
Detection of Carbapenem Resistance of Proteus mirabilis Strains Isolated from Foxes, Raccoons and Minks in China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates and Growth Conditions
2.2. Antimicrobial Susceptibility Testing
2.3. Detection of Antimicrobial Resistance Genes
2.4. Detection of Virulence Genes
2.5. Infection of Galleria Mellonella Larvae
2.6. Determination of In Vivo Bacterial Loads
2.7. Phylogenetic Tree Analysis
2.8. Data Availability
3. Results
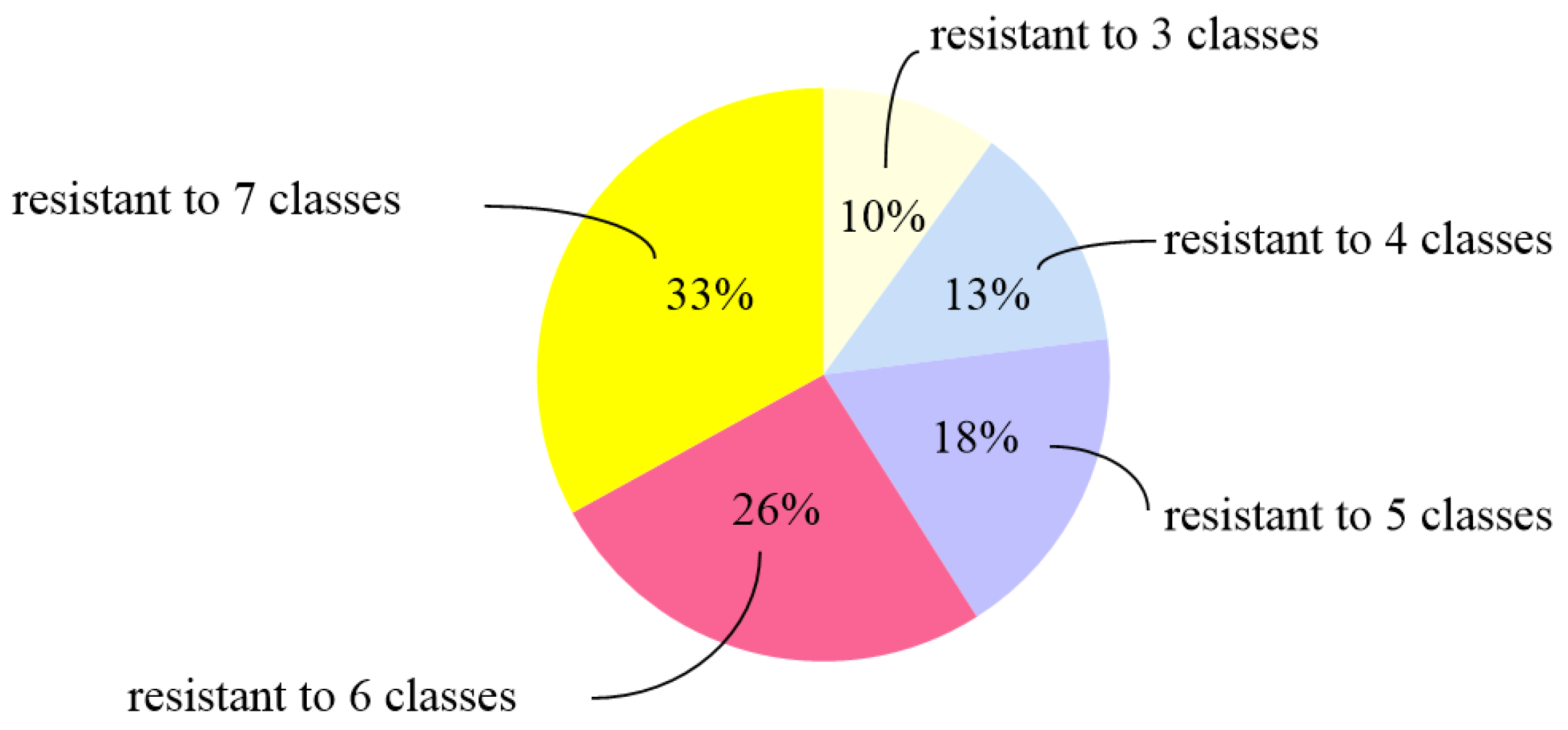
3.1. Antimicrobial Susceptibility of P. mirabilis Isolates
3.2. Antimicrobial Resistance Genes of P. mirabilis
3.3. Virulence Genes of P. mirabilis Isolates
3.4. Pathogenicity of P. mirabilis Isolates
3.4.1. Pathogenicity Comparison of P. mirabilis Isolates from Different Animal Sources
3.4.2. Pathogenicity of P. mirabilis Isolates with Different Virulence Genes
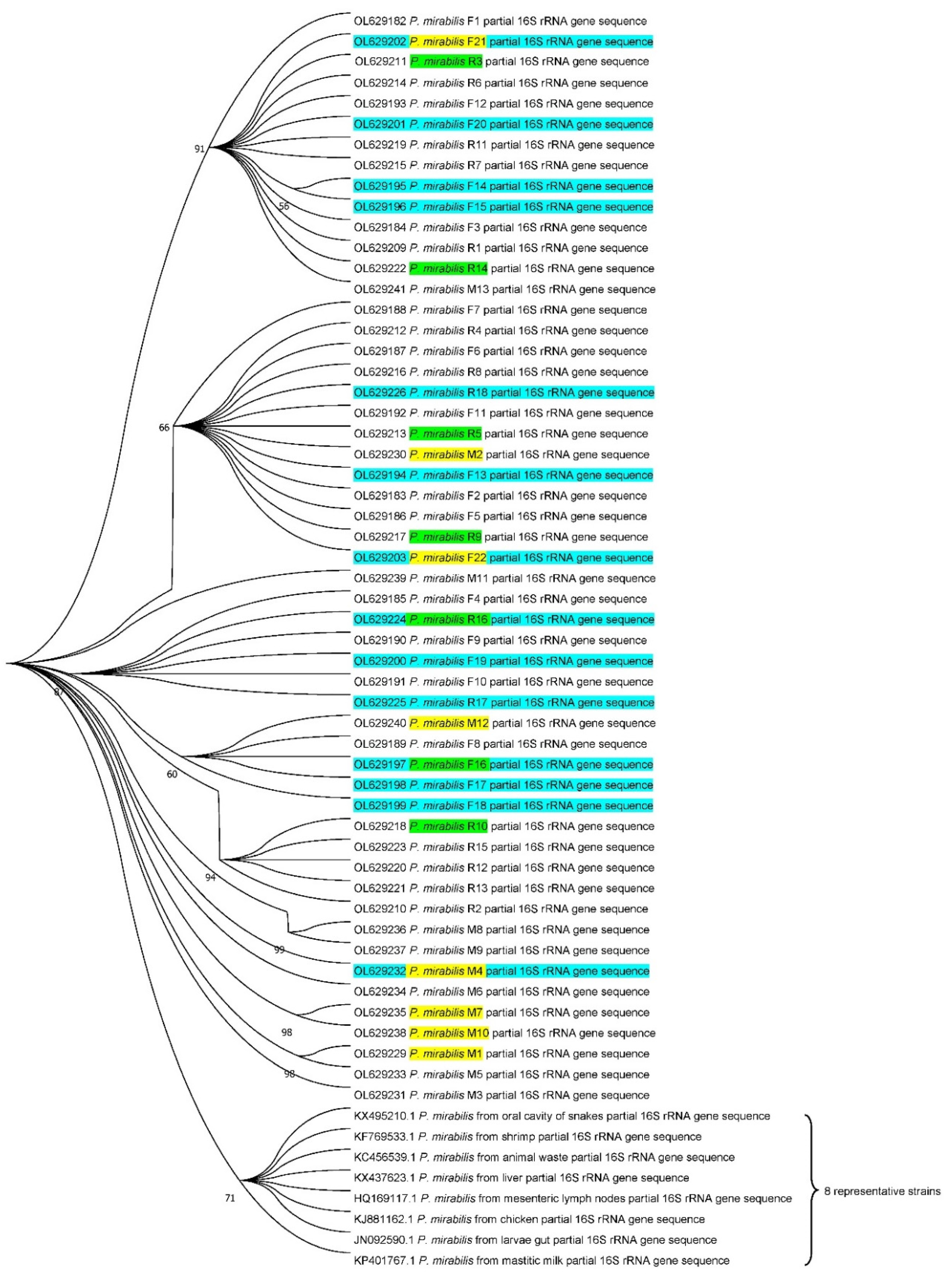
3.5. Phylogenetic Analysis of P. mirabilis Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene | Sequence (5’-3’) | Size (bp) | Reference |
|---|---|---|---|---|
| Beta-lactams | blaTEM | F: ATAAAATTCTTGAAGACGAAA | 643 | [27] |
| R: GACAGTTACCAATGCTTAATC | ||||
| blaSHV | F: TTATCTCCCTGTTAGCCACC | 860 | [27] | |
| R: GATTTGCTGATTTCGCTCGG | ||||
| blaPSE | F: TAGGTGTTTCCGTTCTTG | 150 | [28] | |
| R: TCATTTCGCTCTTCCATT | ||||
| blaOXA-1 | F: TCAACTTTCAAGATCGCA | 591 | [27] | |
| R: GTGTGTTTAGAATGGTGA | ||||
| blaOXA-23 | F: GATCGGATTGGAGAACCAGA | 501 | [30] | |
| R: ATTTCTGACCGCATTTCCAT | ||||
| blaOXA-24 | F: TTCCCCTAACATGAATTTGT | 1024 | [30] | |
| R: GTACTAATCAAAGTTGTGAA | ||||
| R: TGGATTGCACTTCATCTTGG | ||||
| blaOXA-58 | F: TGGCACGCATTTAGACCG | 507 | [30] | |
| R: AAACCCACATACCAACCC | ||||
| blaOXA-48 | F: GCGTGGTTAAGGATGAACAC | 438 | [29] | |
| R: CATCAAGTTCAACCCAACCG | ||||
| blaCTX-M | F: CGCTTTGCGATGTGCAG | 550 | [27] | |
| R: ACCGCGATATCGTTGGT | ||||
| blaKPC | F: GACGGAAAGCTTACAAAAACTGACA | 259 | [31] | |
| R: CTTGTCATCCTTGTTAGGCG | ||||
| blaNDM | F: GGTTTGGCGATCTGGTTTTC | 181 | [31] | |
| R: ATCCAGTTGAGGATCTGGGC | ||||
| blaIMP | F: GGAATAGAGTGGCTTAATTCTC | 275 | [31] | |
| R: CAAGCTTCTATATTTGCGTCACC | ||||
| blaVIM | F: GATGAGTTGCTTTTGATTGATACAGC | 153 | [31] | |
| R: CGCCCGAAGGACATCAA | ||||
| Quinolones | qnrA | F: ATTTCTCACGCCAGGATTTG | 519 | [27] |
| R: GATCGGCAAAGGTCAGGTCA | ||||
| qnrB | F: GATCGTGAAAGCCAGAAAGG | 513 | [27] | |
| R: ACGATGCCTGGTAGTTGTCC | ||||
| qnrC | F: GGTTGTACATTTATTGAATC | 666 | [27] | |
| R: TCCACTTTACGAGGTTCT | ||||
| qnrS | F: ACGACATTCGTCAACTGCAA | 417 | [27] | |
| R: TAAATTGGCACCCTGTAGGC | ||||
| oqxA | F: GATCAGTCAGTGGGATAGTTT | 670 | [35] | |
| R: TACTCGGCGTTAACTGATTA | ||||
| aac(6′)-Ib-cr | F: TTGCGATGCTCTATGAGTGGCTA | 482 | [27] | |
| R: CTCGAATGCCTGGCGTGTTT | ||||
| Aminoglycosides | aaC1 | F: ACCTACTCCCAACATCAGCC | 528 | [32] |
| R: ATATAGATCTCACTACGCGC | ||||
| aaC2 | F: ACTGTGATGGGATACGCGTC | 482 | [32] | |
| R: CTCCGTCAGCGTTTCAGCTA | ||||
| aaC3 | F: CACAAGAACGTGGTCCGCTA | 185 | [32] | |
