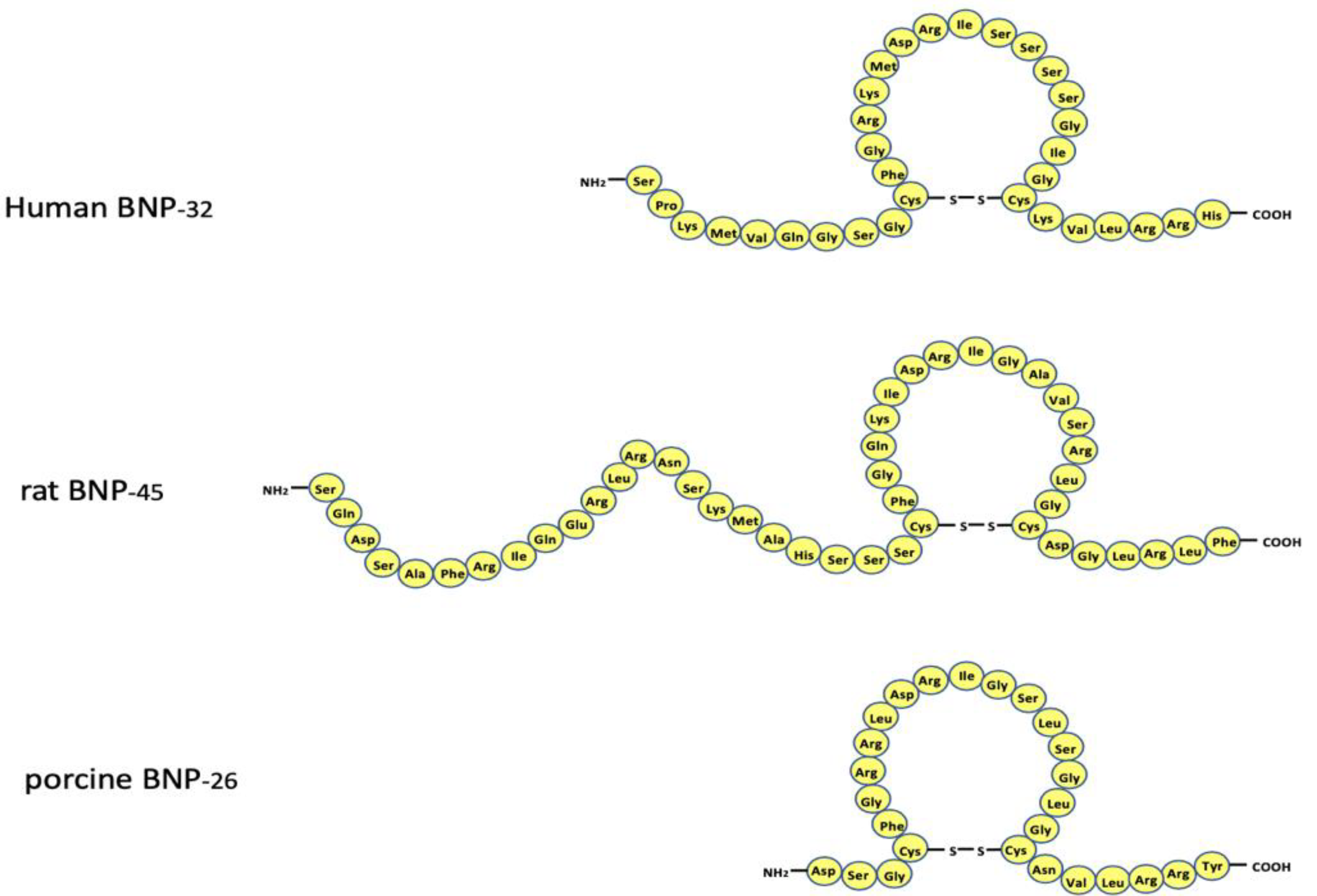
2. Structure of the BNP Gene and the Amino Acid Sequence of BNP
3. Localization and mRNA Expression of BNP
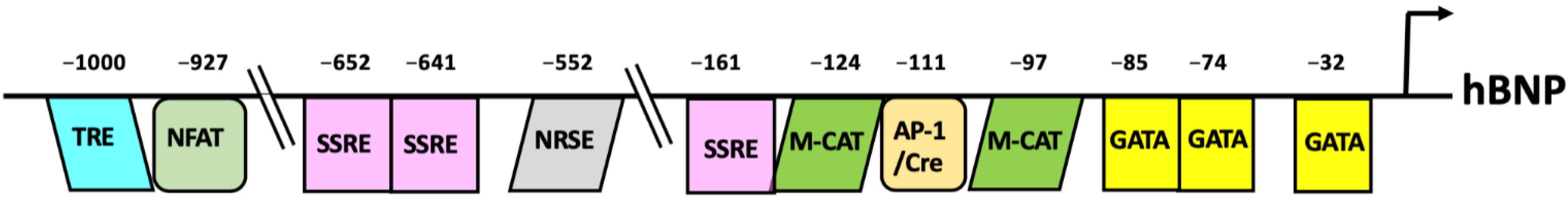
4. Transcriptional Regulation of BNP Gene
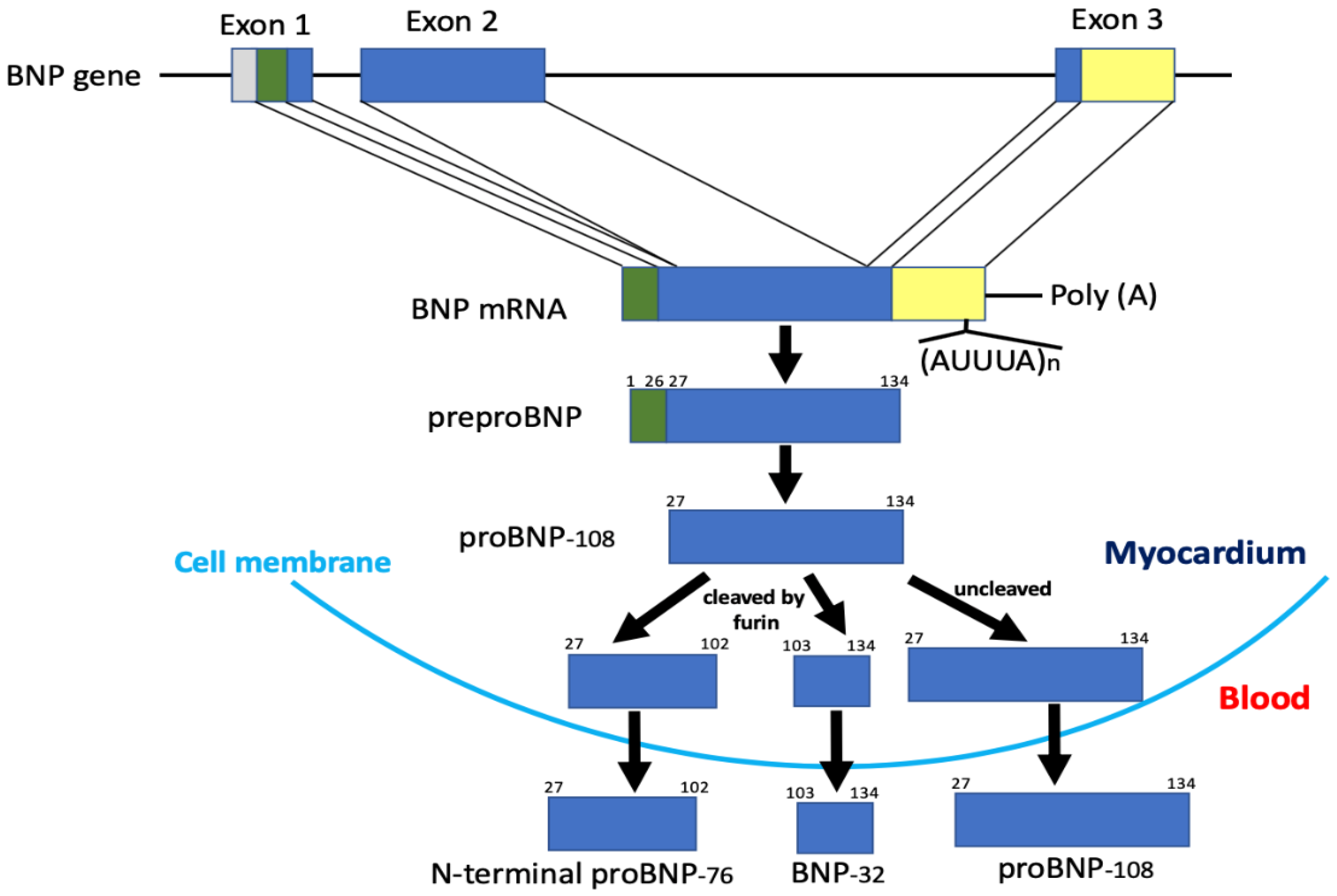
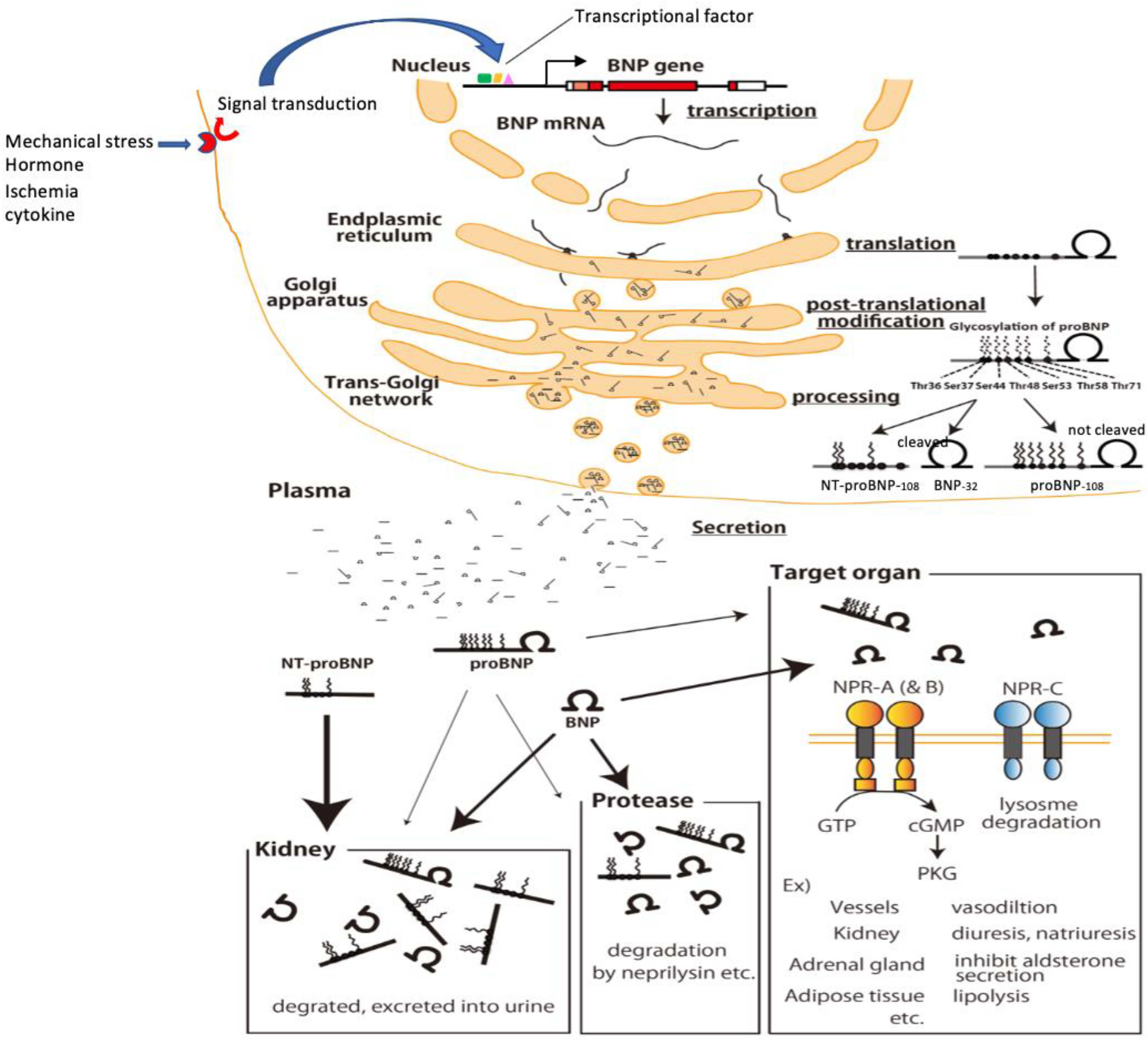
5. Production, Processing, Secretion, and Metabolism of BNP
6. Substrate Specificity of Neprilysin for ANP and BNP
7. Changes in BNP after ARNI Administration in a Clinical Study
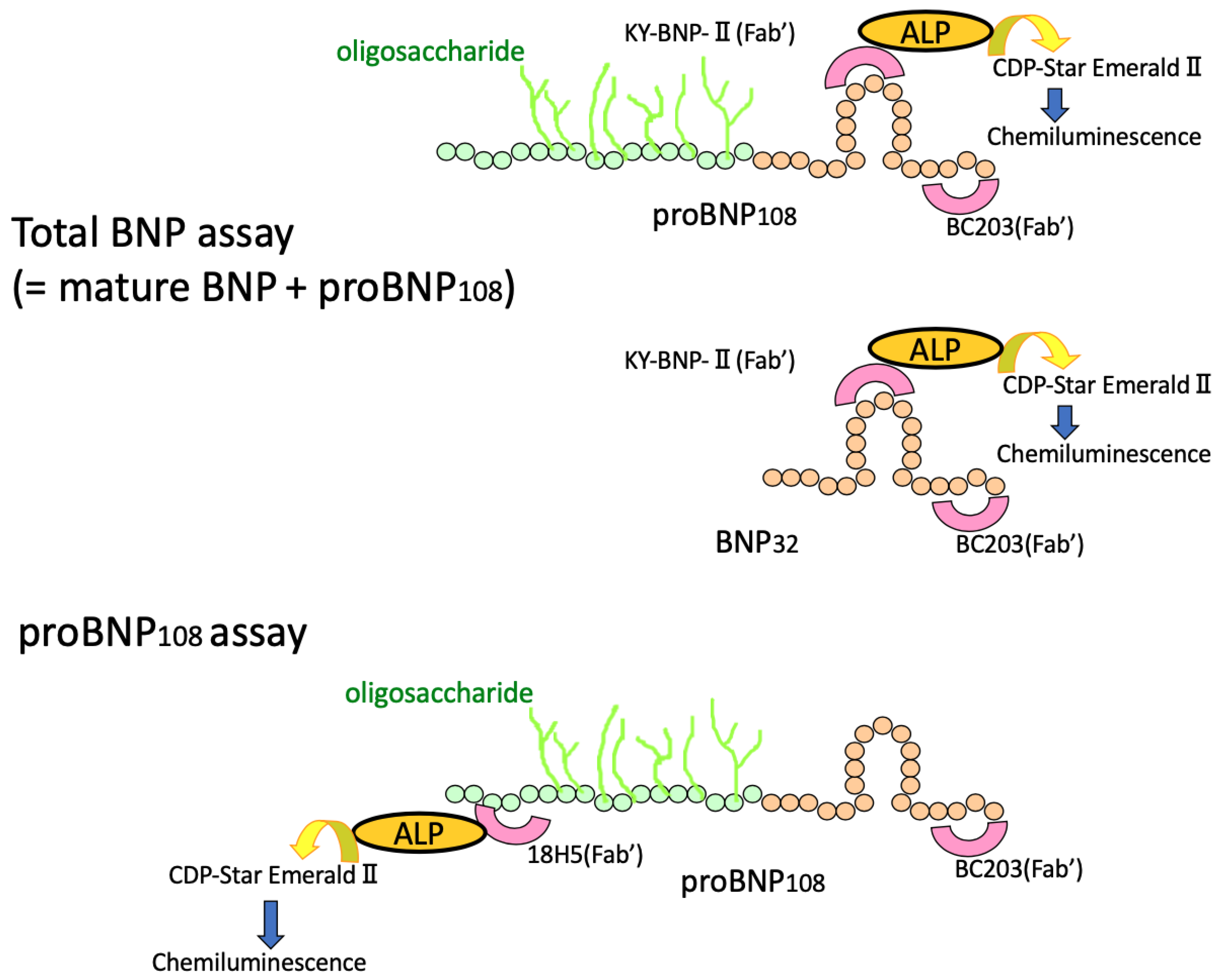
8. Immunoreactive BNP in the Plasma Means “proBNP + Mature BNP”
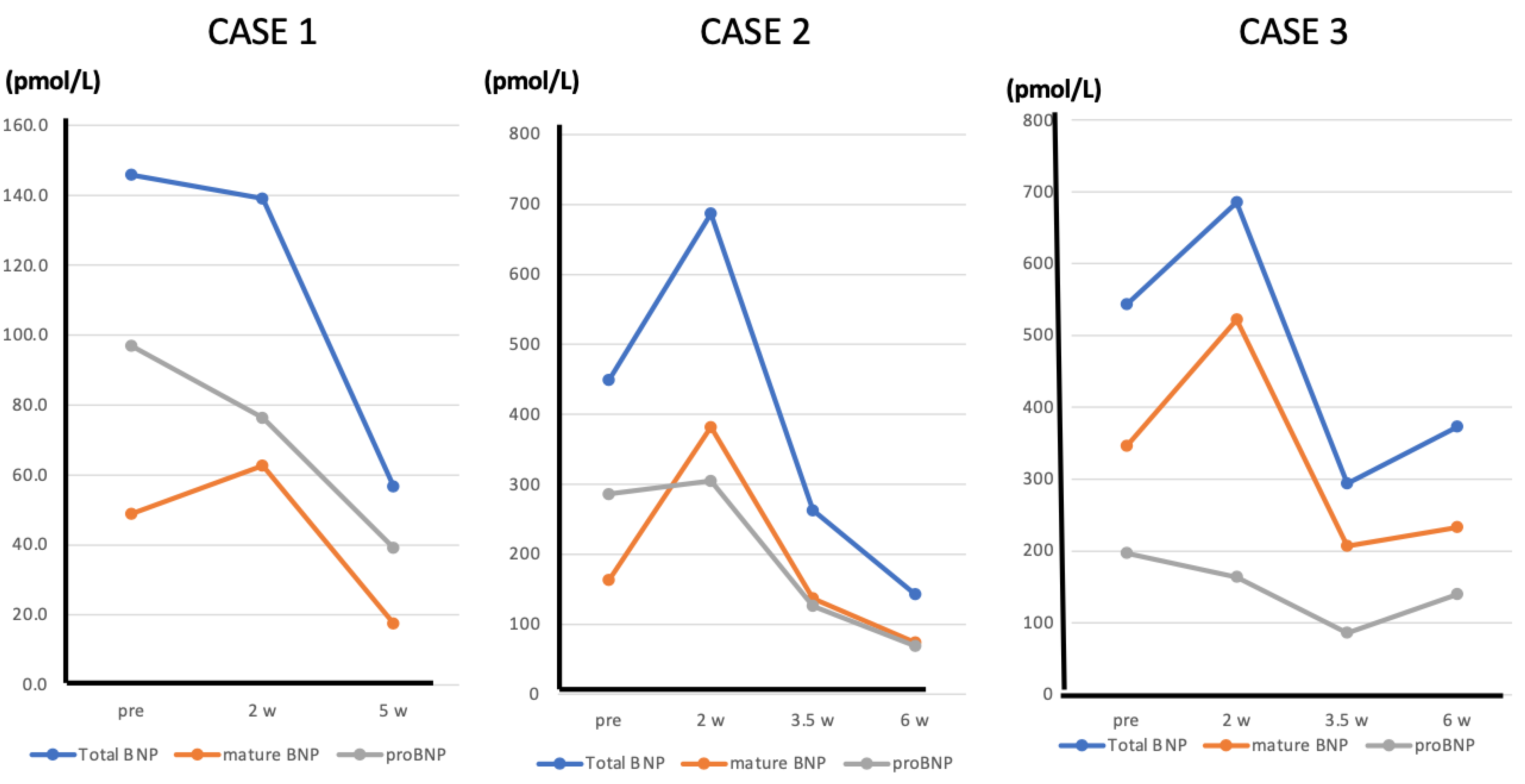
9. The Change of “Mature BNP” during ARNI Treatment—Case Presentation
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saito, Y.; Nakao, K.; Itoh, H.; Yamada, T.; Mukoyama, M.; Arai, H.; Hosoda, K.; Shirakami, G.; Suga, S.; Minamino, N.; et al. Brain natriuretic peptide is a novel cardiac hormone. Biochem. Biophys. Res. Commun. 1989, 158, 360–368. [Google Scholar] [CrossRef]
- Sudoh, T.; Kangawa, K.; Minamino, N.; Matsuo, H. A new natriuretic peptide in porcine brain. Nature 1988, 332, 78–81. [Google Scholar] [CrossRef]
- Nishikimi, T.; Yoshihara, F.; Morimoto, A.; Ishikawa, K.; Ishimitsu, T.; Saito, Y.; Kangawa, K.; Matsuo, H.; Omae, T.; Matsuoka, H. Relationship between left ventricular geometry and natriuretic peptide levels in essential hypertension. Hypertension 1996, 28, 22–30. [Google Scholar] [CrossRef]
- Morita, E.; Yasue, H.; Yoshimura, M.; Ogawa, H.; Jougasaki, M.; Matsumura, T.; Mukoyama, M.; Nakao, K. Increased plasma levels of brain natriuretic peptide in patients with acutemyocardial infarction. Circulation 1993, 88, 82–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaya, N.; Nishikimi, T.; Goto, Y.; Miyao, Y.; Kobayashi, Y.; Morii, I.; Daikoku, S.; Matsumoto, T.; Miyazaki, S.; Matsuoka, H.; et al. Plasma brain natriuretic peptide is a biochemical marker for the prediction of progressive ventricular remodeling after acute myocardial infarction. Am. Heart J. 1998, 135, 21–28. [Google Scholar] [CrossRef]
- Nishikimi, T.; Mori, Y.; Ishimura, K.; Tadokoro, K.; Yagi, H.; Yabe, A.; Horinaka, S.; Matsuoka, H. Association of plasma atrial natriuretic peptide, N-terminal proatrial natriuretic peptide, and brain natriuretic peptide levels with coronary arterystenosis in patients with normal left ventricular systolic function. Am. J. Med. 2004, 116, 517–523. [Google Scholar] [CrossRef]
- Nagaya, N.; Nishikimi, T.; Okano, Y.; Uematsu, M.; Satoh, T.; Kyotani, S.; Kuribayashi, S.; Hamada, S.; Kakishita, M.; Nakanishi, N.; et al. Plasma brain natriuretic peptide levels increase in proportion to the extent of right ventricular dysfunction in pulmonary hypertension. J. Am. Coll. Cardiol. 1998, 31, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Mukoyama, M.; Nakao, K.; Hosoda, K.; Suga, S.; Saito, Y.; Ogawa, Y.; Shirakami, G.; Jougasaki, M.; Obata, K.; Yasue, H.; et al. Brain natriuretic peptide as a novel cardiac hormone in humans. J. Clin. Investig. 1991, 87, 1402–1412. [Google Scholar] [CrossRef]
- Yasue, H.; Yoshimura, M.; Sumida, H.; Kikuta, K.; Kugiyama, K.; Jougasaki, M.; Ogawa, H.; Okumura, K.; Mukoyama, M.; Nakao, K. Localization and mechanism of secretion of B-type natriuretic peptide in comparison with those of A-type natriuretic peptide in normal subjects and patients with heart failure. Circulation 1994, 90, 195–203. [Google Scholar] [CrossRef] [Green Version]
- McDonagh, T.A.; Metra, M.; Adamo, M.; Gardner, R.S.; Baumbach, A.; Böhm, M.; Burri, H.; Butler, J.; Čelutkienė, J.; Chioncel, O.; et al. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. Heart J. 2021, 42, 3599–3726. [Google Scholar]
