
Plasma Proteomic Changes of Atherosclerosis after Exercise in ApoE Knockout Mice

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Morphological Analysis of Atherosclerotic Plaque
2.3. Proteomic Analysis
2.4. Immunohistochemical Examination
2.5. Statistical Analysis
3. Results
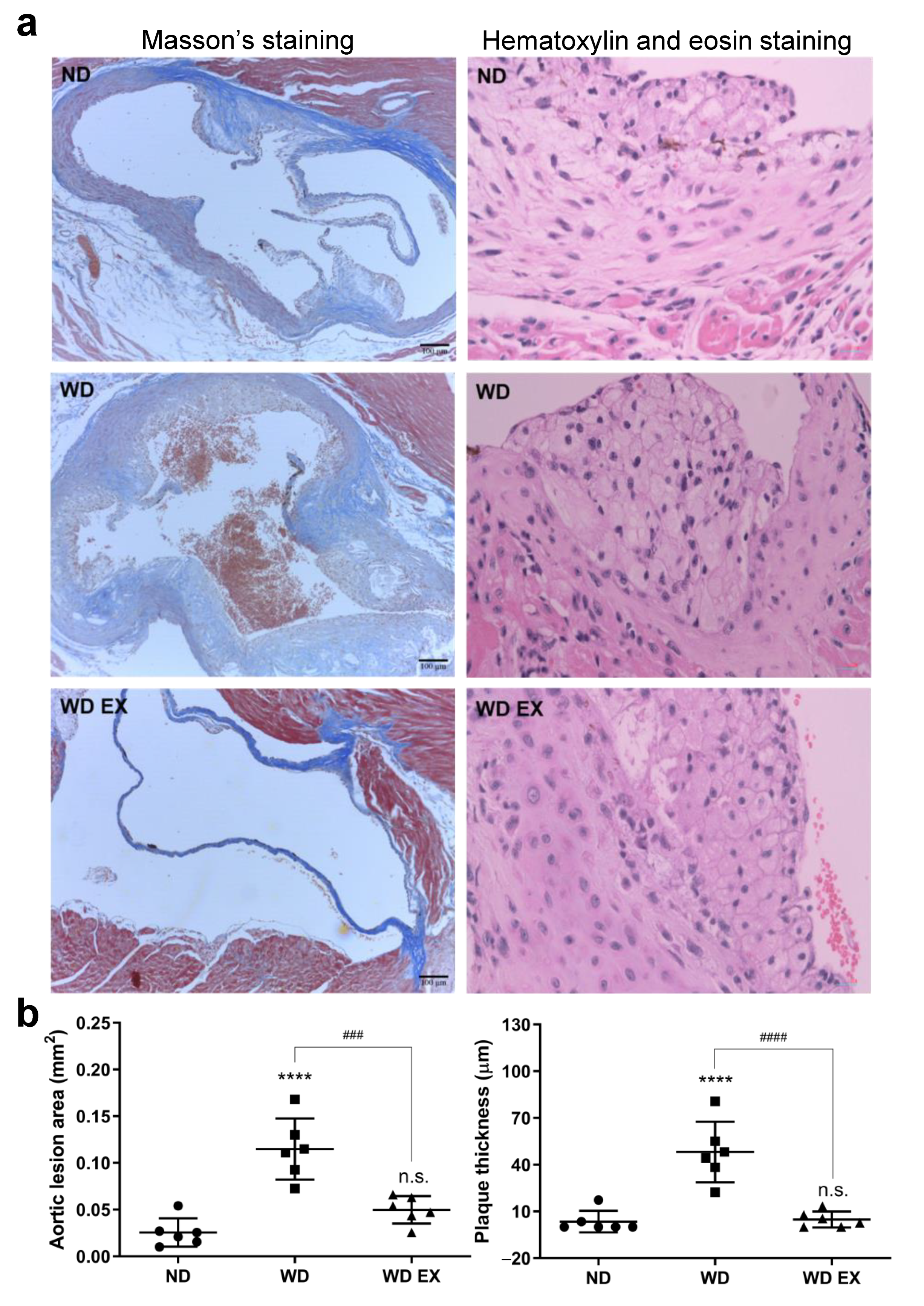
3.1. The Effect of Exercise on the Development of Atherosclerotic Plaque
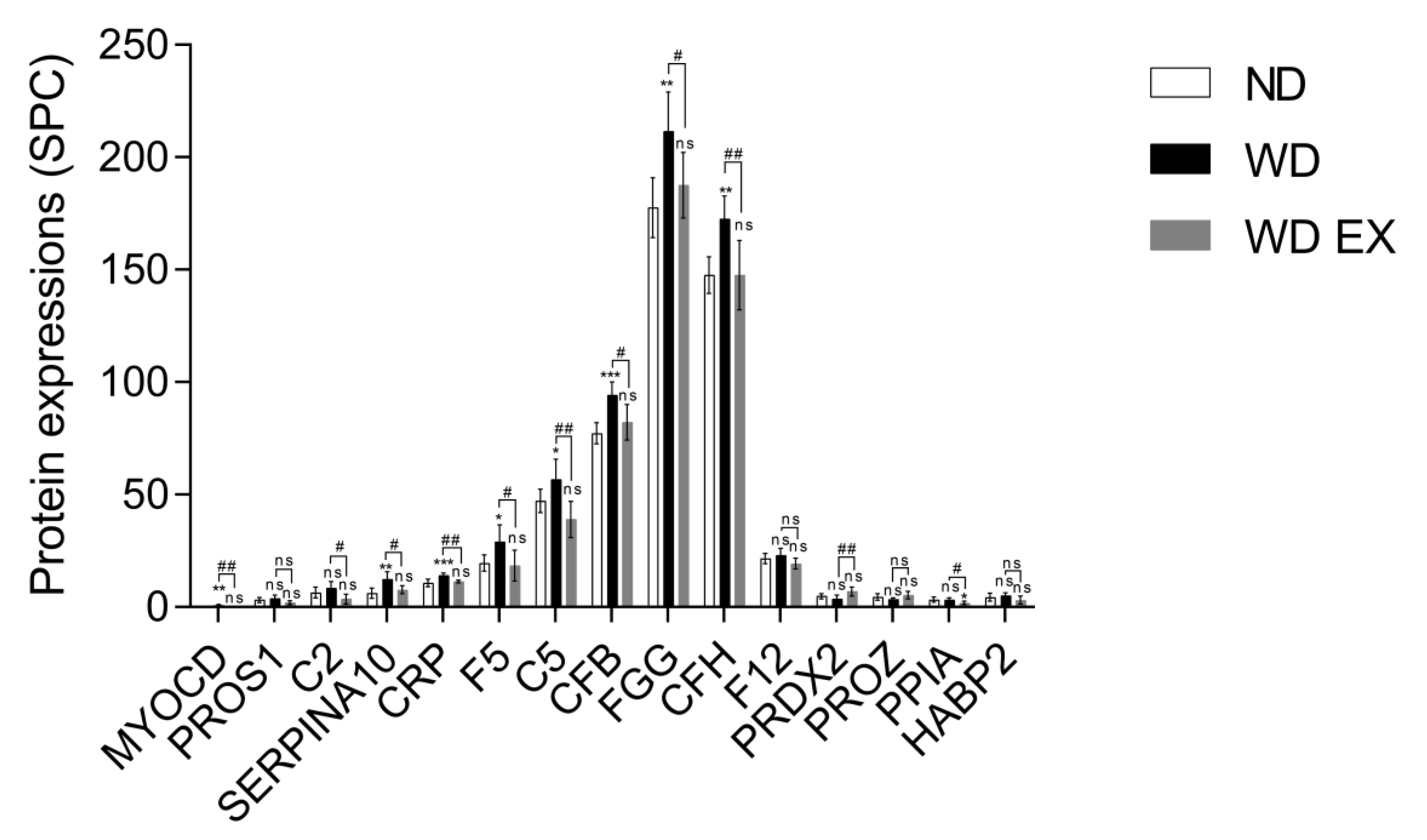
3.2. The Effect of Exercise on Plasma Proteomic Changes
3.3. Effects of Exercise on Plasma Protein–Regulated Biofunction Pathways
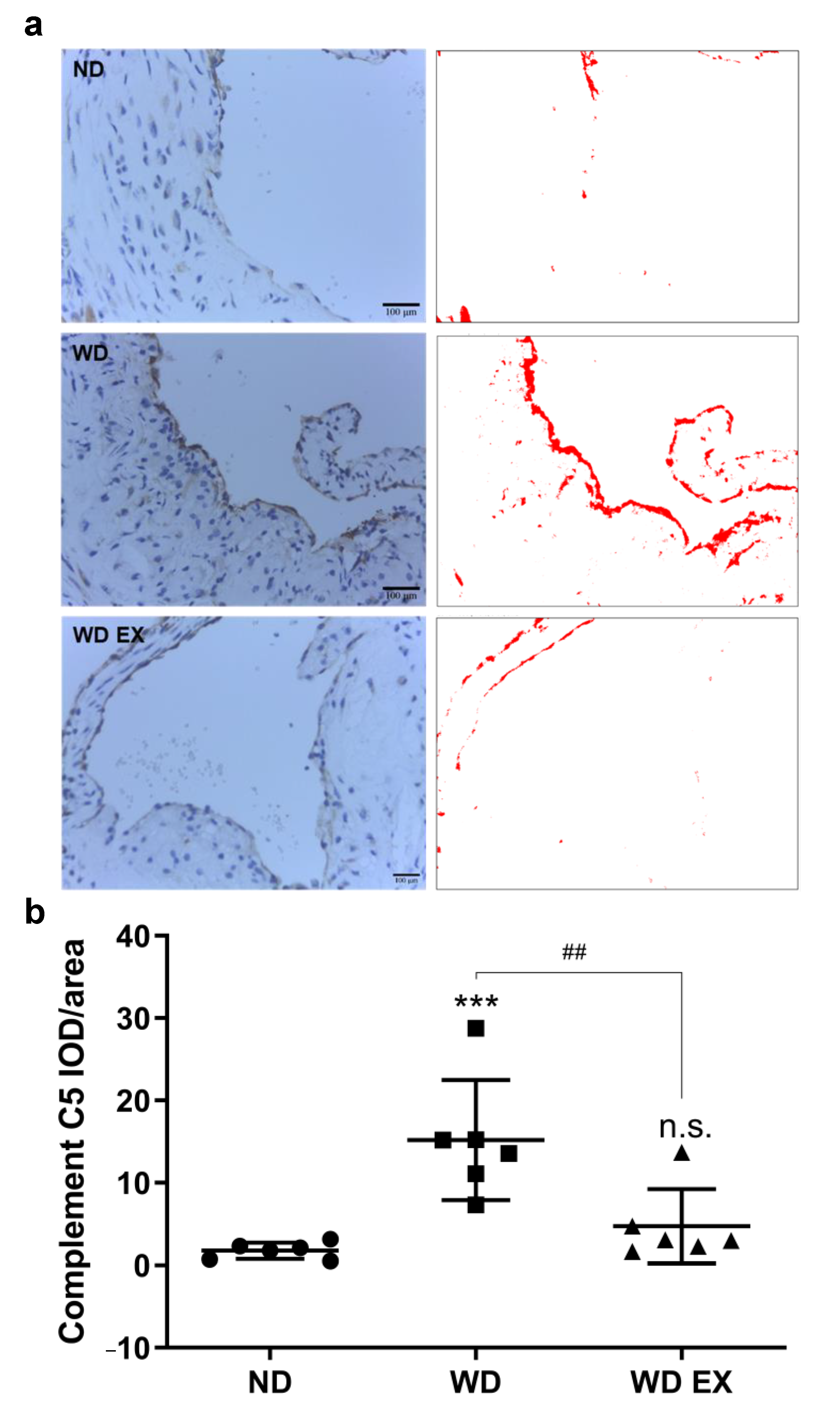
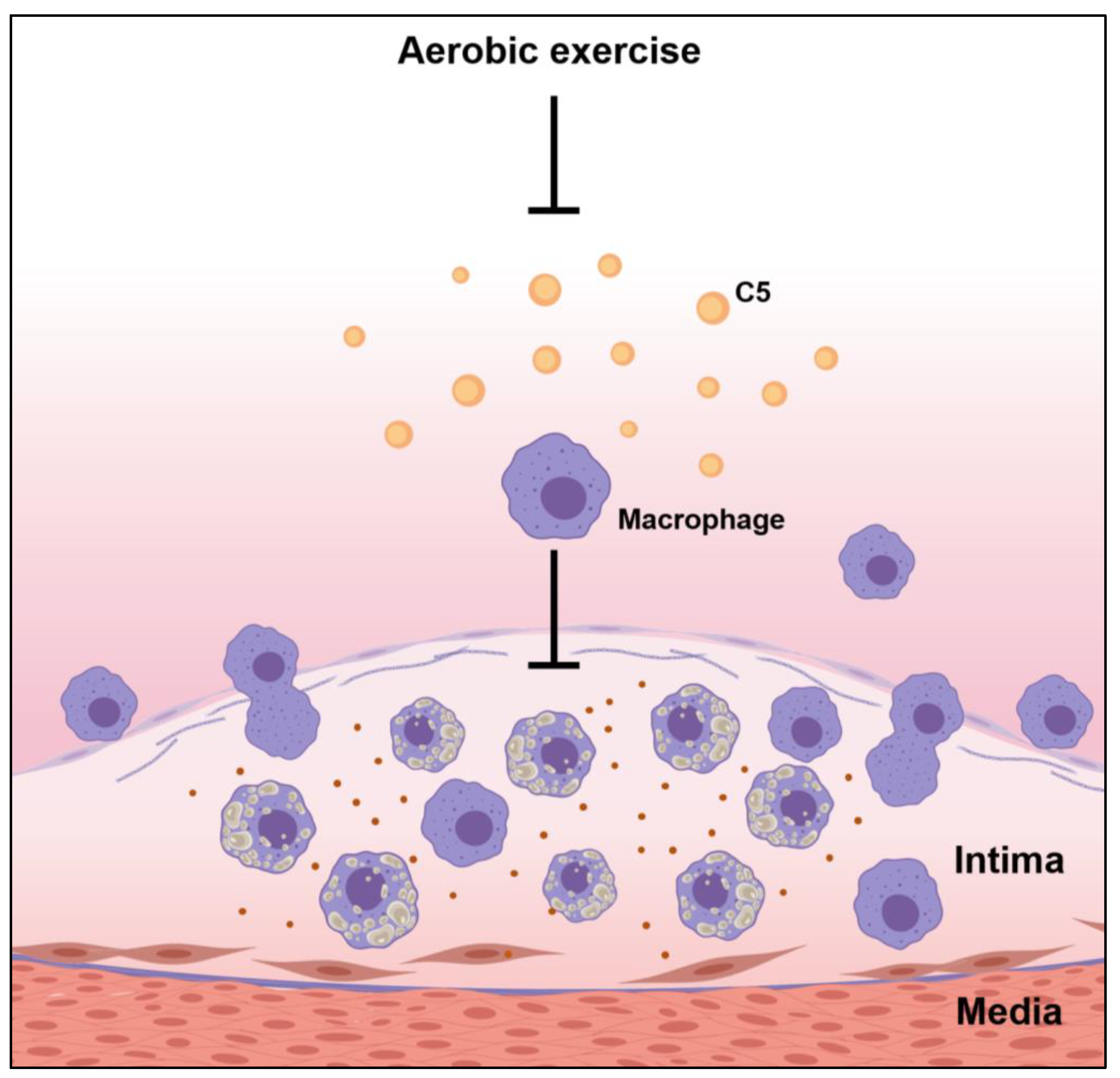
3.4. Effects of Exercise on Complement Factor C5 in the Aortic Root
