
Neuregulin (NRG-1β) Is Pro-Myogenic and Anti-Cachectic in Respiratory Muscles of Post-Myocardial Infarcted Swine

 ,
,
Abstract
:Simple Summary
Abstract
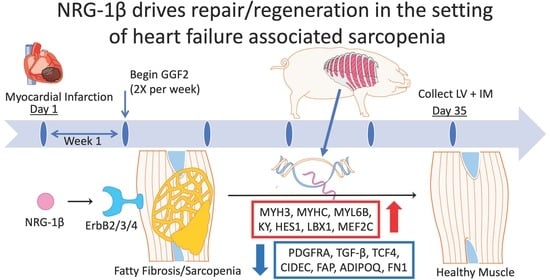
1. Introduction
2. Materials and Methods
3. Results
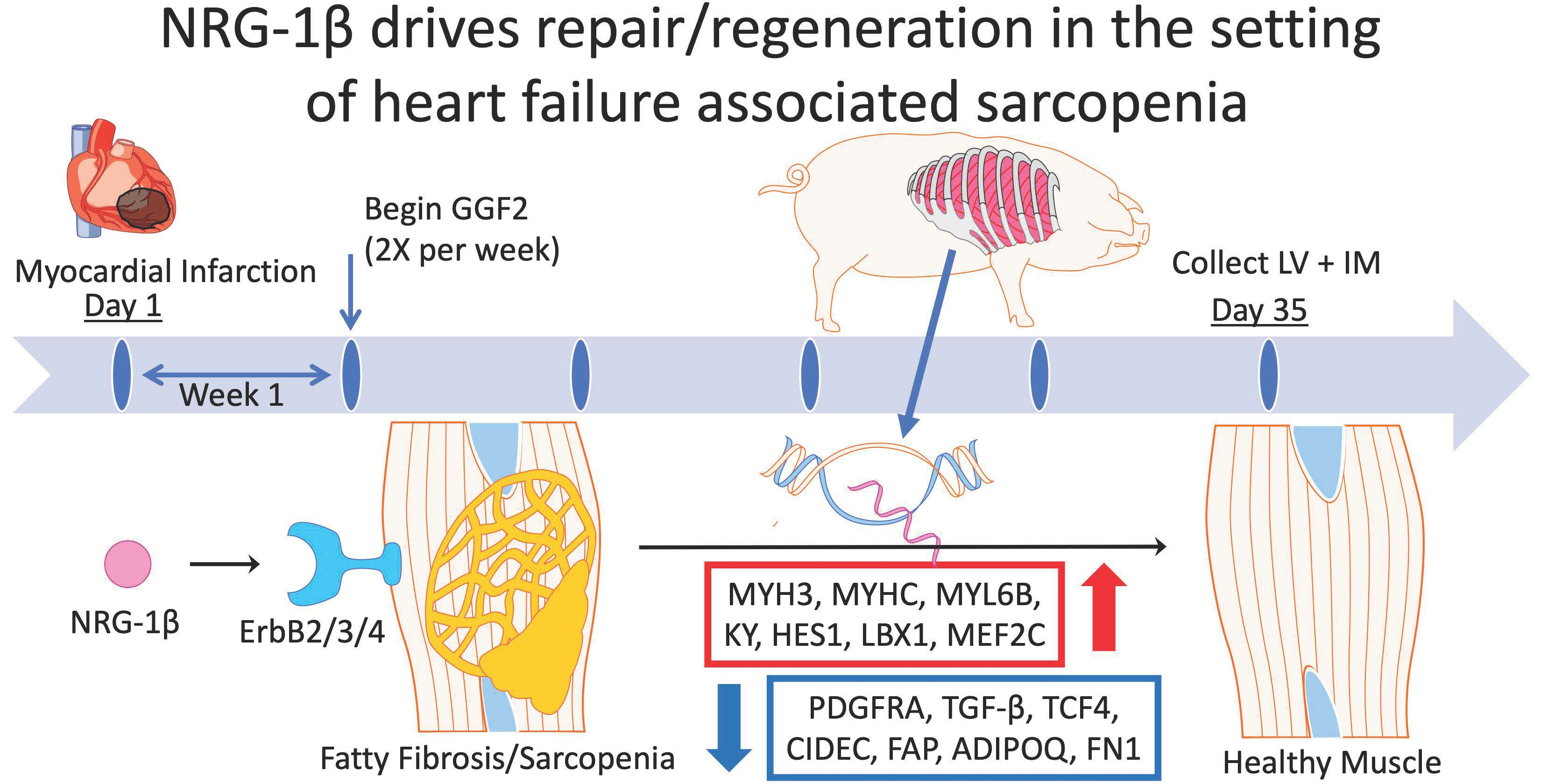
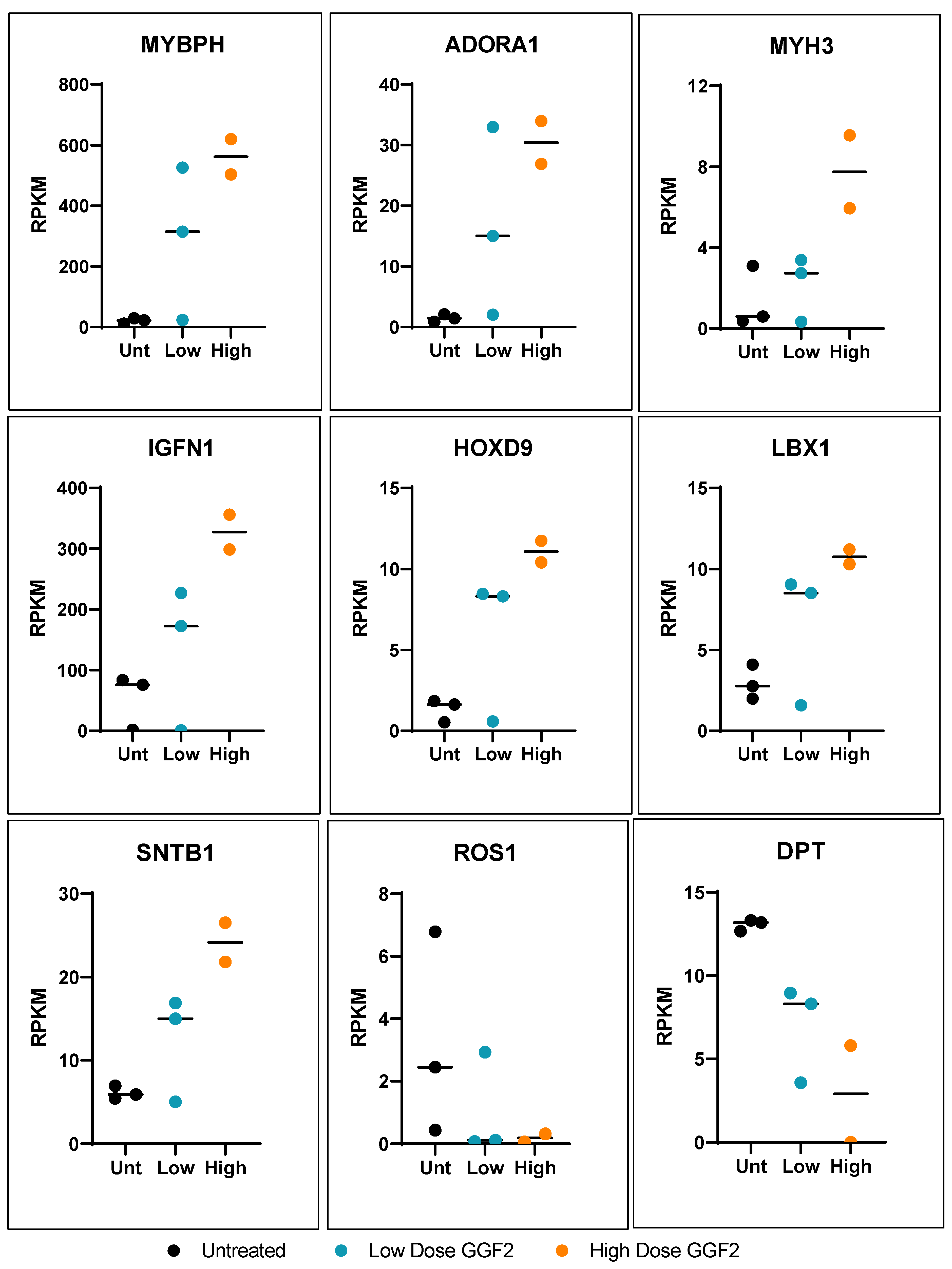
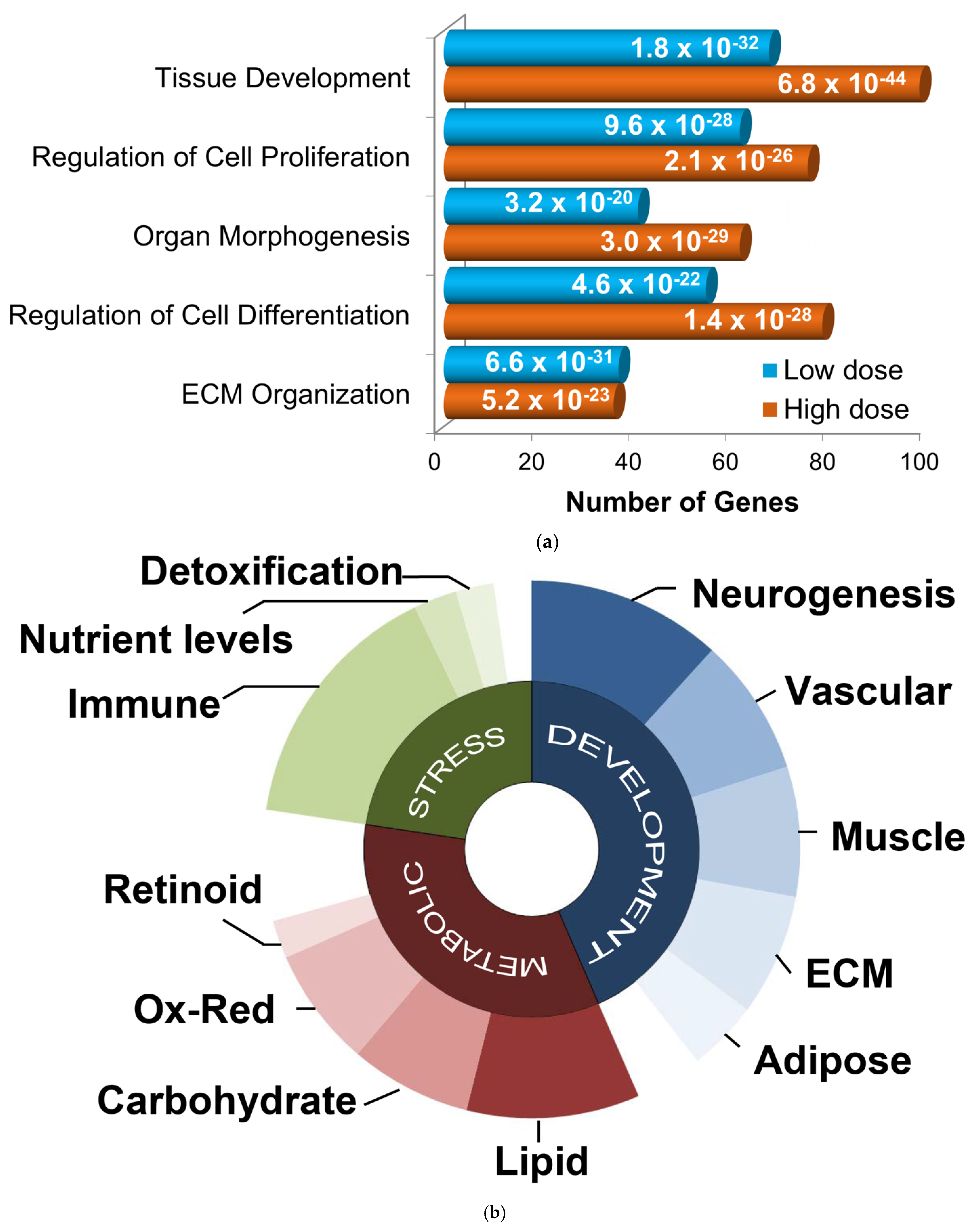
3.1. GGF2 Alters Intercostal Muscle Gene Expression
3.2. Dose-Dependent Effects of GGF2
3.3. Functional Enrichment of GGF2 Altered Genes
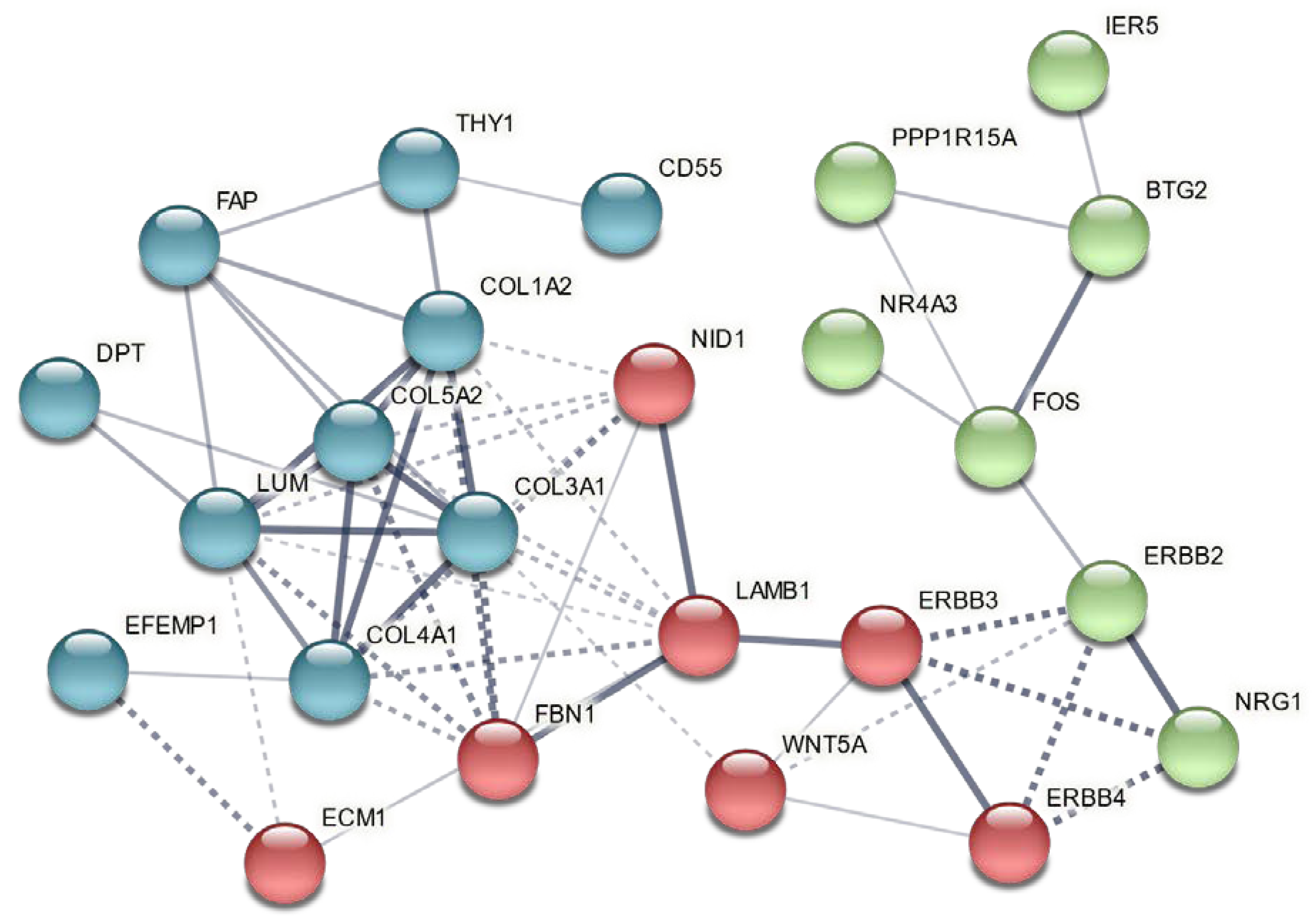
3.4. Genes Altered in Skeletal and Cardiac Muscles of GGF2-Treated Post-MI Pigs
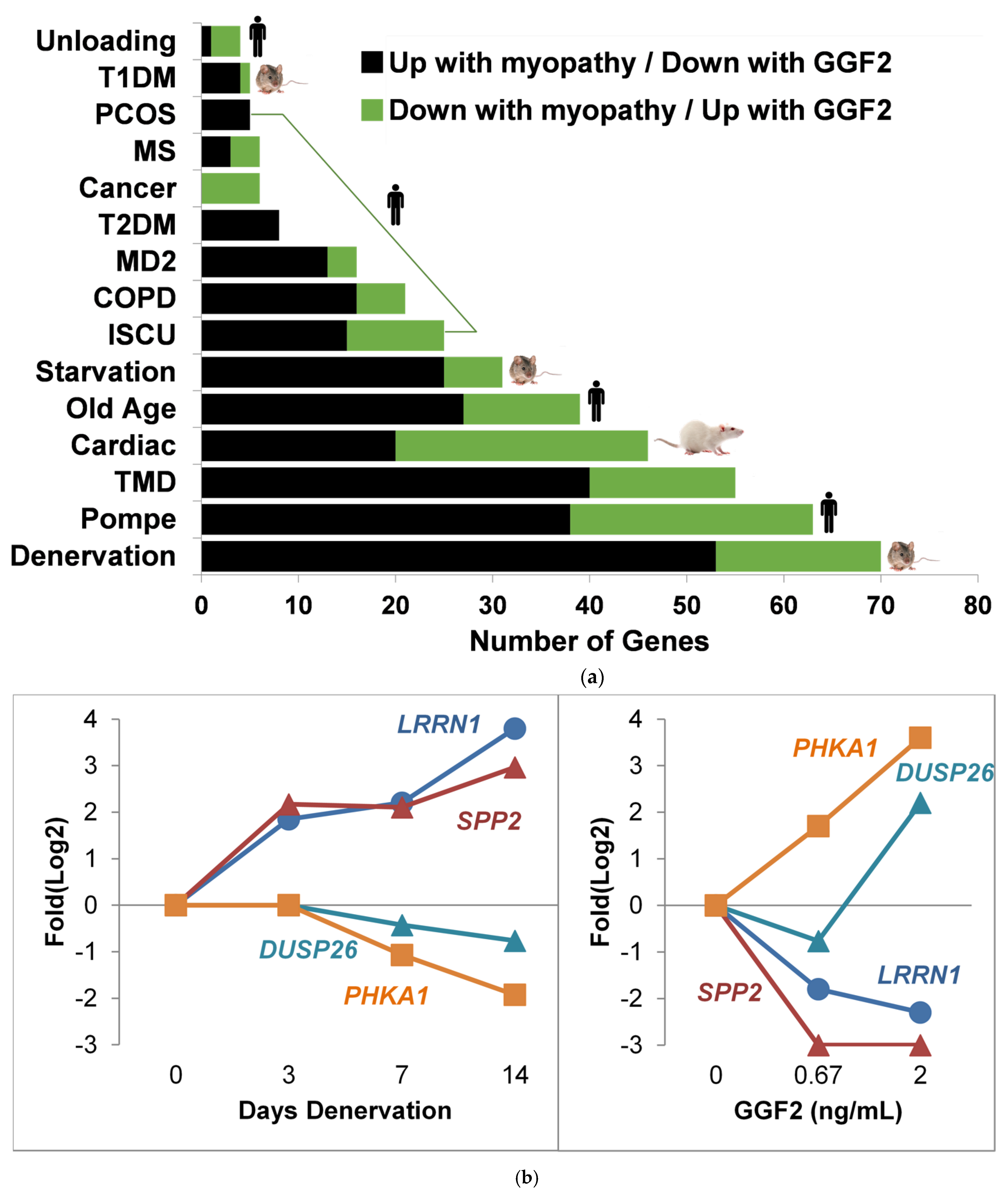
3.5. Comparison to Skeletal Muscle Myopathies
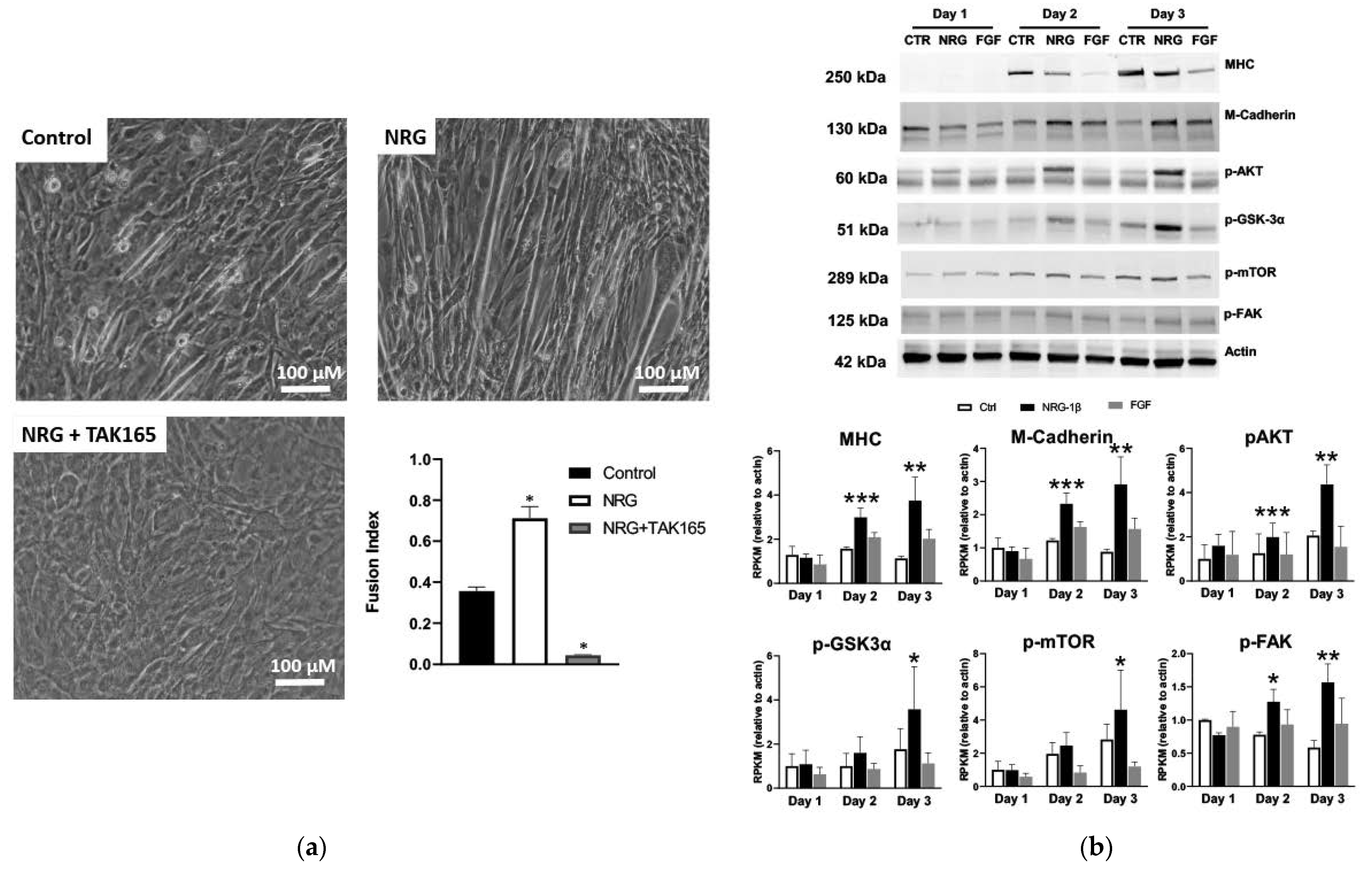
3.6. GGF2/NRG-1β Stimulates ErbB2-Dependent Myoblast Differentiation
3.7. A Putative GGF2-Induced Skeletal Muscle Signaling Pathway
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kharraz, Y.; Guerra, J.; Pessina, P.; Serrano, A.L.; Muñoz-Cánoves, P. Understanding the Process of Fibrosis in Duchenne Muscular Dystrophy. BioMed Res. Int. 2014, 2014, 965631. [Google Scholar] [CrossRef] [PubMed]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef] [PubMed]
- Seene, T.; Kaasik, P. Muscle weakness in the elderly: Role of sarcopenia, dynapenia, and possibilities for rehabilitation. Eur. Rev. Aging Phys. Act. 2012, 9, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Yokota, T.; Kinugawa, S.; Hirabayashi, K.; Yamato, M.; Takada, S.; Suga, T.; Nakano, I.; Fukushima, A.; Matsushima, S.; Okita, K.; et al. Systemic oxidative stress is associated with lower aerobic capacity and impaired skeletal muscle energy metabolism in heart failure patients. Sci. Rep. 2021, 11, 2272. [Google Scholar] [CrossRef] [PubMed]
- Zizola, C.; Schulze, P.C. Metabolic and structural impairment of skeletal muscle in heart failure. Heart Fail. Rev. 2013, 18, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Keller-Ross, M.L.; Larson, M.; Johnson, B.D. Skeletal Muscle Fatigability in Heart Failure. Front. Physiol. 2019, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Kinugawa, S.; Takada, S.; Matsushima, S.; Okita, K.; Tsutsui, H. Skeletal Muscle Abnormalities in Heart Failure. Int. Heart J. 2015, 56, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Maskin, C.S.; Forman, R.; Sonnenblick, E.H.; Frishman, W.H.; LeJemtel, T.H. Failure of dobutamine to increase exercise capacity despite hemodynamic improvement in severe chronic heart failure. Am. J. Cardiol. 1983, 51, 177–182. [Google Scholar] [CrossRef]
- Mancini, D.M.; Coyle, E.; Coggan, A.; Beltz, J.; Ferraro, N.; Montain, S.; Wilson, J.R. Contribution of intrinsic skeletal muscle changes to 31P NMR skeletal muscle metabolic abnormalities in patients with chronic heart failure. Circulation 1989, 80, 1338–1346. [Google Scholar] [CrossRef] [Green Version]
- Britsch, S.; Li, L.; Kirchhoff, S.; Theuring, F.; Brinkmann, V.; Birchmeier, C.; Riethmacher, D. The ErbB2 and ErbB3 receptors and their ligand, neuregulin-1, are essential for development of the sympathetic nervous system. Genes Dev. 1998, 12, 1825–1836. [Google Scholar] [CrossRef] [Green Version]
- Sandrock, A.W., Jr.; Goodearl, A.D.; Yin, Q.W.; Chang, D.; Fischbach, G.D. ARIA is concentrated in nerve terminals at neuromuscular junctions and at other synapses. J. Neurosci. 1995, 15, 6124–6136. [Google Scholar] [CrossRef] [PubMed]
- Florini, J.R.; Samuel, D.S.; Ewton, D.Z.; Kirk, C.; Sklar, R.M. Stimulation of myogenic differentiation by a neuregulin, glial growth factor 2. Are neuregulins the long-sought muscle trophic factors secreted by nerves? J. Biol. Chem. 1996, 271, 12699–12702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Chi, S.; Lee, K.H.; Rhee, S.; Kwon, Y.K.; Chung, C.H.; Kwon, H.; Kang, M.S. Neuregulin stimulates myogenic differentiation in an autocrine manner. J. Biol. Chem. 1999, 274, 15395–15400. [Google Scholar] [CrossRef] [Green Version]
- Ford, B.D.; Han, B.; Fischbach, G.D. Differentiation-dependent regulation of skeletal myogenesis by neuregulin-1. Biochem. Biophys. Res. Commun. 2003, 306, 276–281. [Google Scholar] [CrossRef]
- Jo, S.A.; Zhu, X.; Marchionni, M.A.; Burden, S.J. Neuregulins are concentrated at nerve-muscle synapses and activate ACh–receptor gene expression. Nature 1995, 373, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Van Ho, A.T.; Hayashi, S.; Brohl, D.; Aurade, F.; Rattenbach, R.; Relaix, F. Neural crest cell lineage restricts skeletal muscle progenitor cell differentiation through Neuregulin1-ErbB3 signaling. Dev. Cell 2011, 21, 273–287. [Google Scholar] [CrossRef]