- Heidenreich, P.A.; Bozkurt, B.; Aguilar, D.; Allen, L.A.; Byun, J.J.; Colvin, M.M.; Deswal, A.; Drazner, M.H.; Dunlay, S.M.; Evers, L.R.; et al. 2022 AHA/ACC/HFSA Guideline for the Management of Heart Failure: A Report of the American College of Cardiology/American Heart Association Joint Committee on Clinical Practice Guidelines. Circulation 2022, 145, e895–e1032. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.P.; Borque, L.; Rus, A.; Vicente, B.; Ramírez, Y.; Lasa, L. Comparison of BNP and NT-proBNP assays in the approach to the emergency diagnosis of acute dyspnea. J. Clin. Lab. Anal. 2006, 20, 227–232. [Google Scholar] [CrossRef]
- Clerico, A.; Fontana, M.; Zyw, L.; Passino, C.; Emdin, M. Comparison of the diagnostic accuracy of brain natriuretic peptide (BNP) and the N-terminal part of the propeptide of BNP immunoassays in chronic and acute heart failure: A systematic review. Clin. Chem. 2007, 53, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikimi, T.; Nakagawa, Y. Potential pitfalls when interpreting plasma BNP levels in heart failure practice. J. Cardiol. 2021, 78, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Januzzi, J.L.; van Kimmenade, R.; Lainchbury, J.; Bayes-Genis, A.; Ordonez-Llanos, J.; Santalo-Bel, M.; Pinto, Y.M.; Richards, M. NT-proBNP testing for diagnosis and short-term prognosis in acute destabilized heart failure: An international pooled analysis of 1256 patients: The International Collaborative of NT-proBNP Study. Eur. Heart J. 2006, 27, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Dao, Q.; Krishnaswamy, P.; Kazanegra, R.; Harrison, A.; Amirnovin, R.; Lenert, L.; Clopton, P.; Alberto, J.; Hlavin, P.; Maisel, A.S. Utility of B-type natriuretic peptide in the diagnosis of congestive heart failurein an urgent-care setting. J. Am. Coll. Cardiol. 2001, 37, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Maisel, A.S.; Krishnaswamy, P.; Nowak, R.M.; McCord, J.; Hollander, J.E.; Duc, P.; Omland, T.; Storrow, A.B.; Abraham, W.T.; Wu, A.H.; et al. Rapid measurement of B-type natriuretic peptide in the emergency diagnosis of heart failure. N. Engl. J. Med. 2002, 347, 161–167. [Google Scholar] [CrossRef]
- McCullough, P.A.; Nowak, R.M.; McCord, J.; Hollander, J.E.; Herrmann, H.C.; Steg, P.G.; Duc, P.; Westheim, A.; Omland, T.; Knudsen, C.W.; et al. B-type natriuretic peptide and clinical judgment in emergency diagnosis of heart failure: Analysis from Breathing Not Properly (BNP) Multinational Study. Circulation 2002, 106, 416–422. [Google Scholar] [CrossRef] [Green Version]
- McMurray, J.J.; Packer, M.; Desai, A.S.; Gong, J.; Lefkowitz, M.P.; Rizkala, A.R.; Rouleau, J.L.; Shi, V.C.; Solomon, S.D.; Swedberg, K.; et al. Angiotensin-neprilysin inhibition versus enalapril in heart failure. N. Engl. J. Med. 2014, 371, 993–1004. [Google Scholar] [CrossRef] [Green Version]
- Packer, M.; McMurray, J.J.; Desai, A.S.; Gong, J.; Lefkowitz, M.P.; Rizkala, A.R.; Rouleau, J.L.; Shi, V.C.; Solomon, S.D.; Swedberg, K.; et al. Angiotensin receptor neprilysin inhibition compared with enalapril on the risk of clinical progression in surviving patients with heart failure. Circulation 2015, 131, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Tateyama, H.; Hino, J.; Minamino, N.; Kangawa, K.; Ogihara, T.; Matsuo, H. Characterization of immunoreactive brain natriuretic peptide in human cardiac atrium. Biochem. Biophys. Res. Commun. 1990, 166, 1080–1087. [Google Scholar] [CrossRef]
- Hino, J.; Tateyama, H.; Minamino, N.; Kangawa, K.; Matsuo, H. Isolation and identification of human brain natriuretic peptides in cardiac atrium. Biochem. Biophys. Res. Commun. 1990, 167, 693–700. [Google Scholar] [CrossRef]
- Minamino, N.; Horio, H.; Nishikimi, T. Natriuretic peptides in the cardiovascular system. In The Handbook of Biologically Active Peptides, 1st ed.; Kastin, A.J., Ed.; Academic Press: New York, NY, USA; London, UK, 2006; pp. 1217–1225. [Google Scholar]