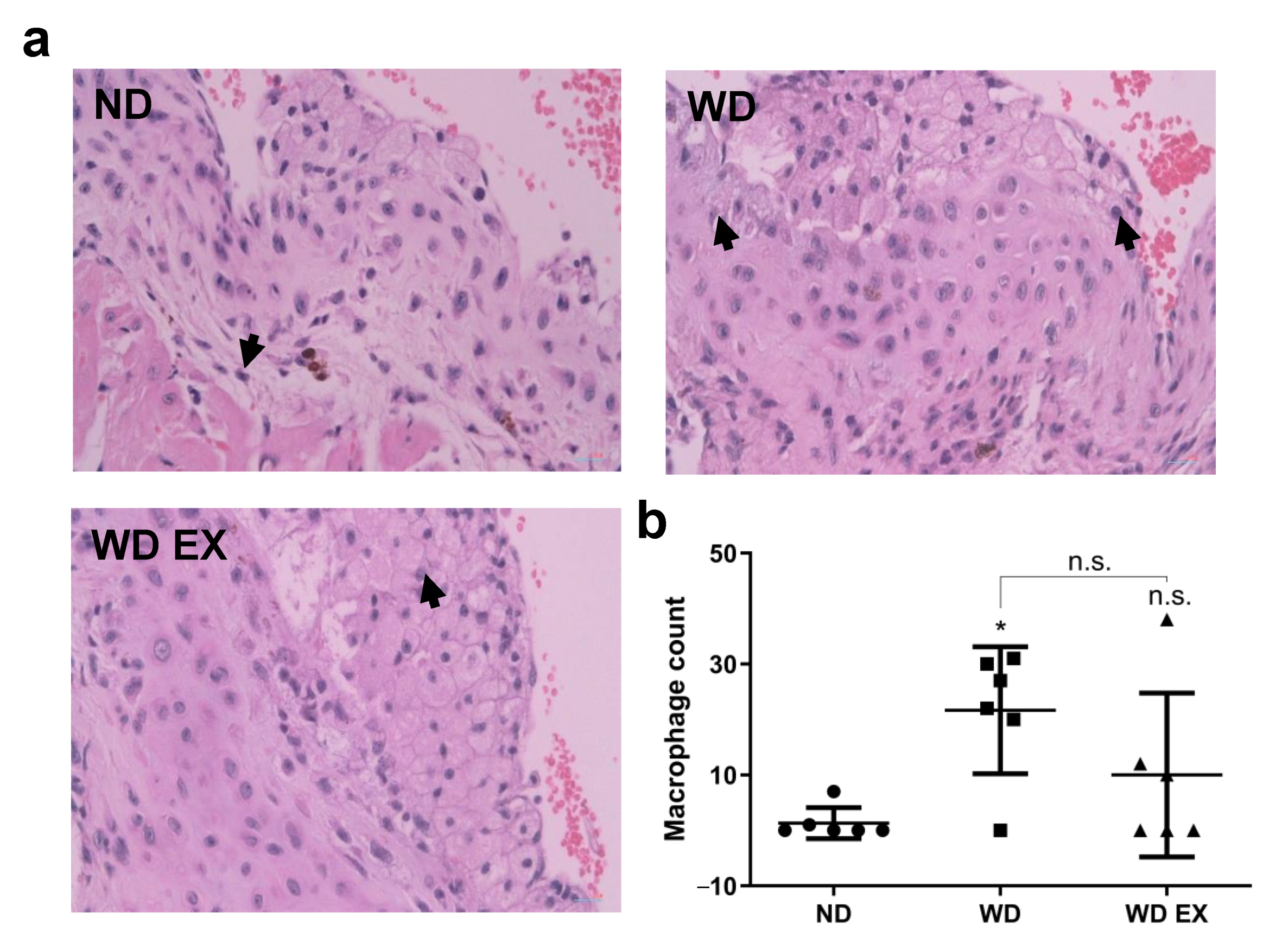
3.5. Effects of Exercise on Macrophage Infiltration of the Cholesterol-Driven Plaque Formed in the Aortic Root
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation List
| Apoeem1Narl/Narl, ApoE knockout | Apolipoprotein E knockout mice |
| CAN | Acetonitrile |
| CID | Collision-induced dissociation |
| CVD | Cardiovascular disease |
| DAB | Diaminobenzidine |
| DTT | Dithiothreitol |
| IAA | Iodoacetamide |
| IOD | Integrated optical density |
| IPA | Ingenuity pathway analysis |
| LC-MS/MS | Liquid chromatography-tandem mass spectrometry |
| ND | Mormal diet |
| WD EX | WD with 12 weeks exercise intervention |
| WD | Western diet |
References
