
Morphological and Molecular Description of Sarcocystis myodes n. sp. from the Bank Vole (Clethrionomys glareolus) in Lithuania


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Samples
2.2. Morphological Examination
2.3. DNA Extraction and PCR
2.4. Sequence Analysis
3. Results
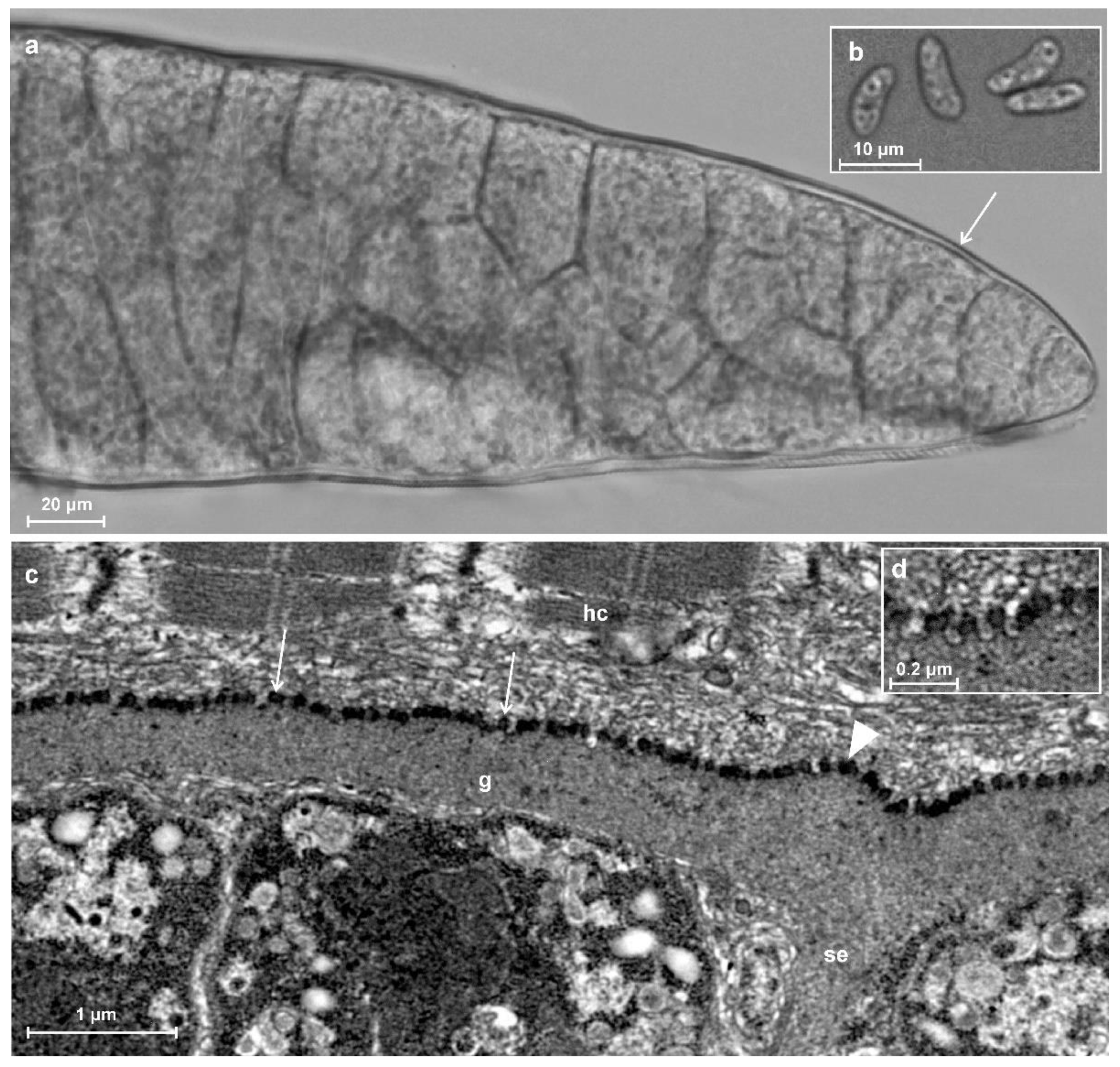
3.1. Prevalence and Morphological Description of Sarcocysts
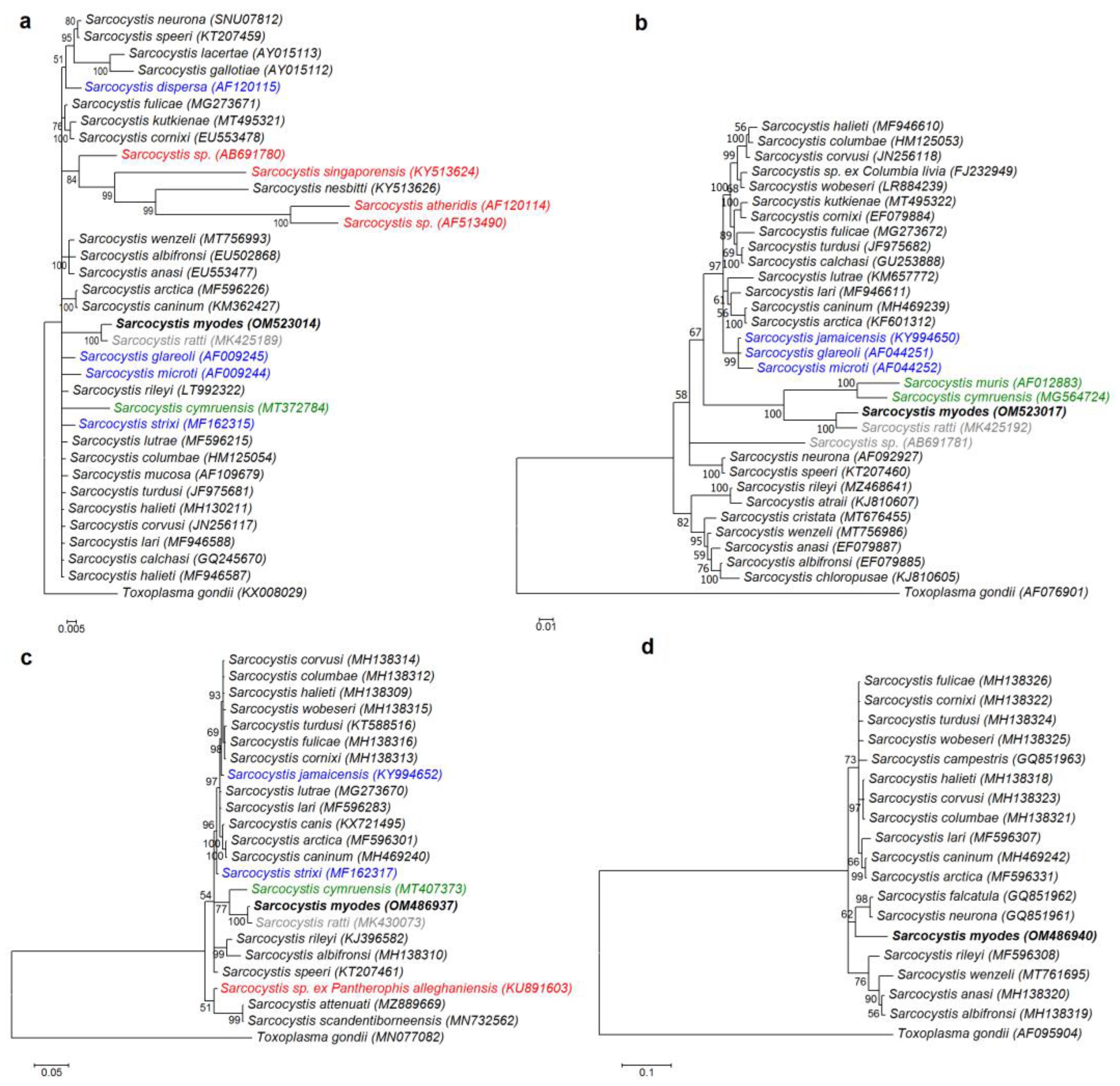
3.2. Molecular Characterization and Phylogeny
3.3. Description of Sarcocystis myodes n. sp.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Wilson, D.E.; Lacher, T.E.; Mittermeier, R.A. Handbook of the Mammals of the World; Lynx Edicions: Barselona, Spain, 2017; Volume 7. [Google Scholar]
- Malecha, A.W.; Antczak, M. Diet of the European polecat Mustela putorius in an agricultural area in Poland. Folia. Zool. 2013, 62, 48–53. [Google Scholar] [CrossRef]
- Grabham, A.A.; Ventress, G.; Hayward, M.W. The diet of denning female European pine martens (Martes martes) in Galloway Forest District, South West Scotland, Great Britain. Mammal. Res. 2019, 64, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Gryz, J.; Krauze-Gryz, D. Changes in the tawny owl Strix aluco diet along an urbanisation gradient. Biologia 2019, 74, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 7039–7044. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Pastor, R.; Escudero, R.; Lambin, X.; Vidal, M.D.; Gil, H.; Jado, I.; Rodríguez-Vargas, M.; Luque-Larena, J.J.; Mougeot, F. Zoonotic pathogens in fluctuating common vole (Microtus arvalis) populations: Occurrence and dynamics. Parasitology 2018, 146, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Votýpka, J.; Hypsa, V.; Jirků, M.; Flegr, J.; Vávra, J.; Lukes, J. Molecular phylogenetic relatedness of Frenkelia spp. (Protozoa, Apicomplexa) to Sarcocystis falcatula Stiles 1893: Is the genus Sarcocystis paraphyletic? J. Eukaryot. Microbiol. 1998, 45, 137–141. [Google Scholar] [CrossRef]
- Dolezel, D.; Koudela, B.; Jirků, M.; Hypsa, V.; Oborník, M.; Votýpka, J.; Modrý, D.; Šlapeta, J.R.; Lukes, J. Phylogenetic analysis of Sarcocystis spp. of mammals and reptiles supports the coevolution of Sarcocystis spp. with their final hosts. Int. J. Parasitol. 1999, 29, 795–798. [Google Scholar] [CrossRef]