- Nishikimi, T.; Nakagawa, Y.; Minamino, N.; Ikeda, M.; Tabei, K.; Fujishima, A.; Takayama, K.; Akimoto, K.; Yamada, C.; Nakao, K.; et al. Pro-B-type natriuretic peptide is cleaved intracellularly: Impact of distance between O-glycosylation and cleavage sites. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R639–R649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, Y.; Itoh, H.; Nakagawa, O.; Shirakami, G.; Tamura, N.; Yoshimasa, T.; Nagata, K.; Yoshida, N.; Nakao, K. Characterization of the 5′-flanking region and chromosomal assignment of the human brain natriuretic peptide gene. J. Mol. Med. 1995, 73, 457–463. [Google Scholar] [CrossRef]
- Nishikimi, T.; Kuwahara, K.; Nakao, K. Current biochemistry, molecular biology, and clinical relevance of natriuretic peptides. J. Cardiol. 2011, 57, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Goetze, J.P.; Bruneau, B.G.; Ramos, H.R.; Ogawa, T.; de Bold, M.K.; de Bold, A.J. Cardiac natriuretic peptides. Nat. Rev. Cardiol. 2020, 17, 698–717. [Google Scholar] [CrossRef]
- Schellenberger, U.; O’Rear, J.; Guzzetta, A.; Jue, R.A.; Protter, A.A.; Pollitt, N.S. The precursor to B-type natriuretic peptide is an O-linked glycoprotein. Arch. Biochem. Biophys. 2006, 451, 160–166. [Google Scholar] [CrossRef]
- Nishikimi, T.; Minamino, N.; Masashi, I.; Takeda, Y.; Tadokoro, K.; Shibasaki, I.; Fukuda, H.; Horiuchi, Y.; Oikawa, S.; Ieiri, T.; et al. Diversity of molecular forms of plasma brain natriuretic peptide in heart failure—Different proBNP- 108 to BNP-32 ratios in atrial and ventricular overload. Heart 2010, 96, 432–439. [Google Scholar] [CrossRef]
- Nakayama, K. Furin: A mammalian subtilisin/Kex2p-like endo- protease involved in processing of a wide variety of precursor proteins. Biochem. J. 1997, 327, 625–635. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, I.; Rieunier, F.; Larue, C.; Delagneau, J.F.; Granier, C.; Pau, B.; Ferrière, M.; Saussine, M.; Cristol, J.P.; Dupuy, A.M.; et al. Assay for measurement of intact B-type natriuretic peptide prohormone in blood. Clin. Chem. 2006, 52, 1054–1061. [Google Scholar] [CrossRef]
- Lam, C.S.; Burnett, J.C., Jr.; Costello-Boerrigter, L.; Rodeheffer, R.J.; Redfield, M.M. Alternate circulating pro-B-type natriuretic peptide and B-type natriuretic peptide forms in the general population. J. Am. Coll. Cardiol. 2007, 49, 1193–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogelvang, R.; Goetze, J.P.; Schnohr, P.; Lange, P.; Sogaard, P.; Rehfeld, J.F.; Jensen, J.S. Discriminating between cardiac and pulmonary dysfunction in the general population with dyspnea by plasma pro-B-type natriuretic peptide. J. Am. Coll. Cardiol. 2007, 50, 1694–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, F.; O’Rear, J.; Schellenberger, U.; Tai, L.; Lasecki, M.; Schreiner, G.F.; Apple, F.S.; Maisel, A.S.; Pollitt, N.S.; Protter, A.A. Evidence for functional heterogeneity of circulating B-type natriuretic peptide. J. Am. Coll. Cardiol. 2007, 49, 1071–1078. [Google Scholar] [CrossRef] [Green Version]
- Seferian, K.R.; Tamm, N.N.; Semenov, A.G.; Mukharyamova, K.S.; Tolstaya, A.A.; Koshkina, E.V.; Kara, A.N.; Krasnoselsky, M.I.; Apple, F.S.; Esakova, T.V.; et al. The brain natriuretic peptide (BNP) precursor is the major immunoreactive form of BNP in patients with heart failure. Clin. Chem. 2007, 53, 866–873. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, Y.; Nishikimi, T.; Kuwahara, K.; Fujishima, A.; Oka, S.; Tsutamoto, T.; Kinoshita, H.; Nakao, K.; Cho, K.; Inazumi, H.; et al. MiR30-GALNT1/2 Axis-Mediated Glycosylation Contributes to the Increased Secretion of Inactive Human Prohormone for Brain Natriuretic Peptide (proBNP) From Failing Hearts. J. Am. Heart Assoc. 2017, 6, e003601. [Google Scholar] [CrossRef] [Green Version]
- Semenov, A.G.; Postnikov, A.B.; Tamm, N.N.; Seferian, K.R.; Karpova, N.S.; Bloshchitsyna, M.N.; Koshkina, E.V.; Krasnoselsky, M.I.; Serebryanaya, D.V.; Katrukha, A.G. Processing of pro-brain natriuretic peptide is suppressed by O-glycosylation in the region close to the cleavage site. Clin. Chem. 2009, 55, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, Y.; Nakao, K.; Mukoyama, M.; Shirakami, G.; Itoh, H.; Hosoda, K.; Saito, Y.; Arai, H.; Suga, S.; Jougasaki, M.; et al. Rat brain natriuretic peptide--tissue distribution and molecular form. Endocrinology 1990, 126, 2225–2227. [Google Scholar] [CrossRef] [PubMed]