- Frostegard, J. Immunity, atherosclerosis and cardiovascular disease. BMC Med. 2013, 11, 117. [Google Scholar] [CrossRef] [Green Version]
- Lusis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, C.; Chimowitz, M.I.Y. Stroke caused by atherosclerosis of the major intracranial arteries. Circ. Res. 2017, 120, 502–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd-Jones, D.M.; Hong, Y.; Labarthe, D.; Mozaffarian, D.; Appel, L.J.; Van Horn, L.; Greenlund, K.; Daniels, S.; Nichol, G.; Tomaselli, G.F.; et al. Defining and setting national goals for cardiovascular health promotion and disease reduction: The American Heart Association’s strategic impact goal through 2020 and beyond. Circulation 2010, 121, 586–613. [Google Scholar] [CrossRef] [Green Version]
- Chiuve, S.E.; McCullough, M.L.; Sacks, F.M.; Rimm, E.B. Healthy lifestyle factors in the primary prevention of coronary heart disease among men: Benefits among users and nonusers of lipid-lowering and antihypertensive medications. Circulation 2006, 114, 160–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stampfer, M.J.; Hu, F.B.; Manson, J.E.; Rimm, E.B.; Willett, W.C. Primary prevention of coronary heart disease in women through diet and lifestyle. N. Engl. J. Med. 2000, 343, 16–22. [Google Scholar] [CrossRef]
- Thompson, P.D.; Buchner, D.; Piña, I.L.; Balady, G.J.; Williams, M.A.; Marcus, B.H.; Berra, K.; Blair, S.N.; Costa, F.; Franklin, B.; et al. Exercise and physical