| R: AACAGGTAAGCATCCGCATC | ||||
| aadA | F: GTGGATGGCGGCCTGAAGCC | 535 | [34] | |
| R: AATGCCCAGTCGGCAGCG | ||||
| aadB | F: ATGGACACAACGCAGGTC | 495 | [33] | |
| R: TTAGGCCGCATATCGCGACC | ||||
| aphA6 | F: ATGGAATTGCCCAATATTATTC | 399 | [33] | |
| R: TCAATTCAATTCATCAAGTTTTA | ||||
| Folate pathway antagonists | sul1 | F: CTTCGATGAGAGCCGGCGG C | 238 | [37] |
| R: GCAAGGCGGAAACCCGCGCC | ||||
| sul2 | F: CGGCATCGTCAACATAAC C | 722 | [36] | |
| R: GTGTGCGGATGAAGTCAG | ||||
| sul3 | F: AGATGTGATTGATTTGGGAGC | 443 | [38] | |
| R: TAGTTGTTTCTGGATTAGAGCCT | ||||
| Phenicols | cmlA | F: TGTCATTTACGGCATACTCG | 900 | [37] |
| R: ATCAGGCATCCCATTCCCAT | ||||
| floR | F: CACGTTGAGCCTCTATATGG | 890 | [27] | |
| R: ATGCAGAAGTAGAACGCGAC |
| Gene | Sequence (5′→3′) | Fragment | Annealing Temperature |
|---|---|---|---|
| ureC | F: GTTATTCGTGATGGGATGGG | 375 bp | 52 °C |
| R: ATAAAGGTGGTTACGCCAG | |||
| mrpA | F: ATTTCAGGAAACAAAAGATG | 410 bp | 39 °C |
| R: TTCTTACTGATAAGACATTG | |||
| zapA | F: ACCGCAGGAAAACATATAGCCC | 493 bp | 52 °C |
| R: GCGACTATCTTCCGCATAATCA | |||
| atfA | F: CATAATTTCTAGACCTGCCCTAGCA | 365 bp | 49 °C |
| R: CTGCTTGGATCCGTAATTTTTAACG | |||
| atfC | F: AGAAAGGGATCCTACAAATTAA | 472 bp | 49 °C |
| R: TATAGCATGCATTTAAATTGCC | |||
| ucaA | F: GTAAAGTTGTTGCGCAAAC | 365 bp | 49 °C |
| R: TTGAGCCACTGTGGATACA | |||
| pmfA | F: GGATCATCTATAATGAAACTG | 534 bp | 52 °C |
| R: CTGATAATCAACTTGGAAGTT | |||
| rsbA | F: TCGATTTCAGTGTTTGGCCAT | 1647 bp | 55 °C |
| R: TCGATTTCAGTGTTTGGCCAT | |||
| rsmA | F: TAGCGAGTGTTGACGAGTGG | 562 bp | 56 °C |
| R: AGCGAGGTGAAGAACGAGAA | |||
| hmpA | F: CCAGTGAATTAACGGCAGGT | 654 bp | 55 °C |
| R: CGTGCCCAGTAATGGCTAAT |
References
- Drzewiecka, D. Significance and roles of Proteus spp. bacteria in natural environments. Microb. Ecol. 2016, 72, 741–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, S.; Stickler, D.; Mobley, H.; Shirtliff, M. Complicated catheter-associated urinary tract infections due to Escherichia coli and Proteus mirabilis. Clin. Microbiol. Rev. 2008, 21, 26–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Gaind, R.; Kothari, C.; Sehgal, R.; Shamweel, A.; Thukral, S.S.; Chellani, H.K. VEB-1 extended-spectrum β-lactamase-producing multidrug-resistant Proteus mirabilis sepsis outbreak in a neonatal intensive care unit in India: Clinical and diagnostic implications. JMM Case Rep. 2016, 3, e005056. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Wang, M.; Liu, L.; Zhou, Z.; Wen, S.; Rozalski, A.; Wang, L. 16S-23S rDNA internal transcribed spacer regions in four Proteus species. J. Microbiol. Methods 2009, 77, 109–118. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, S.; Yu, J.; Zhang, H.; Yuan, Z.; Sun, Y.; Zhang, L.; Zhu, Y.; Song, H. An outbreak of Proteus mirabilis food poisoning associated with eating stewed pork balls in brown sauce, Beijing. Food Control 2009, 21, 302305. [Google Scholar] [CrossRef]