- Knappe, S.; Wu, F.; Madlansacay, M.R.; Wu, Q. Identification of domain structures in the propeptide of corin essential for the processing of proatrial natriuretic peptide. J. Biol. Chem. 2004, 279, 34464–34471. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.C.; Knudson, O.; Wu, F.; Morser, J.; Dole, W.P.; Wu, Q. Hypertension in mice lacking the proatrial natriuretic peptide convertase corin. Proc. Natl. Acad. Sci. USA 2005, 102, 785–790. [Google Scholar] [CrossRef] [Green Version]
- Nishikimi, T.; Kuwahara, K.; Nakagawa, Y.; Kangawa, K.; Minamino, N.; Nakao, K. Complexity of molecular forms of B-type natriuretic peptide in heart failure. Heart 2013, 99, 677–679. [Google Scholar] [CrossRef]
- Waldo, S.W.; Beede, J.; Isakson, S.; Villard-Saussine, S.; Fareh, J.; Clopton, P.; Fitzgerald, R.L.; Maisel, A.S. Pro-B-type natriuretic peptide levels in acute decompensated heart failure. J. Am. Coll. Cardiol. 2008, 51, 1874–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, O.; Ogawa, Y.; Itoh, H.; Suga, S.; Komatsu, Y.; Kishimoto, I.; Nishino, K.; Yoshimasa, T.; Nakao, K. Rapid transcriptional activation and early mRNA turnover of brain natriuretic peptide in cardiocyte hypertrophy. Evidence for brain natriuretic peptide as an “emergency” cardiac hormone against ventricular overload. J. Clin. Investig. 1995, 96, 1280–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.K.; Banas, K.; de Bold, A.J. Determinants of inducible brain natriuretic peptide promoter activity. Regul. Pept. 2005, 128, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, K.; Nishikimi, T.; Nakao, K. Transcriptional regulation of the fetal cardiac gene program. J. Pharmacol. Sci. 2012, 119, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Hayek, S.; Nemer, M. Cardiac natriuretic peptides: From basic discovery to clinical practice. Cardiovasc. Ther. 2011, 29, 362–376. [Google Scholar] [CrossRef]
- He, Q.; Wang, D.; Yang, X.P.; Carretero, O.A.; LaPointe, M.C. Inducible regulation of human brain natriuretic peptide promoter in transgenic mice. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H368–H376. [Google Scholar] [CrossRef]
- LaPointe, M.C.; Wu, G.; Garami, M.; Yang, X.P.; Gardner, D.G. Tissue- specific expression of the human brain natriuretic peptide gene in cardiac myocytes. Hypertension 1996, 27, 715–722. [Google Scholar] [CrossRef]
- Kuwahara, K.; Nakao, K. Regulation and significance of atrial and brain natriuretic peptides as cardiac hormones. Endocr. J. 2010, 57, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Pikkarainen, S.; Tokola, H.; Majalahti-Palviainen, T.; Kerkela, R.; Hautala, N.; Bhalla, S.S.; Charron, F.; Nemer, M.; Vuolteenaho, O.; Ruskoaho, H. GATA-4 is a nuclear mediator of mechanical stretch-activated hypertrophic program. J. Biol. Chem. 2003, 278, 23807–23816. [Google Scholar] [CrossRef] [Green Version]
- Herzig, T.C.; Jobe, S.M.; Aoki, H.; Molkentin, J.D.; Cowley, A.W., Jr.; Izumo, S.; Markham, B.E. Angiotensin II type1a receptor gene expression in the heart: AP-1 and GATA-4 participate in the response to pressure overload. Proc. Natl. Acad. Sci. USA 1997, 94, 7543–7548. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, E.; Saito, Y.; Harada, M.; Kamitani, S.; Kuwahara, K.; Miyamoto, Y.; Ishikawa, M.; Hamanaka, I.; Kajiyama, N.; Takahashi, N.; et al. Outside-in signalling of fibronectin stimulates cardiomyocyte hypertrophy in cultured neonatal rat ventricular myocytes. J. Mol. Cell. Cardiol. 2000, 32, 765–776. [Google Scholar] [CrossRef]