- Mugridge, N.B.; Morrison, D.A.; Johnson, A.M.; Luton, K.; Dubey, J.P.; Votýpka, J.; Tenter, A.M. Phylogenetic relationships of the genus Frenkelia: A review of its history and new knowledge gained from comparison of large subunit ribosomal ribonucleic acid gene sequences. Int. J. Parasitol. 1999, 29, 957–972. [Google Scholar] [CrossRef]
- Mugridge, N.B.; Morrison, D.A.; Jäkel, T.; Heckeroth, A.R.; Tenter, A.M.; Johnson, A.M. Effects of sequence alignment and structural domains of ribosomal DNA on phylogeny reconstruction for the protozoan family sarcocystidae. Mol. Biol. Evol. 2000, 17, 1842–1853. [Google Scholar] [CrossRef]
- Šlapeta, J.R.; Kyselová, I.; Richardson, A.O.; Modrý, D.; Lukeš, J. Phylogeny and sequence variability of the Sarcocystis singaporensis Zaman and Colley, (1975) 1976 ssrDNA. Parasitol. Res. 2002, 88, 810–815. [Google Scholar] [CrossRef]
- Hu, J.J.; Meng, Y.; Guo, Y.M.; Liao, J.Y.; Song, J.L. Completion of the life cycle of Sarcocystis zuoi, a parasite from the Norway rat, Rattus norvegicus. J. Parasitol. 2012, 98, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Antunes Murata, F.H.; Cerqueira-Cézar, C.K.; Thompson, P.C.; Tiwari, K.; Mowery, J.D.; Verma, S.K.; Rosenthal, B.M.; Sharma, R.N.; Dubey, J.P. Sarcocystis cymruensis: Discovery in Western Hemisphere in the Brown rat (Rattus norvegicus) from Grenada, West Indies: Redescription, molecular characterization, and transmission to IFN-γ gene knockout mice via sporocysts from experimentally infected domestic cat (Felis catus). Parasitol. Res. 2018, 117, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Kirillova, V.; Gavarāne, I.; Grāvele, E.; Butkauskas, D.; Rudaitytė-Lukošienė, E.; Kirjušina, M. Morphological and molecular description of Sarcocystis ratti n. sp. from the black rat (Rattus rattus) in Latvia. Parasitol. Res. 2019, 118, 2689–2694. [Google Scholar] [CrossRef] [PubMed]
- Prūsaitė, J. Lithuanian Fauna. Mammals; Mokslas: Vilnius, Lietuva, 1988. [Google Scholar]
- Balčiauskas, L.; Trakimas, G.; Juškaitis, R.; Ulevičius, A.; Balčiauskienė, L. Atlas of Lithuanian Mammals, Amphibians and Reptiles, 2nd ed.; Akstis: Vilnius, Lithuania, 1999. [Google Scholar]
- Balčiauskas, L.; Čepukienė, A.; Balčiauskienė, L. Small mammal community response to early meadow–forest succession. For. Ecosyst. 2017, 4, 11. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L.; Garbaras, A.; Stirkė, V. Diversity and diet differences of small mammals in commensal habitats. Diversity 2021, 13, 346. [Google Scholar] [CrossRef]