- Gong, Z.; Shi, X.; Bai, F.; He, X.; Zhang, H.; Li, Y.; Yu, W.; Lin, Y.; Qiu, Y.; Chen, Q.; et al. Characterization of a novel diarrheagenic strain of Proteus mirabilis associated with food poisoning in China. Front. Microbiol. 2019, 10, 2810. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Lin, Y.; Qiu, Y.; Li, Y.; Jiang, M.; Chen, Q.; Jiang, Y.; Yuan, J.; Cao, H.; Hu, Q.; et al. Comparative screening of digestion tract toxic genes in Proteus mirabilis. PLoS ONE 2016, 11, e0151873. [Google Scholar] [CrossRef] [Green Version]
- Pattanayak, S.; Kumar, P.R.; Sahoo, M.K.; Paul, A.; Sahoo, P.K. First field-based evidence of association of Proteus mirabilis causing large scale mortality in Indian major carp farming. Aquaculture 2018, 495, 435–442. [Google Scholar] [CrossRef]
- Cabassi, C.S.; Taddei, S.; Predari, G.; Galvani, G.; Ghidini, F.; Schiano, E.; Cavirani, S. Bacteriologic findings in ostrich ( struthio camelus ) eggs from farms with reproductive failures. Avian Dis. 2004, 48, 716–722. [Google Scholar] [CrossRef]
- Milagro, F.-D.; Monica, C.; Alexandra, G.-A.M.; Pulchérie, G.; Paula, S. Occurrence of Proteus mirabilis associated with two species of venezuelan oysters. Rev. Do Inst. Med. Trop. Sao Paulo 2007, 49, 355–359. [Google Scholar]
- Sun, Y.; Wen, S.; Zhao, L.; Xia, Q.; Wang, H. Association among biofilm formation, virulence gene expression, and antibiotic resistance in Proteus mirabilis isolates from diarrhetic animals in Northeast China. BMC Vet. Res. 2020, 16, 176. [Google Scholar] [CrossRef] [PubMed]
- Malayeri, H.Z.; Jamshidi, S.; Salehi, T.Z. Identification and antimicrobial susceptibility patterns of bacteria causing otitis externa in dogs. Vet. Res. Commun. 2010, 34, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, present, and future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghafourian, S.; Sadeghifard, N.; Soheili, S.; Sekawi, Z. Extended spectrum beta-lactamases: Definition, classification and epidemiology. Curr. Issues Mol. Biol. 2015, 17, 11–21. [Google Scholar]
- Tibbetts, R.; Frye, J.G.; Marschall, J.; Warren, D.; Dunne, W. Detection of KPC-2 in a clinical isolate of Proteus mirabilis and first reported description of carbapenemase resistance caused by a KPC beta-lactamase in P. mirabilis. J. Clin. Microbiol. 2008, 46, 3080–3083. [Google Scholar] [CrossRef] [Green Version]
- Cabral, A.B.; Maciel, M.A.; Barros, J.F.; Antunes, M.M.; Lopes, A.C. Detection of blaKPC-2 in Proteus mirabilis in Brazil. Rev. Soc. Bras. Med. Trop. 2015, 48, 94–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girlich, D.; Bonnin, R.A.; Dortet, L.; Naas, T. Genetics of acquired antibiotic resistance genes in Proteus spp. Front. Microbiol. 2020, 11, 256. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Zhang, J.; Wang, H.N.; Lei, C.W. Whole genome sequence of a New Delhi metallo-beta-lactamase 1-producing Proteus mirabilis isolate SNYG35 from broiler chicken in China. J. Glob. Antimicrob. Resist. 2021, 24, 266–269. [Google Scholar] [CrossRef]