- Liang, F.; Gardner, D.G. Mechanical strain activates BNP gene transcription through a p38/NF-kappaB-dependent mechanism. J. Clin. Investig. 1999, 104, 1603–1612. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, E.; Saito, Y.; Kuwahara, K.; Harada, M.; Miyamoto, Y.; Hamanaka, I.; Kajiyama, N.; Takahashi, N.; Izumi, T.; Kawakami, R.; et al. Fibronectin signaling stimulates BNP gene transcription by inhibiting neuron- restrictive silencer element-dependent repression. Cardiovasc. Res. 2002, 53, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, K.; Saito, Y.; Takano, M.; Arai, Y.; Yasuno, S.; Nakagawa, Y.; Takahashi, N.; Adachi, Y.; Takemura, G.; Horie, M.; et al. NRSF reg- ulates the fetal cardiac gene program and maintains normal cardiac structure and function. EMBO J. 2003, 22, 6310–6321. [Google Scholar] [CrossRef]
- Inazumi, H.; Kuwahara, K.; Nakagawa, Y.; Kuwabara, Y.; Numaga-Tomita, T.; Kashihara, T.; Nakada, T.; Kurebayashi, N.; Oya, M.; Nonaka, M.; et al. NRSF-GNAO1 Pathway Contributes to the Regulation of Cardiac Ca2+ Homeostasis. Circ. Res. 2022, 130, 234–248. [Google Scholar] [CrossRef]
- Nishikimi, T.; Ikeda, M.; Takeda, Y.; Ishimitsu, T.; Shibasaki, I.; Fukuda, H.; Kinoshita, H.; Nakagawa, Y.; Kuwahara, K.; Nakao, K. The effect of glycosylation on plasma N-terminal proBNP-76 levels in patients with heart or renal failure. Heart 2012, 98, 152–161. [Google Scholar] [CrossRef]
- Singh, S.; Lowe, D.G.; Thorpe, D.S.; Rodriguez, H.; Kuang, W.J.; Dangott, L.J.; Chinkers, M.; Goeddel, D.V.; Garbers, D.L. Membrane guanylate cyclase is a cell-surface receptor with homology to protein kinases. Nature 1988, 334, 708–712. [Google Scholar] [CrossRef]
- Nishikimi, T.; Maeda, N.; Matsuoka, H. The role of natriuretic peptides in cardioprotection. Cardiovasc. Res. 2006, 69, 318–328. [Google Scholar] [CrossRef]
- Fuller, F.; Porter, J.G.; Arfsten, A.E.; Miller, J.; Schilling, J.W.; Scarborough, R.M.; Lewicki, J.A.; Schenk, D.B. Atrial natriuretic peptide clearance receptor. Complete sequence and functional expression of cDNA clones. J. Biol. Chem. 1988, 263, 9395–9401. [Google Scholar] [CrossRef]
- Anand-Srivastava, M.B. Natriuretic peptide receptor-C signaling and regulation. Peptides 2005, 26, 1044–1059. [Google Scholar] [CrossRef]
- Rose, R.A.; Giles, W.R. Natriuretic peptide C receptor signalling in the heart and vasculature. J. Physiol. 2008, 586, 353–366. [Google Scholar] [CrossRef]
- Moyes, A.J.; Hobbs, A.J. C-type Natriuretic Peptide: A Multifaceted Paracrine Regulator in the Heart and Vasculature. Int. J. Mol. Sci. 2019, 20, 2281. [Google Scholar] [CrossRef] [Green Version]
- Abbas, S.S.; Mahmoud, H.M.; Schaalan, M.F.; El-Abhar, H.S. Involvement of brain natriuretic peptide signaling pathway in the cardioprotective action of sitagliptin. Pharmacol. Rep. 2018, 70, 720–729. [Google Scholar] [CrossRef]
- Müller, D.; Schulze, C.; Baumeister, H.; Buck, F.; Richter, D. Rat insulin-degrading enzyme: Cleavage pattern of the natriuretic peptide hormones ANP, BNP, and CNP revealed by HPLC and mass spectrometry. Biochemistry 1992, 31, 11138–11143. [Google Scholar] [CrossRef]
- Pankow, K.; Wang, Y.; Gembardt, F.; Krause, E.; Sun, X.; Krause, G.; Schultheiss, H.-P.; Siems, W.; Walther, T. Successive action of meprin A and neprilysin catabolizes B-type natriuretic peptide. Circ. Res. 2007, 101, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Bourne, A.; Kenny, A.J. The hydrolysis of brain and atrial natriuretic peptides by porcine choroid plexus is attributable to endopeptidase-24.11. Biochem. J. 1990, 271, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Semenov, A.G.; Katrukha, A.G. Different Susceptibility of B-Type Natriuretic Peptide (BNP) and BNP Precursor (proBNP) to Cleavage by Neprilysin: The N-Terminal Part Does Matter. Clin. Chem. 2016, 62, 617–622. [Google Scholar] [CrossRef] [Green Version]
- Palmer, S.C.; Endre, Z.H.; Richards, A.M.; Yandle, T.G. Characterization of NT-proBNP in human urine. Clin. Chem. 2009, 55, 1126–1134. [Google Scholar] [CrossRef] [Green Version]