- Häfner, U.; Matuschka, F.R. Life cycle studies on Sarcocystis dirumpens sp. n. with regard to host specificity. Z. Parasitenkd. 1984, 70, 715–720. [Google Scholar] [CrossRef]
- Matuschka, F.R. Sarcocystis clethrionomyelaphis n. sp. from snakes of the genus Elaphe and different voles of the family Arvicolidae. J. Parasitol. 1986, 72, 226–231. [Google Scholar] [CrossRef]
- Matuschka, F.R.; Heydorn, A.O.; Mehlhorn, H.; Abd-Al-Aal, Z.; Diesing, L.; Biehler, A. Experimental transmission of Sarcocystis muriviperae n. sp. to laboratory mice by sporocysts from the Palestinian viper (Vipera palestinae): A light and electron microscope study. Parasitol. Res. 1987, 73, 33–40. [Google Scholar] [CrossRef]
- Upton, S.J.; McKown, R.D. The red-tailed hawk, Buteo jamaicensis, a native definitive host of Frenkelia microti (Apicomplexa) in North America. J. Wildl. Dis. 1992, 28, 85–90. [Google Scholar] [CrossRef]
- Svobodová, M.; Vořišek, P.; Votýpka, J.; Weidinger, K. Heteroxenous coccidia (Apicomplexa: Sarcocystidae) in the populations of their final and intermediate hosts: European buzzard and small mammals. Acta Protozool. 2004, 43, 251–260. [Google Scholar]
- Kutkienė, L.; Prakas, P.; Sruoga, A.; Butkauskas, D. The mallard duck (Anas platyrhynchos) as intermediate host for Sarcocystis wobeseri sp. nov. from the barnacle goose (Branta leucopsis). Parasitol. Res. 2010, 107, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Wendte, J.M.; Miller, M.A.; Nandra, A.K.; Peat, S.M.; Crosbie, P.R.; Conrad, P.A.; Grigg, M.E. Limited genetic diversity among Sarcocystis neurona strains infecting southern sea otters precludes distinction between marine and terrestrial isolates. Vet. Parasitol. 2010, 169, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gjerde, B. Phylogenetic relationships among Sarcocystis species in cervids, cattle and sheep inferred from the mitochondrial cytochrome c oxidase subunit I gene. Int. J. Parasitol. 2013, 43, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Butkauskas, D.; Švažas, S.; Stanevičius, V. Morphological and genetic characterisation of Sarcocystis halieti from the great cormorant (Phalacrocorax carbo). Parasitol. Res. 2018, 117, 3663–3667. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Milne, I.; Wright, F.; Rowe, G.; Marshall, D.; Husmeier, D.; McGuire, G. TOPALi: Software for automatic identification of recombinant sequences within DNA multiple alignments. Bioinformatics 2004, 20, 1806–1807. [Google Scholar] [CrossRef]