- Galindo, C.L.; Kasasbeh, E.; Murphy, A.; Ryzhov, S.; Lenihan, S.; Ahmad, F.A.; Williams, P.; Nunnally, A.; Adcock, J.; Song, Y.; et al. Anti-remodeling and anti-fibrotic effects of the neuregulin-1beta glial growth factor 2 in a large animal model of heart failure. J. Am. Heart Assoc. 2014, 3, e000773. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Sawyer, D.B.; Troy, E.L.; McEwen, C.; Cleator, J.H.; Murphy, A.; Caggiano, A.O.; Eisen, A.; Parry, T.J. Species-specific effects of neuregulin-1beta (cimaglermin alfa) on glucose handling in animal models and humans with heart failure. Toxicol. Appl. Pharmacol. 2017, 332, 92–99. [Google Scholar] [CrossRef]
- Parry, T.J.; Ganguly, A.; Troy, E.L.; Luis Guerrero, J.; Iaci, J.F.; Srinivas, M.; Vecchione, A.M.; Button, D.C.; Hackett, C.S.; Zolty, R.; et al. Effects of neuregulin GGF2 (cimaglermin alfa) dose and treatment frequency on left ventricular function in rats following myocardial infarction. Eur. J. Pharmacol. 2017, 796, 76–89. [Google Scholar] [CrossRef]
- Liu, Q.; Zhu, X.; Xu, L.; Fu, Y.; Garvey, W.T. 6-Mercaptopurine augments glucose transport activity in skeletal muscle cells in part via a mechanism dependent upon orphan nuclear receptor NR4A3. Am. J. Physiol.-Endocrinol. Metab. 2013, 305, E1081–E1092. [Google Scholar] [CrossRef] [Green Version]
- Pearen, M.A.; Ryall, J.G.; Maxwell, M.A.; Ohkura, N.; Lynch, G.S.; Muscat, G.E. The orphan nuclear receptor, NOR-1, is a target of beta-adrenergic signaling in skeletal muscle. Endocrinology 2006, 147, 5217–5227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Luo, C.; Zhu, T.; Li, L.; Zhang, W. Elevated c-fos expression is correlated with phenotypic switching of human vascular smooth muscle cells derived from lower limb venous varicosities. J. Vasc. Surg. Venous Lymphat. Disord. 2021, 9, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Trouche, D.; Grigoriev, M.; Lenormand, J.-L.; Robin, P.; Alexandre Leibovitch, S.; Sassone-Corsi, P.; Harel-Bellan, A. Repression of c-fos promoter by MyoD on muscle cell differentiation. Nature 1993, 363, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Ouyang, Y.; Bai, Y.; Gong, J.; Liao, H. miR-155-5p inhibits the viability of vascular smooth muscle cell via targeting FOS and ZIC3 to promote aneurysm formation. Eur. J. Pharmacol. 2019, 853, 145–152. [Google Scholar] [CrossRef]
- Shah, V.O.; Dominic, E.A.; Moseley, P.; Pickett, G.; Fleet, M.; Ness, S.; Raj, D.S.C. Hemodialysis Modulates Gene Expression Profile in Skeletal Muscle. Am. J. Kidney Dis. 2006, 48, 616–628. [Google Scholar] [CrossRef]
- Tamboli, R.A.; Hajri, T.; Jiang, A.; Marks-Shulman, P.A.; Williams, D.B.; Clements, R.H.; Melvin, W.; Bowen, B.P.; Shyr, Y.; Abumrad, N.N.; et al. Reduction in inflammatory gene expression in skeletal muscle from Roux-en-Y gastric bypass patients randomized to omentectomy. PLoS ONE 2011, 6, e28577. [Google Scholar] [CrossRef] [Green Version]
- Pestov, N.B.; Adams, G.; Shakhparonov, M.I.; Modyanov, N.N. Identification of a novel gene of the X,K-ATPase beta-subunit family that is predominantly expressed in skeletal and heart muscles. FEBS Lett. 1999, 456, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Pestov, N.B.; Zhao, H.; Basrur, V.; Modyanov, N.N. Isolation and characterization of BetaM protein encoded by ATP1B4—A unique member of the Na,K-ATPase β-subunit gene family. Biochem. Biophys. Res. Commun. 2011, 412, 543–548. [Google Scholar] [CrossRef]
- Roediger, M.; Miosge, N.; Gersdorff, N. Tissue distribution of the laminin β1 and β2 chain during embryonic and fetal human development. J. Mol. Histol. 2010, 41, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Radmanesh, F.; Caglayan, A.O.; Silhavy, J.L.; Yilmaz, C.; Cantagrel, V.; Omar, T.; Rosti, B.; Kaymakcalan, H.; Gabriel, S.; Li, M.; et al. Mutations in LAMB1 cause cobblestone brain malformation without muscular or ocular abnormalities. Am. J. Hum. Genet. 2013, 92, 468–474. [Google Scholar] [CrossRef] [Green Version]
- Engebretsen, K.V.; Lunde, I.G.; Strand, M.E.; Waehre, A.; Sjaastad, I.; Marstein, H.S.; Skrbic, B.; Dahl, C.P.; Askevold, E.T.; Christensen, G.; et al. Lumican is increased in experimental and clinical heart failure, and its production by cardiac fibroblasts is induced by mechanical and proinflammatory stimuli. FEBS J. 2013, 280, 2382–2398. [Google Scholar] [CrossRef] [PubMed]