- Kang, Q.; Wang, X.; Zhao, J.; Liu, Z.; Ji, F.; Chang, H.; Yang, J.; Hu, S.; Jia, T.; Wang, X.; et al. Multidrug-resistant Proteus mirabilis isolates carrying blaOXA-1 and blaNDM-1 from wildlife in China: Increasing public health risk. Integr. Zool. 2021, 16, 798–809. [Google Scholar] [CrossRef]
- Lei, C.W.; Zhang, A.Y.; Liu, B.H.; Wang, H.N.; Yang, L.Q.; Guan, Z.B.; Xu, C.W.; Zhang, D.D.; Yang, Y.Q. Two novel Salmonella genomic island 1 variants in Proteus mirabilis isolates from swine farms in China. Antimicrob. Agents Chemother. 2015, 59, 4336–4338. [Google Scholar] [CrossRef] [Green Version]
- Algammal, A.M.; Hashem, H.R.; Alfifi, K.J.; Hetta, H.F.; Sheraba, N.S.; Ramadan, H.; El-Tarabili, R.M. atpD gene sequencing, multidrug resistance traits, virulence-determinants, and antimicrobial resistance genes of emerging XDR and MDR-Proteus mirabilis. Sci. Rep. 2021, 11, 9476. [Google Scholar] [CrossRef] [PubMed]
- Leulmi, Z.; Kandouli, C.; Mihoubi, I.; Benlabed, K.; Lezzar, A.; Rolain, J.M. First report of blaOXA-24 carbapenemase gene, armA methyltransferase and aac(6′)-Ib-cr among multidrug-resistant clinical isolates of Proteus mirabilis in Algeria. J. Glob. Antimicrob. Resist 2019, 16, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Al Laham, N.; Chavda, K.D.; Mediavilla, J.R.; Jacobs, M.R.; Bonomo, R.A.; Kreiswirth, B.N. First report of an OXA-48-producing multidrug-resistant Proteus mirabilis strain from Gaza, Palestine. Antimicrob. Agents Chemother. 2015, 59, 4305–4307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.M.; Caroline, R.d.S.; Carvalho, S.L.; Hugo, M.V.; Gabriel, L.B.M.; Henrique, M.G.G.; Henrique, D.d.O.B.; Kenji, N.E.; Carla, F.G.L.; Carolina, V.E.; et al. Proteus mirabilis from community-acquired urinary tract infections (UTI-CA) shares genetic similarity and virulence factors with isolates from chicken, beef and pork meat. Microb. Pathog. 2021, 158, 105098. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing 27th ed CLSI Supplement M100-S27; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Clinical Laboratory Standards Institute. Performance Standards for Antimicrobial Disk Susceptibility Tests for Bacteria Isolated from Animals: CLSI Supplement VET01S; Replaces VET01-S2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Ahmed, A.M.; Yusuke, M.; Maiko, S.; Akito, M.; Hitoshi, W.; Yukio, F.; Tadashi, S. Zoo animals as reservoirs of gram-negative bacteria harboring integrons and antimicrobial resistance genes. Appl. Environ. Microbiol. 2007, 73, 6686–6690. [Google Scholar] [CrossRef] [Green Version]
- Puah, S.-M.; Puthucheary, S.D.; Liew, F.-Y.; Chua, K.-H. Aeromonas aquariorum clinical isolates: Antimicrobial profiles, plasmids and genetic determinants. Int. J. Antimicrob. Agents 2013, 41, 281–284. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Hou, C.; Yang, F. Drug-resistant gene of blaOXA-23, blaOXA-24, blaOXA-51 and blaOXA-58 in Acinetobacter baumannii. Int. J. Clin. Exp. Med. 2015, 8, 13859–13863. [Google Scholar]