- Ruiz, A.; Frenkel, J.K. Recognition of cyclic transmission of Sarcocystis muris by cats. J. Infect. Dis. 1976, 133, 409–418. [Google Scholar] [CrossRef]
- Rommel, M. The ferret (Putorius putorius furo), an additional final host of Sarcocystis muris. Z. Parasitenkd. 1979, 58, 187–188. (In German) [Google Scholar] [CrossRef]
- Krampitz, H.E.; Rommel, M.; Geisel, O.; Kaiser, E. Beiträge zum Lebenszyklus der Frenkelien II. Die ungeschlechtliche Entwicklung von Frenkelia clethrionomyobuteonis in der Rötelmaus. Z. Parasitenkd. 1976, 51, 7–14. [Google Scholar] [CrossRef]
- Verma, S.K.; von Dohlen, A.R.; Mowery, J.D.; Scott, D.; Rosenthal, B.M.; Dubey, J.P.; Lindsay, D.S. Sarcocystis jamaicensis n. sp., from Red-Tailed Hawks (Buteo jamaicensis) definitive host and IFN-γ gene knockout mice as experimental intermediate host. J. Parasitol. 2017, 103, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Černá, Z.; Kolárová, I.; Sulc, P. Contribution to the problem of cyst-producing coccidians. Folia Parasitol. 1978, 25, 9–16. [Google Scholar]
- Paperna, I.; Martelli, P. Fine structural development of microgamonts of Sarcocystis singaporensis in Python reticulatus. Parasitol. Res. 2000, 86, 1022–1025. [Google Scholar] [CrossRef] [PubMed]
- Šlapeta, J.R.; Modrý, D.; Koudela, B. Sarcocystis atheridis sp. nov., a new sarcosporidian coccidium from Nitsche’s bush viper, Atheris nitschei Tornier, 1902, from Uganda. Parasitol. Res. 1999, 85, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Arnastauskienė, T.; Grikienienė, J. Infection of small mammals with sarcosporidians in the South-Eastern Baltic region. Ecology 1993, 2, 47–56. (In Russian) [Google Scholar]
- Grikienienė, J.; Mažeikytė, R. Investigation of sarcosporidians (Sarcocystis) of small mammals in Kamasta landscape reserve and its surroundings. Acta Zool. Litu. 2000, 10, 55–68. [Google Scholar] [CrossRef]
- Grikienienė, J.; Malakauskas, M.; Mažeikytė, R.; Balčiauskas, L.; Senutaitė, J. Muscle parasites (Sarcocystis, Trichinella, Alaria) of wild mammals in Lithuania. Theriol. Litu. 2001, 1, 29–46. [Google Scholar]
- Grikienienė, J. Investigations into endoparasites of small mammals in the environs of Lake Drūkšiai. Acta Zool. Litu. 2005, 15, 109–114. [Google Scholar] [CrossRef]
- Fernández-Escobar, M.; Millán, J.; Chirife, A.D.; Ortega-Mora, L.M.; Calero-Bernal, R. Molecular survey for cyst-forming coccidia (Toxoplasma gondii, Neospora caninum, Sarcocystis spp.) in Mediterranean periurban micromammals. Parasitol. Res. 2020, 119, 2679–2686. [Google Scholar] [CrossRef]