- Mohammadzadeh, N.; Lunde, I.G.; Andenæs, K.; Strand, M.E.; Aronsen, J.M.; Skrbic, B.; Marstein, H.S.; Bandlien, C.; Nygård, S.; Gorham, J.; et al. The extracellular matrix proteoglycan lumican improves survival and counteracts cardiac dilatation and failure in mice subjected to pressure overload. Sci. Rep. 2019, 9, 9206. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, D.; Mao, L.; Abraham, D.M.; Bursac, N. Lack of Thy1 defines a pathogenic fraction of cardiac fibroblasts in heart failure. Biomaterials 2020, 236, 119824. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Ibrahim, A.; Hensley, M.T.; Shen, D.; Sun, B.; Middleton, R.; Liu, W.; Smith, R.R.; Marbán, E. Relative roles of CD90 and c-kit to the regenerative efficacy of cardiosphere-derived cells in humans and in a mouse model of myocardial infarction. J. Am. Heart Assoc. 2014, 3, e001260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamada, T.; Wessagowit, V.; South, A.P.; Ashton, G.H.; Chan, I.; Oyama, N.; Siriwattana, A.; Jewhasuchin, P.; Charuwichitratana, S.; Thappa, D.M.; et al. Extracellular matrix protein 1 gene (ECM1) mutations in lipoid proteinosis and genotype-phenotype correlation. J. Investig. Dermatol. 2003, 120, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Hardy, S.A.; Mabotuwana, N.S.; Murtha, L.A.; Coulter, B.; Sanchez-Bezanilla, S.; Al-Omary, M.S.; Senanayake, T.; Loering, S.; Starkey, M.; Lee, R.J.; et al. Novel role of extracellular matrix protein 1 (ECM1) in cardiac aging and myocardial infarction. PLoS ONE 2019, 14, e0212230. [Google Scholar] [CrossRef] [Green Version]
- Uitte de Willige, S.; Keane, F.M.; Bowen, D.G.; Malfliet, J.; Zhang, H.E.; Maneck, B.; McCaughan, G.W.; Leebeek, F.W.G.; Rijken, D.C.; Gorrell, M.D. Circulating fibroblast activation protein activity and antigen levels correlate strongly when measured in liver disease and coronary heart disease. PLoS ONE 2017, 12, e0178987. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Cheng, X.; Rong, R.; Gao, Y.; Tang, X.; Chen, Y. High expression of fibroblast activation protein (FAP) predicts poor outcome in high-grade serous ovarian cancer. BMC Cancer 2020, 20, 1032. [Google Scholar] [CrossRef]
- Solano-Iturri, J.D.; Beitia, M.; Errarte, P.; Calvete-Candenas, J.; Etxezarraga, M.C.; Loizate, A.; Echevarria, E.; Badiola, I.; Larrinaga, G. Altered expression of fibroblast activation protein-α (FAP) in colorectal adenoma-carcinoma sequence and in lymph node and liver metastases. Aging 2020, 12, 10337–10358. [Google Scholar] [CrossRef]
- Kim, T.; Ahmad, K.; Shaikh, S.; Jan, A.T.; Seo, M.-G.; Lee, E.J.; Choi, I. Dermatopontin in Skeletal Muscle Extracellular Matrix Regulates Myogenesis. Cells 2019, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Bastos, A.N.; Alves, M.M.; Monte-Alto-Costa, A.; Machado, D.G.; Cavalcante, G.J.; Panico, M.; Porto, L.C. α-smooth muscle actin, fibrillin-1, apoptosis and proliferation detection in primary varicose lower limb veins of women. Int. Angiol. 2011, 30, 262–271. [Google Scholar] [PubMed]
- Chen, L.; Ge, Q.; Black, J.L.; Deng, L.; Burgess, J.K.; Oliver, B.G. Differential regulation of extracellular matrix and soluble fibulin-1 levels by TGF-β₁ in airway smooth muscle cells. PLoS ONE 2013, 8, e65544. [Google Scholar] [CrossRef] [PubMed]
- Franken, R.; Teixido-Tura, G.; Brion, M.; Forteza, A.; Rodriguez-Palomares, J.; Gutierrez, L.; Garcia Dorado, D.; Pals, G.; Mulder, B.J.; Evangelista, A. Relationship between fibrillin-1 genotype and severity of cardiovascular involvement in Marfan syndrome. Heart 2017, 103, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- McNiven, V.; Ito, Y.A.; Hartley, T.; Kernohan, K.; Miller, E.; Armour, C.M. NID1 variant associated with occipital cephaloceles in a family expressing a spectrum of phenotypes. Am. J. Med. Genet. Part A 2019, 179, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, K.; Hoischen, S.; Hafner, M.; Nischt, R. Genomic sequences and structural organization of the human nidogen gene (NID). Genomics 1995, 27, 245–250. [Google Scholar] [CrossRef]
- Kim, E.H.; Galchev, V.I.; Kim, J.Y.; Misek, S.A.; Stevenson, T.K.; Campbell, M.D.; Pagani, F.D.; Day, S.M.; Johnson, T.C.; Washburn, J.G.; et al. Differential protein expression and basal lamina remodeling in human heart failure. Proteom. Clin. Appl. 2016, 10, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Wadley, G.D.; Lamon, S.; Alexander, S.E.; McMullen, J.R.; Bernardo, B.C. Noncoding RNAs regulating cardiac muscle mass. J. Appl. Physiol. 2018, 127, 633–644. [Google Scholar] [CrossRef]
- Chen, L.; Bai, J.; Li, Y. miR-29 mediates exercise-induced skeletal muscle angiogenesis by targeting VEGFA, COL4A1 and COL4A2 via the PI3K/Akt signaling pathway. Mol. Med. Rep. 2020, 22, 661–670. [Google Scholar] [CrossRef]
- Labelle-Dumais, C.; Schuitema, V.; Hayashi, G.; Hoff, K.; Gong, W.; Dao, D.Q.; Ullian, E.M.; Oishi, P.; Margeta, M.; Gould, D.B. COL4A1 Mutations Cause Neuromuscular Disease with Tissue-Specific Mechanistic Heterogeneity. Am. J. Hum. Genet. 2019, 104, 847–860. [Google Scholar] [CrossRef] [Green Version]