- Nishikimi, T.; Okamoto, H.; Nakamura, M.; Ogawa, N.; Horii, K.; Nagata, K.; Nakagawa, Y.; Kinoshita, H.; Yamada, C.; Nakao, K.; et al. Direct immunochemiluminescent assay for proBNP and total BNP in human plasma proBNP and total BNP levels in normal and heart failure. PLoS ONE 2013, 8, e53233. [Google Scholar] [CrossRef]
- Spanaus, K.S.; Kronenberg, F.; Ritz, E.; Schlapbach, R.; Fliser, D.; Hersberger, M.; Kollerits, B.; König, P.; von Eckardstein, A.; Mild-to-Moderate Kidney Disease Study Group. B-type natriuretic peptide concentrations predict the progression of nondiabetic chronic kidney disease: The Mild-to-Moderate Kidney Disease Study. Clin. Chem. 2007, 53, 1264–1272. [Google Scholar] [CrossRef] [Green Version]
- McGregor, A.; Richards, M.; Espiner, E.; Yandle, T.; Ikram, H. Brain natriuretic peptide administered to man: Actions and metabolism. J. Clin. Endocrinol. Metab. 1990, 70, 1103–1107. [Google Scholar] [CrossRef]
- Yandle, T.G.; Richards, A.M.; Nicholls, M.G.; Cuneo, R.; Espiner, E.A.; Livesey, J.H. Metabolic clearance rate and plasma half life of alpha-human atrial natriuretic peptide in man. Life Sci. 1986, 38, 1827–1833. [Google Scholar] [CrossRef]
- Suga, S.; Nakao, K.; Hosoda, K.; Mukoyama, M.; Ogawa, Y.; Shirakami, G.; Arai, H.; Saito, Y.; Kambayashi, Y.; Inouye, K.; et al. Receptor selectivity of natriuretic peptide family, atrial natriuretic peptide, brain natriuretic peptide, and C-type natriuretic peptide. Endocrinology 1992, 130, 229–239. [Google Scholar] [CrossRef]
- Ozaki, J.; Shimizu, H.; Hashimoto, Y.; Itoh, H.; Nakao, K.; Inui, K. Enzymatic inactivation of major circulating forms of atrial and brain natriuretic peptides. Eur. J. Pharmacol. 1999, 370, 307–312. [Google Scholar] [CrossRef]
- Watanabe, Y.; Nakajima, K.; Shimamori, Y.; Fujimoto, Y. Comparison of the hydrolysis of the three types of natriuretic peptides by human kidney neutral endopeptidase 24.11. Biochem. Mol. Med. 1997, 61, 47–51. [Google Scholar] [CrossRef]
- Roques, B.P.; Noble, F.; Daugé, V.; Fournié-Zaluski, M.C.; Beaumont, A. Neutral endopeptidase 24.11: Structure, inhibition, and experimental and clinical pharmacology. Pharmacol. Rev. 1993, 45, 87–146. [Google Scholar]
- Kentsch, M.; Otter, W. Novel neurohormonal modulators in cardiovascular disorders. The therapeutic potential of endopeptidase inhibitors. Drugs R D 1999, 1, 331–338. [Google Scholar] [CrossRef]
- Pankow, K.; Schwiebs, A.; Becker, M.; Siems, W.E.; Krause, G.; Walther, T. Structural substrate conditions required for neutral endopeptidase-mediated natriuretic Peptide degradation. J. Mol. Biol. 2009, 393, 496–503. [Google Scholar] [CrossRef]
- Vodovar, N.; Séronde, M.F.; Laribi, S.; Gayat, E.; Lassus, J.; Januzzi, J.L., Jr.; Boukef, R.; Nouira, S.; Manivet, P.; Samuel, J.L.; et al. Elevated Plasma B-Type Natriuretic Peptide Concentrations Directly Inhibit Circulating Neprilysin Activity in Heart Failure. JACC Heart Fail. 2015, 3, 629–636. [Google Scholar] [CrossRef]
- Murphy, S.P.; Prescott, M.F.; Camacho, A.; Iyer, S.R.; Maisel, A.S.; Felker, G.M.; Butler, J.; Piña, I.L.; Ibrahim, N.E.; Abbas, C.; et al. Atrial Natriuretic Peptide and Treatment with Sacubitril/Valsartan in Heart Failure with Reduced Ejection Fraction. JACC Heart Fail. 2021, 9, 127–136. [Google Scholar] [CrossRef]
- Richards, A.M.; Crozier, I.G.; Espiner, E.A.; Yandle, T.G.; Nicholls, M.G. Plasma brain natriuretic peptide and endopeptidase 24.11 inhibition in hypertension. Hypertension 1993, 22, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Nougué, H.; Pezel, T.; Picard, F.; Sadoune, M.; Arrigo, M.; Beauvais, F.; Launay, J.M.; Cohen-Solal, A.; Vodovar, N.; Logeart, D. Effects of sacubitril/valsartan on neprilysin targets and the metabolism of natriuretic peptides in chronic heart failure: A mechanistic clinical study. Eur. J. Heart Fail. 2019, 21, 598–605. [Google Scholar] [CrossRef]