- Oliveira, M.; Nunes, M.; Barreto Crespo, M.T.; Silva, A.F. The environmental contribution to the dissemination of carbapenem and (fluoro)quinolone resistance genes by discharged and reused wastewater effluents: The role of cellular and extracellular DNA. Water Res. 2020, 182, 116011. [Google Scholar] [CrossRef]
- Navajas-Benito, E.V.; Alonso, C.A.; Sanz, S.; Olarte, C.; Martínez-Olarte, R.; Hidalgo-Sanz, S.; Somalo, S.; Torres, C. Molecular characterization of antibiotic resistance in Escherichia coli strains from a dairy cattle farm and its surroundings. J. Sci. Food Agric. 2017, 97, 362–365. [Google Scholar] [CrossRef]
- Sarhaddi, N.; Soleimanpour, S.; Farsiani, H.; Mosavat, A.; Dolatabadi, S.; Salimizand, H.; Jamehdar, S.A. Elevated prevalence of multidrug-resistant Acinetobacter baumannii with extensive genetic diversity in the largest burn centre of northeast Iran. J. Glob. Antimicrob. Resist. 2017, 8, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.C.; Lijun, Z.; Thi, T.T.V.; Hoang, S.H.P.; Tran, N.T.; Qi, H.; Rui, Z. Association between the phenotypes and genotypes of antimicrobial resistance in Haemophilus parasuis isolates from swine in Quang Binh and Thua Thien Hue Provinces, Vietnam. Engineering 2020, 6, 40–48. [Google Scholar] [CrossRef]
- Ping, L.X.; Jing, X.; Lei, Y.; Liang, L.; Jian, S.; Hong, L.Y.; Xia, J.H. Characterization of CTX-M-14-producing Escherichia coli from food-producing animals. Front. Microbiol. 2015, 6, 1136. [Google Scholar]
- Chu, C.; Chiu, C.; Wu, W.; Chu, C.; Liu, T.; Ou, J. Large drug resistance virulence plasmids of clinical isolates of Salmonella enterica serovar Choleraesuis. Antimicrob. Agents Chemother. 2001, 45, 2299–2303. [Google Scholar] [CrossRef] [Green Version]
- Guerra, B.; Soto, S.; Argüelles, J.; Mendoza, M. Multidrug resistance is mediated by large plasmids carrying a class 1 integron in the emergent Salmonella enterica serotype [4,5,12:i:-]. Antimicrob. Agents Chemother. 2001, 45, 1305–1308. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhao, S.; White, D.G.; Schroeder, C.M.; Lu, R.; Yang, H.; McDermott, P.F.; Ayers, S.; Meng, J. Characterization of multiple-antimicrobial-resistant salmonella serovars isolated from retail meats. Appl. Environ. Microbiol. 2004, 70, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dashti, A.A.; Jadaon, M.M.; Abdulsamad, A.M.; Dashti, H.M. Heat treatment of bacteria: A simple method of DNA extraction for molecular techniques. Kuwait Med. J. 2009, 41, 117–122. [Google Scholar]
- YouQin, Y.; ChunHua, H.; YiLong, C.; Yun, S.; XiaoWei, H.; DeHui, M. Comparative study on pathogenicity in mice and carriage of virulence genes of Proteus mirabilis isolates from various origins. Chin. J. Biol. 2019, 32, 289–294. [Google Scholar]
- Hernandez, R.J.; Hesse, E.; Dowling, A.J.; Coyle, N.M.; Feil, E.J.; Gaze, W.H.; Vos, M. Using the wax moth larva Galleria mellonella infection model to detect emerging bacterial pathogens. PeerJ 2019, 6, e6150. [Google Scholar] [CrossRef] [Green Version]