- Meienberg, J.; Rohrbach, M.; Neuenschwander, S.; Spanaus, K.; Giunta, C.; Alonso, S.; Arnold, E.; Henggeler, C.; Regenass, S.; Patrignani, A.; et al. Hemizygous deletion of COL3A1, COL5A2, and MSTN causes a complex phenotype with aortic dissection: A lesson for and from true haploinsufficiency. Eur. J. Hum. Genet. 2010, 18, 1315–1321. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.Y.; Shi, M.J.; Zeng, Z.H.; Chen, C.; Liu, T.Z.; Wu, Q.J.; Li, S.; Li, S. The Role of COL5A2 in Patients with Muscle-Invasive Bladder Cancer: A Bioinformatics Analysis of Public Datasets Involving 787 Subjects and 29 Cell Lines. Front. Oncol. 2018, 8, 659. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, K.; Zabojszcza, J.; Carl, M.; Taubert, S.; Lass, A.; Harris, C.L.; Ho, M.; Schulz, H.; Hummel, O.; Hubner, N.; et al. Increased susceptibility to complement attack due to down-regulation of decay-accelerating factor/CD55 in dysferlin-deficient muscular dystrophy. J. Immunol. 2005, 175, 6219–6225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuivaniemi, H.; Tromp, G. Type III collagen (COL3A1): Gene and protein structure, tissue distribution, and associated diseases. Gene 2019, 707, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Ghavampur, S.; Bondallaz, P.; Will, L.; Grenningloh, G.; Pu Schel, A.W. Rnd1 regulates axon extension by enhancing the microtubule destabilizing activity of SCG10. J. Biol. Chem. 2009, 284, 363–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Liu, S.; Zhao, Z.; Zhang, Y.; Yang, Y.; Luo, B. Relationship between PPP1R15A gene polymorphism (rs611251) and Epstein-Barr virus-associated tumors. Acta Virol. 2017, 61, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Sakurai, H. Heat-induced expression of the immediate-early gene IER5 and its involvement in the proliferation of heat-shocked cells. FEBS J. 2015, 282, 332–340. [Google Scholar] [CrossRef]
- Nakamura, M.; Basavarajaiah, P.; Rousset, E.; Beraud, C.; Latreille, D.; Henaoui, I.S.; Lassot, I.; Mari, B.; Kiernan, R. Spt6 levels are modulated by PAAF1 and proteasome to regulate the HIV-1 LTR. Retrovirology 2012, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Luo, C.; Yin, Z.; Li, P.; Wang, S.; Chen, J.; He, Q.; Zhou, J. Downregulation of let-7b promotes COL1A1 and COL1A2 expression in dermis and skin fibroblasts during heat wound repair. Mol. Med. Rep. 2016, 13, 2683–2688. [Google Scholar] [CrossRef]
- Camaj, P.; Seeliger, H.; Ischenko, I.; Krebs, S.; Blum, H.; De Toni, E.N.; Faktorova, D.; Jauch, K.W.; Bruns, C.J. EFEMP1 binds the EGF receptor and activates MAPK and Akt pathways in pancreatic carcinoma cells. Biol. Chem. 2009, 390, 1293–1302. [Google Scholar] [CrossRef] [Green Version]
- Lei, B.; Wan, B.; Peng, J.; Yang, Y.; Lv, D.; Zhou, X.; Shu, F.; Li, F.; Zhong, L.; Wu, H.; et al. PRPS2 Expression Correlates with Sertoli-Cell Only Syndrome and Inhibits the Apoptosis of TM4 Sertoli Cells. J. Urol. 2015, 194, 1491–1497. [Google Scholar] [CrossRef]
- Miao, W.; Wang, Y. Targeted Quantitative Kinome Analysis Identifies PRPS2 as a Promoter for Colorectal Cancer Metastasis. J. Proteome Res. 2019, 18, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Song, J.; Ernst, P.; Latimer, M.N.; Ha, C.-M.; Goh, K.Y.; Ma, W.; Rajasekaran, N.-S.; Zhang, J.; Liu, X.; et al. MitoQ regulates redox-related noncoding RNAs to preserve mitochondrial network integrity in pressure-overload heart failure. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H682–H695. [Google Scholar] [CrossRef] [PubMed]
- Son, G.H.; Park, S.H.; Kim, Y.; Kim, J.Y.; Kim, J.W.; Chung, S.; Kim, Y.H.; Kim, H.; Hwang, J.J.; Seo, J.S. Postmortem mRNA expression patterns in left ventricular myocardial tissues and their implications for forensic diagnosis of sudden cardiac death. Mol. Cells 2014, 37, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Meier, J.J. GLP-1 receptor agonists in the treatment of type 2 diabetes–state-of-the-art. Mol. Metab. 2021, 46, 101102. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhong, H.; Qin, Y.; Wei, W.; Li, Z.; Huang, M.; Luo, X. ARMCX Family Gene Expression Analysis and Potential Prognostic Biomarkers for Prediction of Clinical Outcome in Patients with Gastric Carcinoma. BioMed Res. Int. 2020, 2020, 3575038. [Google Scholar] [CrossRef]
- Wellner, M.; Dechend, R.; Park, J.K.; Shagdarsuren, E.; Al-Saadi, N.; Kirsch, T.; Gratze, P.; Schneider, W.; Meiners, S.; Fiebeler, A.; et al. Cardiac gene expression profile in rats with terminal heart failure and cachexia. Physiol. Genom. 2005, 20, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Soares, R.J.; Cagnin, S.; Chemello, F.; Silvestrin, M.; Musaro, A.; De Pitta, C.; Lanfranchi, G.; Sandri, M. Involvement of microRNAs in the regulation of muscle wasting during catabolic conditions. J. Biol. Chem. 2014, 289, 21909–21925. [Google Scholar] [CrossRef] [Green Version]