- Pereira, N.L.; Desai, V.K.; Jaffe, A.S. Beneficial effects of sacubitril/valsartan in heart failure with reduced ejection fraction: Pas à cause du BNP? Eur. J. Heart Fail. 2019, 21, 609–612. [Google Scholar] [CrossRef]
- Velazquez, E.J.; Morrow, D.A.; DeVore, A.D.; Duffy, C.I.; Ambrosy, A.P.; McCague, K.; Rocha, R.; Braunwald, E.; PIONEER-HF Investigators. Angiotensin-Neprilysin Inhibition in Acute Decompensated Heart Failure. N. Engl. J. Med. 2019, 380, 539–548. [Google Scholar] [CrossRef]
- Myhre, P.L.; Vaduganathan, M.; Claggett, B.; Packer, M.; Desai, A.S.; Rouleau, J.L.; Zile, M.R.; Swedberg, K.; Lefkowitz, M.; Shi, V.; et al. B-Type Natriuretic Peptide during Treatment with Sacubitril/Valsartan: The PARADIGM-HF Trial. J. Am. Coll. Cardiol. 2019, 73, 1264–1272. [Google Scholar] [CrossRef]
- Abellas-Sequeiros, M.; Del Prado Díaz, S.; Del Rey, J.M.; Alonso-Salinas, G.L.; Zamorano, J.L. Sacubitril/valsartan and B-type natriuretic peptide value: Recalculating route. Eur. J. Heart Fail. 2020, 22, 162–163. [Google Scholar] [CrossRef]
- Pugliese, N.R.; Fabiani, I.; Zywicki, V.; Mazzola, M.; D’Agostino, A.; Galeotti, G.G.; Dini, F.L. Effects of sacubitril/valsartan on B-type natriuretic peptide circulating levels and loop diuretic dose in a case series of stabilized heart failure patients with left ventricular ejection fraction ≤35. Curr. Med. Res. Opin. 2019, 35, 13–18. [Google Scholar] [CrossRef]
- Tiepolo, A.; Nougué, H.; Damoisel, C.; Launay, J.M.; Vodovar, N.; Mebazaa, A. Evolution of B-type natriuretic peptide and N-terminal pro-brain natriuretic peptide during acute decompensated heart failure in a chronic heart failure patient with reduced ejection fraction treated with Sacubitril/Valsartan: A case report. Eur. Heart J. Case Rep. 2019, 3, ytz108. [Google Scholar] [CrossRef]
- Dillon, E.M.; Wei, S.D.; Gupta, D.K.; Nian, H.; Rodibaugh, B.S.; Bachmann, K.N.; Naftilan, A.J.; Stevenson, L.W.; Brown, N.J. Active B-Type Natriuretic Peptide Measured by Mass Spectrometry and Response to Sacubitril/Valsartan. J. Card. Fail. 2021, 27, 1231–1239. [Google Scholar] [CrossRef]
- Apple, F.S.; Panteghini, M.; Ravkilde, J.; Mair, J.; Wu, A.H.; Tate, J.; Pagani, F.; Christenson, R.H.; Jaffe, A.S. Quality specifications for B-type natriuretic peptide assays. Clin. Chem. 2005, 51, 486–493. [Google Scholar] [CrossRef] [Green Version]
- Krishnaswami, A. The role of B-type and other natriuretic peptides in health and disease. Perm. J. 2008, 12, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Ping, P.; Zhu, Q.; Ye, P.; Luo, L. Brain Natriuretic Peptide and Its Biochemical, Analytical, and Clinical Issues in Heart Failure: A Narrative Review. Front. Physiol. 2018, 9, 692. [Google Scholar] [CrossRef]
- Prontera, C.; Zaninotto, M.; Giovannini, S.; Zucchelli, G.C.; Pilo, A.; Sciacovelli, L.; Plebani, M.; Clerico, A. Proficiency testing project for brain natriuretic peptide (BNP) and the N-terminal part of the propeptide of BNP (NT-proBNP) immunoassays: The Cardio Ormocheck study. Clin. Chem. Lab. Med. 2009, 47, 762–768. [Google Scholar] [CrossRef] [Green Version]
- Clerico, A.; Zaninotto, M.; Prontera, C.; Giovannini, S.; Ndreu, R.; Franzini, M.; Zucchelli, G.C.; Plebani, M.; Study Group on Cardiovascular Risk Biomarkers of the Italian Society of Clinical Biochemistry. State of the art of BNP and NT-proBNP immunoassays: The CardioOrmoCheck study. Clin. Chim. Acta 2012, 414, 112–119. [Google Scholar] [CrossRef]
- Semenov, A.G.; Tamm, N.N.; Apple, F.S.; Schulz, K.M.; Love, S.A.; Ler, R.; Feygina, E.E.; Katrukha, A.G. Searching for a BNP standard: Glycosylated proBNP as a common calibrator enables improved comparability of commercial BNP immunoassays. Clin. Biochem. 2017, 50, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, N.E.; McCarthy, C.P.; Shrestha, S.; Gaggin, H.K.; Mukai, R.; Szymonifka, J.; Apple, F.S.; Burnett, J.C., Jr.; Iyer, S.; Januzzi, J.L., Jr. Effect of Neprilysin Inhibition on Various Natriuretic Peptide Assays. J. Am. Coll. Cardiol. 2019, 73, 1273–1284. [Google Scholar] [CrossRef]