- Peleg, O.; Eliopoulos, G.; Moellering, R. Galleria mellonella as a model system to study Acinetobacter baumannii pathogenesis and therapeutics. In Proceedings of the Infectious Diseases Society of America 2008 Annual Meeting, Washington, DC, USA, 25–28 October 2008; pp. 2605–2609. [Google Scholar]
- Finney, D.J. The median lethal dose and its estimation. Arch. Toxicol. 1985, 56, 215–218. [Google Scholar] [CrossRef]
- Yang, B.; Wang, Y.; Qian, P.Y. Sensitivity and correlation of hypervariable regions in 16S rRNA genes in phylogenetic analysis. BMC Bioinform. 2016, 17, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanches, M.S.; Baptista, A.A.; de Souza, M.; Menck-Costa, M.F.; Koga, V.L.; Kobayashi, R.K.; Rocha, S.P. Genotypic and phenotypic profiles of virulence factors and antimicrobial resistance of Proteus mirabilis isolated from chicken carcasses: Potential zoonotic risk. Braz. J. Microbiol. 2019, 50, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Thi, N.N.; Niwat, C.; J, C.-M.J. Antimicrobial resistance in bacterial poultry pathogens: A review. Front. Vet. Sci. 2017, 4, 126. [Google Scholar]
- Ahn, J.Y.; Ann, H.W.; Jeon, Y.; Ahn, M.Y.; Oh, D.H.; Kim, Y.C.; Kim, E.J.; Song, J.E.; Jung, I.Y.; Kim, M.H.; et al. The impact of production of extended-spectrum beta-lactamases on the 28-day mortality rate of patients with Proteus mirabilis bacteremia in Korea. BMC Infect. Dis. 2017, 17, 327. [Google Scholar] [CrossRef] [Green Version]
- Girlich, D.; Dortet, L.; Poirel, L.; Nordmann, P. Integration of the blaNDM-1 carbapenemase gene into Proteus genomic island 1 (PGI1-PmPEL) in a Proteus mirabilis clinical isolate. J. Antimicrob. Chemother. 2015, 70, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Hammoudi Halat, D.; Ayoub Moubareck, C. The current burden of carbapenemases: Review of significant properties and dissemination among Gram-negative bacteria. Antibiotics 2020, 9, 186. [Google Scholar] [CrossRef]
- Sofia, M.; George, S.; Karageorgopoulos, D.E.; Mavros, M.N.; Diamantis, K.; Falagas, M.E. In vitro antimicrobial susceptibility to isepamicin of 6,296 Enterobacteriaceae clinical isolates collected at a tertiary care university hospital in Greece. Antimicrob. Agents Chemother. 2012, 56, 3067–3073. [Google Scholar]
- Pasom, W.; Chanawong, A.; Lulitanond, A.; Wilailuckana, C.; Kenprom, S.; Puang-Ngern, P. Plasmid-mediated quinolone resistance genes, aac(6′)-Ib-cr, qnrS, qnrB, and qnrA, in urinary isolates of Escherichia coli and Klebsiella pneumoniae at a teaching hospital, Thailand. Jpn. J. Infect. Dis. 2013, 66, 428–432. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Cai, J.; Zhang, R.; Zhou, H.; Sun, Q.; Chen, G. Emergence of Proteus mirabilis harboring blaKPC-2 and qnrD in a Chinese Hospital. Antimicrob. Agents Chemother. 2012, 56, 2278–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Zhang, J.; Xu, L.; Liu, Y.; Li, P.; Zhu, T.; Cheng, C.; Lu, S.; Xu, T.; Yi, H.; et al. Spread of the florfenicol resistance floR gene among clinical Klebsiella pneumoniae isolates in China. Antimicrob. Resist. Infect. Control. 