- Screen, M.; Jonson, P.H.; Raheem, O.; Palmio, J.; Laaksonen, R.; Lehtimaki, T.; Sirito, M.; Krahe, R.; Hackman, P.; Udd, B. Abnormal splicing of NEDD4 in myotonic dystrophy type 2: Possible link to statin adverse reactions. Am. J. Pathol. 2014, 184, 2322–2332. [Google Scholar] [CrossRef] [Green Version]
- Crooks, D.R.; Natarajan, T.G.; Jeong, S.Y.; Chen, C.; Park, S.Y.; Huang, H.; Ghosh, M.C.; Tong, W.H.; Haller, R.G.; Wu, C.; et al. Elevated FGF21 secretion, PGC-1alpha and ketogenic enzyme expression are hallmarks of iron-sulfur cluster depletion in human skeletal muscle. Hum. Mol. Genet. 2014, 23, 24–39. [Google Scholar] [CrossRef] [Green Version]
- Palermo, A.T.; Palmer, R.E.; So, K.S.; Oba-Shinjo, S.M.; Zhang, M.; Richards, B.; Madhiwalla, S.T.; Finn, P.F.; Hasegawa, A.; Ciociola, K.M.; et al. Transcriptional response to GAA deficiency (Pompe disease) in infantile-onset patients. Mol. Genet. Metab. 2012, 106, 287–300. [Google Scholar] [CrossRef]
- Gallagher, I.J.; Stephens, N.A.; MacDonald, A.J.; Skipworth, R.J.; Husi, H.; Greig, C.A.; Ross, J.A.; Timmons, J.A.; Fearon, K.C. Suppression of skeletal muscle turnover in cancer cachexia: Evidence from the transcriptome in sequential human muscle biopsies. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 2817–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Screen, M.; Raheem, O.; Holmlund-Hampf, J.; Jonson, P.H.; Huovinen, S.; Hackman, P.; Udd, B. Gene expression profiling in tibial muscular dystrophy reveals unfolded protein response and altered autophagy. PLoS ONE 2014, 9, e90819. [Google Scholar] [CrossRef] [PubMed]
- Raue, U.; Trappe, T.A.; Estrem, S.T.; Qian, H.R.; Helvering, L.M.; Smith, R.C.; Trappe, S. Transcriptome signature of resistance exercise adaptations: Mixed muscle and fiber type specific profiles in young and old adults. J. Appl. Physiol. 2012, 112, 1625–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanza, I.R.; Short, D.K.; Short, K.R.; Raghavakaimal, S.; Basu, R.; Joyner, M.J.; McConnell, J.P.; Nair, K.S. Endurance exercise as a countermeasure for aging. Diabetes 2008, 57, 2933–2942. [Google Scholar] [CrossRef] [Green Version]
- Urso, M.L.; Scrimgeour, A.G.; Chen, Y.W.; Thompson, P.D.; Clarkson, P.M. Analysis of human skeletal muscle after 48 h immobilization reveals alterations in mRNA and protein for extracellular matrix components. J. Appl. Physiol. 2006, 101, 1136–1148. [Google Scholar] [CrossRef] [Green Version]
- Reich, K.A.; Chen, Y.W.; Thompson, P.D.; Hoffman, E.P.; Clarkson, P.M. Forty-eight hours of unloading and 24 h of reloading lead to changes in global gene expression patterns related to ubiquitination and oxidative stress in humans. J. Appl. Physiol. 2010, 109, 1404–1415. [Google Scholar] [CrossRef]
- Poelkens, F.; Lammers, G.; Pardoel, E.M.; Tack, C.J.; Hopman, M.T. Upregulation of skeletal muscle inflammatory genes links inflammation with insulin resistance in women with the metabolic syndrome. Exp. Physiol. 2013, 98, 1485–1494. [Google Scholar] [CrossRef]
- Skov, V.; Glintborg, D.; Knudsen, S.; Jensen, T.; Kruse, T.A.; Tan, Q.; Brusgaard, K.; Beck-Nielsen, H.; Hojlund, K. Reduced expression of nuclear-encoded genes involved in mitochondrial oxidative metabolism in skeletal muscle of insulin-resistant women with polycystic ovary syndrome. Diabetes 2007, 56, 2349–2355. [Google Scholar] [CrossRef] [Green Version]
- Skov, V.; Glintborg, D.; Knudsen, S.; Tan, Q.; Jensen, T.; Kruse, T.A.; Beck-Nielsen, H.; Hojlund, K. Pioglitazone enhances mitochondrial biogenesis and ribosomal protein biosynthesis in skeletal muscle in polycystic ovary syndrome. PLoS ONE 2008, 3, e2466. [Google Scholar] [CrossRef]
- Jin, W.; Goldfine, A.B.; Boes, T.; Henry, R.R.; Ciaraldi, T.P.; Kim, E.Y.; Emecan, M.; Fitzpatrick, C.; Sen, A.; Shah, A.; et al. Increased SRF transcriptional activity in human and mouse skeletal muscle is a signature of insulin resistance. J. Clin. Investig. 2011, 121, 918–929. [Google Scholar] [CrossRef] [Green Version]