- Luckenbill, K.N.; Christenson, R.H.; Jaffe, A.S.; Mair, J.; Ordonez-Llanos, J.; Pagani, F.; Tate, J.; Wu, A.H.; Ler, R.; Apple, F.S. Cross-reactivity of BNP, NT-proBNP, and proBNP in commercial BNP and NT-proBNP assays: Preliminary observations from the IFCC Committee for Standardization of Markers of Cardiac Damage. Clin. Chem. 2008, 54, 619–621. [Google Scholar] [CrossRef]
- Nishikimi, T.; Minamino, N.; Horii, K.; Matsuoka, H. Do commercially available assay kits for B-type natriuretic peptide measure Pro-BNP1-108, as well as BNP1-32? Hypertension 2007, 50, e163. [Google Scholar] [CrossRef]
- Semenov, A.G.; Seferian, K.R. Biochemistry of the human B-type natriuretic peptide precursor and molecular aspects of its processing. Clin. Chim. Acta 2011, 412, 850–860. [Google Scholar] [CrossRef]
- Niederkofler, E.E.; Kiernan, U.A.; O’Rear, J.; Menon, S.; Saghir, S.; Protter, A.A.; Nelson, R.W.; Schellenberger, U. Detection of endogenous B-type natriuretic peptide at very low concentrations in patients with heart failure. Circ. Heart Fail. 2008, 1, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.L.; Phelps, M.A.; Wood, C.M.; Schellenberger, U.; Van Le, A.; Perichon, R.; Jaffe, A.S. Comparison of mass spectrometry and clinical assay measurements of circulating fragments of B-type natriuretic peptide in patients with chronic heart failure. Circ. Heart Fail. 2011, 4, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Vodovar, N.; Séronde, M.F.; Laribi, S.; Gayat, E.; Lassus, J.; Boukef, R.; Nouira, S.; Manivet, P.; Samuel, J.L.; Logeart, D.; et al. Post-translational modifications enhance NT-proBNP and BNP production in acute decompensated heart failure. Eur. Heart J. 2014, 35, 3434–3441. [Google Scholar] [CrossRef] [Green Version]
- Takahama, H.; Takashio, S.; Nishikimi, T.; Hayashi, T.; Nagai-Okatani, C.; Nakagawa, Y.; Amaki, M.; Ohara, T.; Hasegawa, T.; Sugano, Y.; et al. Ratio of pro-B-type natriuretic peptide (BNP) to total BNP is decreased in mild, but not severe, acute decompensated heart failure patients: A novel compensatory mechanism for acute heart failure. Int. J. Cardiol. 2018, 258, 165–171. [Google Scholar] [CrossRef]
- Kimura, A.; Takahama, H.; Nishikimi, T.; Takashio, S.; Hayashi, T.; Nagai-Okatani, C.; Nakagawa, Y.; Yasuda, S.; Anzai, T.; Minamino, N.; et al. Molecular ratio of mature B-type natriuretic peptide in acute heart failure: An indicator for ventricular contractile recovery. ESC Heart Fail. 2021, 8, 5617–5621. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunoassay/Instrument | Capture Antibody | Detection Antibody | Standard |
|---|---|---|---|
| Abbott Architect BNP | Mouse monoclonal antibody aa5–13 | Mouse monoclonal antibody aa26–32 | Synthetic BNP-32 |
| Alere/Quidel Triage BNP | Mouse monoclonal antibody aa5–13 | Omniclonal antibody (epitope not characterized) | Recombinant BNP-32 |
| Siemens Centaur BNP | Mouse monoclonal antibody aa27–32 | Mouse monoclonal antibody aa14–21 | Synthetic BNP-32 |
| Pylon Single Epitope BNP | Mouse monoclonal antibody aa11–17 | Recognizes the immune complex of capture antibody with BNP/proBNP | Glycosylated proBNP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishikimi, T.; Nakagawa, Y. B-Type Natriuretic Peptide (BNP) Revisited—Is BNP Still a Biomarker for Heart Failure in the Angiotensin Receptor/Neprilysin Inhibitor Era? Biology 2022, 11, 1034. https://doi.org/10.3390/biology11071034
Nishikimi T, Nakagawa Y. B-Type Natriuretic Peptide (BNP) Revisited—Is BNP Still a Biomarker for Heart Failure in the Angiotensin Receptor/Neprilysin Inhibitor Era? Biology. 2022; 11(7):1034. https://doi.org/10.3390/biology11071034
Chicago/Turabian StyleNishikimi, Toshio, and Yasuaki Nakagawa. 2022. "B-Type Natriuretic Peptide (BNP) Revisited—Is BNP Still a Biomarker for Heart Failure in the Angiotensin Receptor/Neprilysin Inhibitor Era?" Biology 11, no. 7: 1034. https://doi.org/10.3390/biology11071034