2018, 7, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrino, R.; Scavone, P.; Umpiérrez, A.; Maskell, D.J.; Zunino, P. Proteus mirabilis uroepithelial cell adhesin (UCA) fimbria plays a role in the colonization of the urinary tract. Pathog. Dis. 2013, 67, 104–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Insua, J.L.; Llobet, E.; Moranta, D.; Pérez-Gutiérrez, C.; Tomás, A.; Garmendia, J.; Bengoechea, J.A. Modeling Klebsiella pneumoniae pathogenesis by infection of the wax moth Galleria mellonella. Infect. Immun. 2013, 81, 3552–3565. [Google Scholar] [CrossRef] [Green Version]





| Isolates | Year of Isolation | Source | |
|---|---|---|---|
| Animal | Farm Environment 1 | ||
| F1-F22 | 2019 | 12 isolates from fox feces | 9 isolates from soil samples 1 isolate from feed sample |
| R1-R18 | 2019 | 15 isolates from raccoon feces | 3 isolates from soil samples |
| M1-M13 | 2020 | 6 isolates from mink feces, 2 isolates from carcass samples, 3 isolates from throat samples, 1 isolate from annal sample | 1 isolate from feed sample |
| Antibiotic Classes | Antibiotics | Number of Isolates | ||
|---|---|---|---|---|
| Resistant | Intermediate | Susceptible | ||
| Cephems | Cefepime (FEP) | 0 | 1 (1.89%) | 52 (98.11%) |
| Cefotaxime (CTX) | 26 (49.06%) | 8 (15.10%) | 19 (35.85%) | |
| Ceftazidime (CAZ) | 5 (9.43%) | 0 | 48 (90.57%) | |
| Penicillins | Ampicillin (AMP) | 33 (63.46%) | 11 (20.75%) | 9 (16.98%) |
| Carbapenems | Imipenem (IPM) | 38 (71.70%) | 0 | 15 (28.30%) |
| Aminoglycosides | Gentamicin (GM) | 27 (50.94%) | 1 (1.89%) | 25 (47.17%) |
| Streptomycin (STR) | 32 (60.38%) | 13 (24.53%) | 8 (15.10%) | |
| Quinolones | Ofloxacin (OFX) | 9 (16.98%) | 8 (15.09%) | 36 (67.92%) |
| Enrofloxacin (ENR) | 34 (64.15%) | 14 (26.42%) | 5 (9.43%) | |
| Folate pathway antagonists | Trimethoprim-sulfamethoxazole (SXT) | 39 (73.58%) | 1 (1.89%) | 13 (24.53%) |
| Phenicols | Florfenicol (FFC) | 48 (90.57%) | 4 (7.55%) | 1 (1.89%) |
| Name | Gene | Detection Rate | |||
|---|---|---|---|---|---|
| Raccoon | Fox | Mink | >Total | ||
| Beta-lactams | blaTEM | 33.96% | 41.51% | 7.55% | 83.02% |
| blaSHV | 0 | 0 | 0 | 0 | |
| blaPSE | 0 | 0 | 24.53% | 24.53% | |
| blaOXA-1 | 32.08% | 9.43% | 18.87% | 60.38% | |
| blaOXA-23 | 0 | 0 | 0 | 0 | |
| blaOXA-24 | 0 | 3.78% | 11.32% | 15.09% | |
| blaOXA-58 | 0 | 0 | 0 | 0 | |
| blaOXA-48 | 0 | 0 | 0 | 0 | |
| blaCTX-M | 18.87% | 13.21% | 0 | 32.08% | |
| blaKPC | 0 | 0 | 0 | 0 | |
| blaNDM | 11.32% | 1.89% | 0 | 13.21% | |
| blaIMP | 0 | 0 | 0 | 0 | |
| blaVIM | 0 | 0 | 0 | 0 | |
| Aminoglycosides | aaC1 | 0 | 0 | 0 | 0 |
| aaC2 | 0 | 0 | 9.43% | 9.43% | |
| aaC3 | 0 | 0 | 0 | 0 | |
| aadA | 28.30% | 32.08% | 22.64% | 83.02% | |
| aadB | 7.56% | 1.89% | 3.78% | 13.23% | |
| aphA6 | 0 | 5.66% | 5.66% | 11.32% | |
| Quinolones | aac(6′)-Ib-cr | 32.08% | 15.09% | 24.53% | 71.70% |
| qnrA | 3.78% | 1.89% | 0 | 5.66% | |