- Suarez, E.; Bach, D.; Cadefau, J.; Palacin, M.; Zorzano, A.; Guma, A. A novel role of neuregulin in skeletal muscle. Neuregulin stimulates glucose uptake, glucose transporter translocation, and transporter expression in muscle cells. J. Biol. Chem. 2001, 276, 18257–18264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathes, S.; Fahrner, A.; Ghoshdastider, U.; Rudiger, H.A.; Leunig, M.; Wolfrum, C.; Krutzfeldt, J. FGF-2-dependent signaling activated in aged human skeletal muscle promotes intramuscular adipogenesis. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Ma, F.; Zhang, H. Circulating Levels of IL-13, TGF-beta1, and Periostin as Potential Biomarker for Coronary Artery Disease with Acute Heart Failure. Evid.-Based Complement. Altern. Med. 2021, 2021, 1690421. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.Y.; Cai, H.; Qing, H.R.; Li, L.; Zhang, H.P. Recent advances on the role of long non-coding RNA H19 in regulating mammalian muscle growth and development. Yi chuan = Hereditas 2017, 39, 1150–1157. [Google Scholar] [CrossRef]
- Wei, W.; He, H.B.; Zhang, W.Y.; Zhang, H.X.; Bai, J.B.; Liu, H.Z.; Cao, J.H.; Chang, K.C.; Li, X.Y.; Zhao, S.H. miR-29 targets Akt3 to reduce proliferation and facilitate differentiation of myoblasts in skeletal muscle development. Cell Death Dis. 2013, 4, e668. [Google Scholar] [CrossRef] [Green Version]
- Greco, S.; De Simone, M.; Colussi, C.; Zaccagnini, G.; Fasanaro, P.; Pescatori, M.; Cardani, R.; Perbellini, R.; Isaia, E.; Sale, P.; et al. Common micro-RNA signature in skeletal muscle damage and regeneration induced by Duchenne muscular dystrophy and acute ischemia. FASEB J. 2009, 23, 3335–3346. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Jiao, Y.; Zhu, Y.; Ning, Z.; Ye, Z.; Li, Q.X.; Hu, C.; Wang, C. Putative MicroRNA-mRNA Networks Upon Mdfi Overexpression in C2C12 Cell Differentiation and Muscle Fiber Type Transformation. Front. Mol. Biosci. 2021, 8, 675993. [Google Scholar] [CrossRef]
- Qaisar, R.; Karim, A.; Muhammad, T.; Shah, I.; Khan, J. Circulating MicroRNAs as Biomarkers of Accelerated Sarcopenia in Chronic Heart Failure. Glob. Heart 2021, 16, 56. [Google Scholar] [CrossRef]
- Yanai, K.; Kaneko, S.; Ishii, H.; Aomatsu, A.; Ito, K.; Hirai, K.; Ookawara, S.; Ishibashi, K.; Morishita, Y. MicroRNAs in Sarcopenia: A Systematic Review. Front. Med. 2020, 7, 180. [Google Scholar] [CrossRef]
- Sun, M.; Guo, M.; Ma, G.; Zhang, N.; Pan, F.; Fan, X.; Wang, R. MicroRNA-30c-5p protects against myocardial ischemia/reperfusion injury via regulation of Bach1/Nrf2. Toxicol. Appl. Pharmacol. 2021, 426, 115637. [Google Scholar] [CrossRef]
- Meng, S.; Hu, Y.; Zhu, J.; Feng, T.; Quan, X. miR-30c-5p acts as a therapeutic target for ameliorating myocardial ischemia-reperfusion injury. Am. J. Transl. Res. 2021, 13, 2198–2212. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
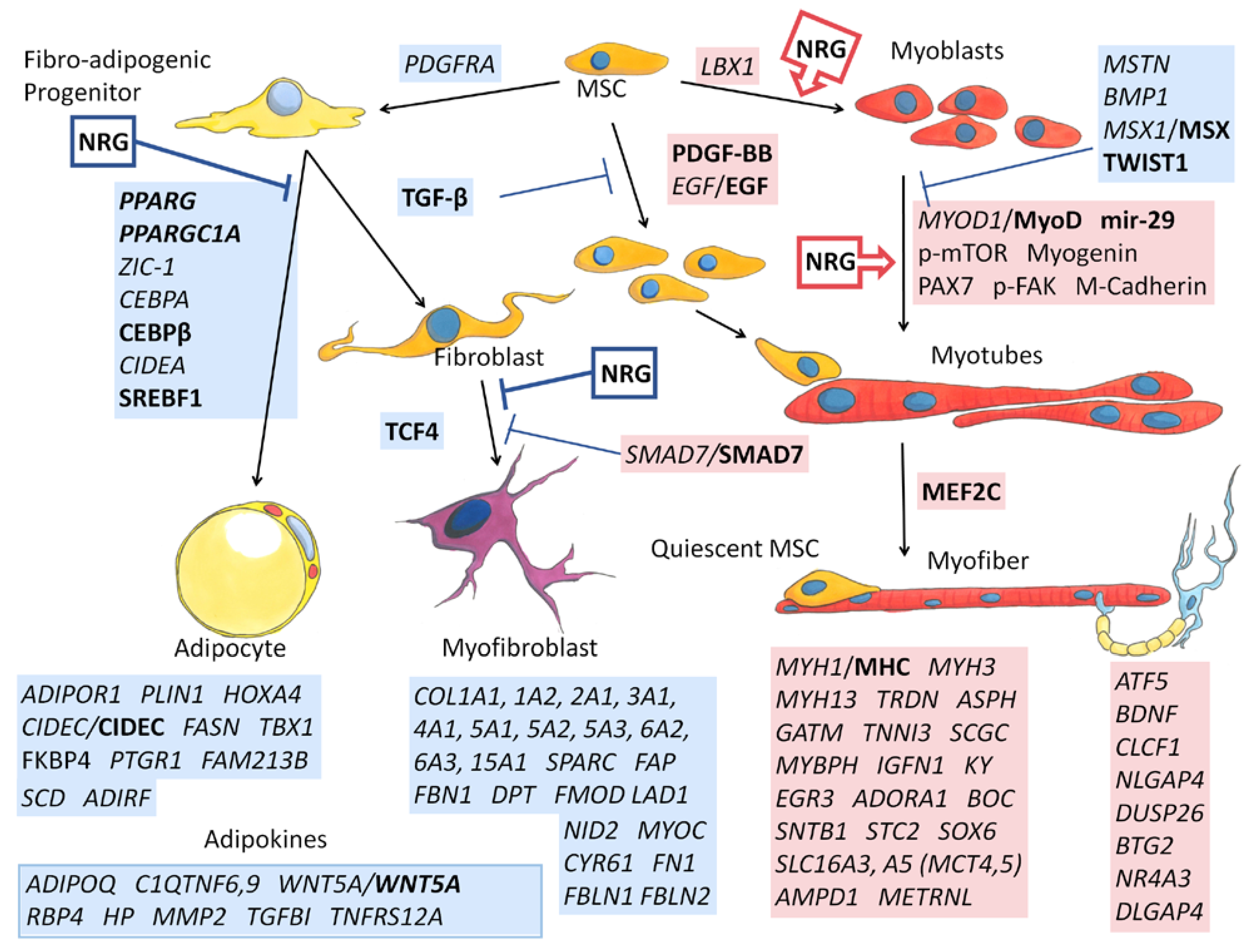{kind=link}
| Gene | Name | SM | LV | Function |
|---|---|---|---|---|
| NR4A3 | Nuclear Receptor Subfamily 4 Group A Member 3 | 10.4 | 12.8 | β-adrenergic inducible, Regulates transcription of fatty acid and muscle mass genes [20,21] |
| FOS | Fos Proto-Oncogene, AP-1 Transcription Factor Subunit | 4.9 | 5.0 | Activate phospholipid synthesis, Regulators of cell proliferation, muscle cell differentiation, and Transformation [22,23,24] |
| RND | Rho Family GTPase | 3.6 | 2.6 | Regulate the organization of the actin cytoskeleton in response to extracellular growth factors [54] |
| BTG2 | BTG Anti-Proliferation Factor 2 | 3.3 | 4.1 | Cell cycle regulation, cell proliferation in skeletal muscle [25] |
| PPP1R15A | Protein Phosphatase 1 Regulatory Subunit 15A | 2.8 | 1.7 | Down-regulates the TGF-beta, growth arrest and DNA damage-inducible protein, promoting cell death [55] |
| IER5 | Immediate Early Response 5 | 2.6 | 4.3 | Cell regulation, proliferation, and resistance to thermal stress. Dephosphorylates HSF1, and ribosomal protein S6 [56] |
| PAAF1 | Proteasomal ATPase Associated Factor 1 | 2.4 | 1.5 | Associated with Heart Conduction Disease, regulation of association of proteasome components [26,57] |
| ATP1B4 | ATPase Na+/K+ Transporting Family Member Beta 4 | 2.0 | 1.5 | Transporting protein, Transcriptional coregulator during muscle development [27,28] |
| COL1A2 | Collagen Type 1 Alpha 2 Chain | −1.8 | −2.7 | Fibrillar forming collagen, putative down-regulated c-Myc target gene, or upregulate let-7b in skin fibroblasts [58]. |
| EFEMP1 | EGF Containing Fibulin Extracellular Matrix Protein 1 | −1.8 | −1.7 | Binds EGFR receptor, autophosphorylation and the activation of downstream signaling pathways, Negative regulator of chondrocyte differentiation [59]. |
| LAMB1 | Laminin Subunit Beta 1 | −1.8 | −1.5 | Cell adhesion, Differentiation, Encoding laminin subunit beta-1, are associated with COB with variable muscular or ocular abnormalities, Expressed in skeletal muscle [29,30] |
| LUM | Lumican | −2.0 | −1.9 | A collagen binding proteoglycan with increased expression in hearts, Regulate tissue repair, collagen fibril organization [31,32] |
| THY1 | Thy-1 Cell Surface Antigen | −2.0 | −2.6 | Cell surface glycoprotein and A pathogenic CF fraction in cardiac fibrosis [33,34] |