- Hu, J.J.; Liao, J.Y.; Meng, Y.; Guo, Y.M.; Chen, X.W.; Zuo, Y.X. Identification of Sarcocystis cymruensis in wild Rattus flavipectus and Rattus norvegicus from Peoples Republic of China and its transmission to rats and cats. J. Parasitol. 2011, 97, 421–424. [Google Scholar] [CrossRef]
- Hu, J.J.; Liu, Q.; Yang, Y.F.; Esch, G.W.; Guo, Y.M.; Zou, F.C. Sarcocystis eothenomysi n. sp. (Apicomplexa: Sarcocystidae) from the large oriental vole Eothenomys miletus (Thomas) (Cricetidae: Microtinae) from Anning, China. Syst. Parasitol. 2014, 89, 73–81. [Google Scholar] [CrossRef]
- Jäkel, T.; Khoprasert, Y.; Sorger, I.; Kliemt, D.; Seehabutr, V.; Suasa-ard, K.; Hongnark, S. Sarcosporidiasis in rodents from Thailand. J. Wildl. Dis. 1997, 33, 860–867. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, P.J.; Watts, C.H.; Dixon, B.R. Ultrastructure of Sarcocystis spp. (Protozoa: Apicomplexa) in rodents from North Sulawesi and West Java, Indonesia. J. Wildl. Dis. 1987, 23, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.; Guo, Y.; Ma, C.; Deng, S.; Hu, J.; Zhang, Y. Redescription and molecular characterization of sarcocysts of Sarcocystis cymruensis from Norway rats (Rattus norvegicus) and Sarcocystis ratti from black rats (R. rattus) in China. Parasitol. Res. 2020, 119, 3785–3791. [Google Scholar] [CrossRef] [PubMed]
- Jasiulionis, M.; Balčiauskas, L. Seasonal and daily activity patterns of mammals in the colony of great cormorants. Mammalia 2021, 85, 439–447. [Google Scholar] [CrossRef]
- Häfner, U.; Frank, W. Morphological studies on the muscle cysts of Sarcocystis dirumpens (Hoare 1933) Häfner and Matuschka 1984 in several host species revealing endopolygeny in metrocytes. Z. Parasitenkd. 1986, 72, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Černá, Z.; Loučková, M. Microtus arvalis as the intermediate host of a coccidian from the kestrel (Falco tinnunculus). Folia Parasitol. 1976, 23, 110. [Google Scholar]
- Dubey, J.P. Sarcocystis montanaensis and S. microti sp. n. from the meadow vole (Microtus pennsylvanicus). Proc. Helminthol. Soc. Wash. 1983, 50, 318–324. [Google Scholar]