| qnrB | 0 | 0 | 22.64% | 22.64% | |
| qnrC | 9.43% | 5.66% | 0 | 15.09% | |
| qnrS | 0 | 0 | 0 | 0 | |
| oqxA | 0 | 0 | 0 | 0 | |
| Folate pathway antagonists | sul1 | 33.96% | 41.51% | 18.87% | 94.34% |
| sul2 | 0 | 0 | 11.32% | 11.32% | |
| sul3 | 32.08% | 20.75% | 1.89% | 54.72% | |
| Phenicols | cmlA | 0 | 0 | 5.66% | 5.66% |
| floR | 33.96% | 41.51% | 13.21% | 88.68% | |
| Gene | Fox | Raccoon | Mink | Total |
|---|---|---|---|---|
| (n = 22) | (n = 18) | (n = 13) | (n = 53) | |
| ureC | 100% | 100% | 100% | 100% |
| zapA | 100% | 100% | 100% | 100% |
| pmfA | 100% | 100% | 100% | 100% |
| mrpA | 100% | 100% | 100% | 100% |
| atfC | 100% | 94.44% | 100% | 98.11% |
| atfA | 100% | 100% | 100% | 100% |
| ucaA | 45.45% | 33.33% | 61.54% | 45.28% |
| rsbA | 68.18% | 61.11% | 92.31% | 71.70% |
| rsmA | 95.45% | 100% | 84.62% | 94.34% |
| hmpA | 95.45% | 100% | 100% | 98.11% |
| Isolates | Bacteria Dose (CFU/Larvae) | Dead | LD50 (6 h) (CFU/Larvae) | ||
|---|---|---|---|---|---|
| 6 h | 12 h | 24 h | |||
| F13 | 1 × 106 | 9 | 10 | 10 | 2.5 × 104 |
| 1 × 105 | 6 | 10 | 10 | ||
| 1 × 104 | 6 | 10 | 10 | ||
| 1 × 103 | 0 | 10 | 10 | ||
| 1 × 102 | 0 | 10 | 10 | ||
| R8 | 1 × 106 | 9 | 10 | 10 | 2.0 × 104 |
| 1 × 105 | 10 | 10 | 10 | ||
| 1 × 104 | 3 | 10 | 10 | ||
| 1 × 103 | 0 | 10 | 10 | ||
| 1 × 102 | 0 | 10 | 10 | ||
| M1 | 1 × 106 | 9 | 10 | 10 | 3.9 × 105 |
| 1 × 105 | 0 | 10 | 10 | ||
| 1 × 104 | 0 | 10 | 10 | ||
| 1 × 103 | 0 | 9 | 10 | ||
| 1 × 102 | 0 | 6 | 10 | ||
| Tested Strain | Virulence Gene | Bacteria Dose (CFU/Larvae) | Dead | LD50 (6 h) (CFU/Larvae) | ||||
|---|---|---|---|---|---|---|---|---|
| ucaA | rsbA | rsmA | 6 h | 12 h | 24 h | |||
| F2 | + | + | + | 1 × 106 | 10 | 10 | 10 | 2.0 × 104 |
| 1 × 105 | 7 | 10 | 10 | |||||
| 1 × 104 | 5 | 10 | 10 | |||||
| 1 × 103 | 0 | 1 | 3 | |||||
| 1 × 102 | 0 | 0 | 2 | |||||
| F3 | - | - | - | 1 × 106 | 10 | 10 | 10 | 7.9 × 103 |
| 1 × 105 | 9 | 10 | 10 | |||||
| 1 × 104 | 7 | 10 | 10 | |||||
| 1 × 103 | 0 | 10 | 10 | |||||
| 1 × 102 | 0 | 8 | 10 | |||||
| F6 | - | + | + | 1 × 106 | 10 | 10 | 10 | 4.0 × 103 |
| 1 × 105 | 10 | 10 | 10 | |||||
| 1 × 104 | 9 | 10 | 10 | |||||
| 1 × 103 | 0 | 10 | 10 | |||||
| 1 × 102 | 0 | 10 | 10 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, P.; Hao, G.; Cao, Y.; Cui, L.; Wang, G.; Sun, S. Detection of Carbapenem Resistance of Proteus mirabilis Strains Isolated from Foxes, Raccoons and Minks in China. Biology 2022, 11, 292. https://doi.org/10.3390/biology11020292
Lv P, Hao G, Cao Y, Cui L, Wang G, Sun S. Detection of Carbapenem Resistance of Proteus mirabilis Strains Isolated from Foxes, Raccoons and Minks in China. Biology. 2022; 11(2):292. https://doi.org/10.3390/biology11020292
Chicago/Turabian StyleLv, Penghao, Guijuan Hao, Yanli Cao, Lulu Cui, Guisheng Wang, and Shuhong Sun. 2022. "Detection of Carbapenem Resistance of Proteus mirabilis Strains Isolated from Foxes, Raccoons and Minks in China" Biology 11, no. 2: 292. https://doi.org/10.3390/biology11020292