| ECM1 | Extracellular Matrix Protein 1 | −2.1 | −2.2 | Response to elevated platelet cytosolic Ca2+ and ERK Signaling, Upregulated in cardiac aging and myocardial infarction [35,36] |
| FAP | Fibroblast Activation Protein Alpha | −2.2 | −1.8 | Expressed in stromal fibroblasts of epithelial cancers, tissue remodeling, healing wounds, Correlate coronary heart disease [37,38,39] |
| DPT | Dermatopontin | −2.3 | −2.1 | Extracellular matrix proteins involved in cell-matrix interaction, Postulated to modify the behavior of TGF-beta [40] |
| FBN1 | Fibrillin 1 | −2.3 | −2.6 | Calcium ion binding and extracellular matrix structural constituent, Differential regulation in smooth muscle cells [41,42,43] |
| PRPS2 | Phosphoribosyl Pyrophosphate Synthetase 2 | −2.3 | −1.7 | Phosphoribosyl pyrophosphate synthetase, protein homodimerization activity, and carbohydrate catabolic process-related genes [60,61] |
| NID1 | Nidogen 1 | −2.4 | −1.8 | Basement membrane glycoproteins, Laminin interactions in the heart [44,45,46] |
| PLSCR4 | Phospholipid Scramblase 4 | −2.5 | −2.3 | Protein coding gene; upregulated in hypertrophic mouse hearts [47,62] |
| WNT5A | Wnt Family Member 5A | −2.5 | −1.5 | Canonical and non-canonical wnt pathways, Regulating developmental pathways during embryogenesis |
| FSCN1 | Fascin Actin-Bundling Protein 1 | −2.5 | −1.5 | Organize F-actin; Involved in cell migration, motility, adhesion, and cellular interactions |
| COL4A1 | Collagen Type IV Alpha 1 Chain | −2.6 | −1.6 | Alpha protein of Type IV collagen, components of basement membranes, Upregulated in the skeletal muscle response [48,49] |
| ACER3 | Alkaline Ceramidase 3 | −2.6 | −1.5 | Protein coding gene |
| COL5A2 | Nidogen 1 | −2.6 | −2.5 | Alpha chain for fibrillar collagen; cardiac repair and involved in Muscle-Invasive Bladder [48,50,51] |
| PRTFDC1 | Phosphoribosyl Transferase Domain Containing 1 | −2.7 | −2.3 | Protein Coding gene, protein homodimerization activity, and magnesium ion binding |
| CD55 | CD55 molecule (Cromer Blood Group) | Reduced | −2.2 | Glycoprotein; Regulates cell decay dysferlin is expressed in skeletal and cardiac muscles [52] |
| COL3A1 | Collagen Type III Alpha 1 Chain | −2.8 | −1.9 | Fibrillar collagen found in extensible connective tissues and the vascular system [53] |
| HBB | Hemoglobin Subunit Beta | −3.0 | −5.9 | Oxygen transport from the lung, Endogenous inhibitor of enkephalin-degrading enzymes such as DPP3, and as a selective antagonist of the P2RX3 receptor which is involved in pain signaling [63] |
| HBA | Hemoglobin Subunit Alpha | −3.6 | −5.9 | Iron ion binding and oxygen transport from the lung to the various peripheral tissue [64] |
| ARMCX2 | Armadillo Repeat Containing X-Linked 2 | −3.7 | −1.8 | Regulate the dynamics and distribution of mitochondria in neural cells; involved in tissue development and tumorigenesis [65] |
| GEO Study ID | Description | Tissue | Species |
|---|---|---|---|
| GSE1557 [66] | Cardiac cachexia (n = 4) | Left ventricle | Rat |
| GSE52676 [67] | Starvation (n = 6) Denervation (n = 9) Type 1 Diabetes (n = 3) | Soleus | Mouse |
| GSE45331 [68] | Myotonic dystrophy type 2 (n = 6) Control (n = 4) | Vastus lat. | Human |
| GSE48574 [69] | ISCU (n = 3) vs. Control (n = 5) | Vastus lat. | Human |
| GSE38680 [70] | Pompe (n = 9) vs. Control (n = 10) Pompe (n = 11) vs. Control (n = 7) | Biceps Quad | Human |
| GSE34111 [71] | Cancer cachexia (n = 12) vs. Control (n = 6) | Quad | Human |
| GSE42806 [72] | Tibial muscular dystrophy (n = 7) vs. Healthy (n = 5) | Extensor digitorum longus | Human |
| GSE25941 [73] | Female: old (78 ± 1 years, n = 11) vs. young (25 ± 1 years, n = 8) Male: old (78 ± 1 years, n = 10) vs. young (25 ± 1 years, n = 7) | Vastus lat. | Human |
| GSE9103 [74] | Old (n = 65.1 ±1.5, n = 10) vs. young (22.7 ± 0.7, n = 10) | Vastus lat. | Human |
| GSE5110 [75] | 48 h immobilization vs. control: male subjects (n = 5) | Vastus lat. | Human |
| GSE21496 [76] | 48 h suspension vs. control: sedentary male subjects (n = 7) | Vastus lat. | Human |
| GSE43760 [77] | Metabolic syndrome (n = 6) vs. healthy (n = 6) | Vastus lat. | Human |
| GSE27536 | COPD low BMI (n = 6) vs. healthy (n = 12) COPD normal BMI (n = 8) vs. healthy (n = 12) | Vastus lat. | Human |
| GSE6798 [78] | Obese + PCOS (n = 16) vs. Control (n = 13) | Vastus lat. | Human |
| GSE8157 [79] | Obese + PCOS (n = 10) vs. Control (n = 13) | Vastus lat. | Human |
| GSE19420 | Type 2 diabetes (n = 10) vs. normoglycemic subjects (n = 12) | Vastus lat. | Human |
| GSE25462 [80] | Type 2 diabetes (n = 10) vs. normoglycemic + no family history of diabetes (n = 15) Type 2 diabetes (n = 10) vs. normoglycemic + family history of type 2 diabetes (n = 25) | Quad | Human |