- Lindsay, D.S.; Upton, S.J.; Blagburn, B.L.; Toivio-Kinnucan, M.; McAllister, C.T.; Trauth, S.E. Sporocysts isolated from the southern copperhead (Agkistrodon contortrix contortrix) produce Sarcocystis montanaensis-like sarcocysts in prairie voles (Microtus ochrogaster). J. Wildl. Dis. 1991, 27, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, D.S.; Upton, S.J.; Biagburn, B.L.; Toivio-Kinnucan, M.; Dubey, J.P.; McAllister, C.T.; Trauths, S.E. Demonstration that Sarcocystis montanaensis has a Speckled kingsnake-Prairie vole life cycle. J. Helminthol. Soc. Wash. 1992, 59, 9–15. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sequence Data | 18S rDNA | 28S rDNA | cox1 | rpoB | ITS1 |
|---|---|---|---|---|---|
| Sequence length (bp) | 1786 | 1545 | 1053 | 762 | 969 |
| Genbank acc. no. | OM523014–OM523016 | OM523017–OM523019 | OM486937–OM486939 | OM486940–OM486942 | OM523020–OM523022 |
| Diversity amongst three isolates (%) | 0 | 0 | 0 | 0 | 0–0.5 |
| Highest sequences similarity values with other Sarcocystis spp. | S. ratti from black rat 99.5–99.6% (MT372787, MK425189); S. rileyi from mallard duck 98.1% (GU120092); S. lutrae from mustelids 98.0% (MG372102-03) | S. ratti from black rat 98.1% (MK425192); S. muris from house mouse 93.5% (AF012883); S. cymruensis from brown rat 93.5% (MG564724) | S. ratti from black rat 99.4% (MK430072, MT407374); S. strixi from laboratory mouse 95.9% (MF162317) | S. fulicae from Eurasian coot 95.7% (MH138326); S. cornixi from hooded crow 95.7% (MH138322), S. caninum from dog 95.7% (MH469242); S. arctica from red fox 95.4–95.7% (MF596311-31) | S. ratti from black rat 77.0–77.3% (MK910965) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudaitytė-Lukošienė, E.; Jasiulionis, M.; Balčiauskas, L.; Prakas, P.; Stirkė, V.; Butkauskas, D. Morphological and Molecular Description of Sarcocystis myodes n. sp. from the Bank Vole (Clethrionomys glareolus) in Lithuania. Biology 2022, 11, 512. https://doi.org/10.3390/biology11040512
Rudaitytė-Lukošienė E, Jasiulionis M, Balčiauskas L, Prakas P, Stirkė V, Butkauskas D. Morphological and Molecular Description of Sarcocystis myodes n. sp. from the Bank Vole (Clethrionomys glareolus) in Lithuania. Biology. 2022; 11(4):512. https://doi.org/10.3390/biology11040512
Chicago/Turabian StyleRudaitytė-Lukošienė, Eglė, Marius Jasiulionis, Linas Balčiauskas, Petras Prakas, Vitalijus Stirkė, and Dalius Butkauskas. 2022. "Morphological and Molecular Description of Sarcocystis myodes n. sp. from the Bank Vole (Clethrionomys glareolus) in Lithuania" Biology 11, no. 4: 512. https://doi.org/10.3390/biology11040512