| Upstream Regulator | Gene Expression | No. Downstream Targets Altered (p-Value) | Z Score (Predicted State) |
|---|---|---|---|
| Transforming growth factor β1 (TGF-β1) | - | 114 (1.6 × 10−23) | Z = −2.297 (Inhibited) |
| Platelet derived growth factor-BB (PDGF BB) | - | 36 (3.7 × 10−22) | Z = 1.567 |
| Angiotensinogen (AGT) | −3.8 | 67 (8.5 × 10−20) | Z = −3.032 (Activated) |
| Fibroblast growth factor 2 (FGF2) | - | 45 (4.8 × 10−20) | nd |
| CAMP Responsive Element Binding Protein 1 (CREB1) | - | 46 (3.5 × 10−19) | Z = 2.424 (Activated) |
| Erb-B2 Receptor Tyrosine Kinase 2 (ERBB2) | - | 60 (5.7 × 10−19) | nd |
| PPARG coactivator 1 α (PCG-1α) | - | 37 (4.4 × 10−18) | Z = 1.877 |
| Aryl hydrocarbon receptor (AHR) | - | 45 (4.5 × 10−17) | Z = 3.63 (Activated) |
| Twist family BHLH transcription factor 1 (TWIST1) | - | 23 (6.7 × 10−17) | Z = −2.449 (Inhibited) |
| Epidermal growth factor (EGF) | 3.5 | 49 (5.3 × 10−16) | nd |
| Cadherin associated protein α1 (α-catenin) | - | 19 (3.0 × 10−15) | Z = 3.118 (Activated) |
| Erb-B3 Receptor Tyrosine Kinase 3 (ERRB3) | - | 18 (2.5 × 10−13) | nd |
| Mothers against DPP homolog 7 (SMAD7) | 2.4 | 18 (3.3 × 10−12) | Z = 3.11 (Activated) |
| CCAAT enhancer binding protein β (C/EBP-β) | - | 37 (1.1 × 10−11) | Z = −2.091 (Inhibited) |
| AKT serine/threonine kinase 1 (AKT1) | - | 26 (1.9 × 10−11) | Z = −1.855 |
| Enalapril (Hypertension medication) | - | 13 (9.3 × 10−11) | Z = 3.073 (Activated) |
| CCAAT enhancer binding protein α (C/EBP-α) | −9.3 | 28 (2.5 × 10−11) | Z = −1.812 |
| Brain-derived neurotrophic factor (BDNF) | 30 (6.7 × 10−11) | nd | |
| Transforming growth factor β3 (TGF-β3) | - | 17 (7.6 × 10−11) | Z = −1.937 |
| Myocilin (MYOC) | −2.5 | 13 (1.7 × 10−10) | nd |
| Myocyte enhancer factor 2C (MEF2C) | - | 15 (3.7 × 10−10) | Z = 2.912 (Activated) |
| Peroxisome proliferator activated receptor γ (PPAR-γ) | −5.4 | 36 (8.6 × 10−10) | Z = −1.987 |
| Sterol regulatory element-binding transcription factor 1 (SREBF1) | - | 20 (2.4 × 10−9) | Z = −1.937 |
| Erb-B3 Receptor Tyrosine Kinase 3 (ERRB4) | - | 12 (3.4 × 10−8) | Z = 2.388 (Activated) |
| Transforming growth factor β1 (TGF-β1) | - | 15 (4.3 × 10−8) | Z = −1.634 |
| microRNA-29b-3p (miR-29b-3p) | - | 13 (4.8 × 10−8) | Z = 3.097 (Activated) |
| Thapsigargin (Calcium reuptake inhibitor) | - | 20 (6.5 × 10−8) | Z = 3.450 (Activated) |
| microRNA-335-3p (miR-335-3p) | - | 8 (6.7 × 10−8) | Z = 2.828 (Activated) |
| CCAAT enhancer binding protein δ (C/EBP-δ) | - | 13 (1.3 × 10−7) | Z = −1.813 |
| Peroxisome proliferator activated receptor α (PPAR-α) | - | 30 (2.5 × 10−7) | Z = −1.363 |
| Clopidogrel (antiplatelet blood-thinning medication) | 11 (4.6 × 10−7) | Z = 3.302 (Activated) | |
| microRNA lethal 7a-5p (Let-7a-5p) | - | 17 (1.4 × 10−6) | Z = 3.682 (Activated) |
| Myocardin (MYOCD) | - | 9 (4.4 × 10−6) | Z = 2.759 (Activated) |
| Cell death inducing DFFA like effector C (CIDEC) | −9.4 | 6 (1.1 × 10−5) | nd |
| Msh homeobox 1 (MSX1) | −1.7 | 5 (1.8 × 10−5) | Z = −1.982 |
| microRNA-30c-5p (miR-30c-5p) | - | 17 (2.1 × 10−5) | Z = 3.117 (Activated) |
| sterol regulatory element binding transcription factor 2 (SREBF2) | - | 9 (5.8 × 10−5) | Z = −2.394 (Inhibited) |
| microRNA-21 (miR-21) | - | 17 (3.3 × 10−5) | Z = −2.668 (Inhibited) |
| Myogenic differentiation 1 (MYOD1) | 2.2 | 16 (3.7 × 10−5) | nd |
| Wnt family member 5a (WNT5a) | −2.5 | 10 (8.8 × 10−5) | Z = −2.394 (Inhibited) |
| Peroxisome proliferator activated receptor δ (PPAR-δ) | - | 15 (8.5 × 10−4) | Z = −1.214 |
| 26s Proteosome (protein complex) | - | 11 (5.5 × 10−4) | Z = −2.035 (Inhibited) |
| Transcription factor 4 (TCF4) | - | 18 (8.7 × 10−4) | Z = −1.554 |
| PPARG coactivator 1 β (PCG-1β) | - | 5 (1.6 × 10−2) | Z = −2.186 (Inhibited) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galindo, C.L.; Nguyen, V.T.; Hill, B.; Easterday, E.; Cleator, J.H.; Sawyer, D.B. Neuregulin (NRG-1β) Is Pro-Myogenic and Anti-Cachectic in Respiratory Muscles of Post-Myocardial Infarcted Swine. Biology 2022, 11, 682. https://doi.org/10.3390/biology11050682
Galindo CL, Nguyen VT, Hill B, Easterday E, Cleator JH, Sawyer DB. Neuregulin (NRG-1β) Is Pro-Myogenic and Anti-Cachectic in Respiratory Muscles of Post-Myocardial Infarcted Swine. Biology. 2022; 11(5):682. https://doi.org/10.3390/biology11050682
Chicago/Turabian StyleGalindo, Cristi L., Van Thuan Nguyen, Braxton Hill, Ethan Easterday, John H. Cleator, and Douglas B. Sawyer. 2022. "Neuregulin (NRG-1β) Is Pro-Myogenic and Anti-Cachectic in Respiratory Muscles of Post-Myocardial Infarcted Swine" Biology 11, no. 5: 682. https://doi.org/10.3390/biology11050682