activity in the prevention and treatment of atherosclerotic cardiovascular disease: A statement from the Council on Clinical Cardiology (Subcommittee on Exercise, Rehabilitation, and Prevention) and the Council on Nutrition, Physical Activity, and Metabolism (Subcommittee on Physical Activity). Circulation 2003, 107, 3109–3116. [Google Scholar]
- Church, T. Exercise in obesity, metabolic syndrome, and diabetes. Prog. Cardiovasc. Dis. 2011, 53, 412–418. [Google Scholar] [CrossRef]
- Tsukiyama, Y.; Ito, T.; Nagaoka, K.; Eguchi, E.; Ogino, K. Effects of exercise training on nitric oxide, blood pressure and antioxidant enzymes. J. Clin. Biochem. Nutr. 2017, 60, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef]
- Brown, T.M.; Bittner, V. Biomarkers of atherosclerosis: Clinical applications. Curr. Cardiol. Rep. 2008, 10, 497–504. [Google Scholar] [CrossRef]
- Wilson, P.W.; D’Agostino, R.B.; Levy, D.; Belanger, A.M.; Silbershatz, H.; Kannel, W.B. Prediction of coronary heart disease using risk factor categories. Circulation 1998, 97, 1837–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, Y.; Plump, A.S.; Raines, E.W.; Breslow, J.L.; Ross, R. ApoE-deficient mice develop lesions of all phases of atherosclerosis throughout the arterial tree. Arterioscler. Thromb. 1994, 14, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawień, J.; Nastałek, P.; Korbut, R. Korbut Mouse models of experimental atherosclerosis. J. Physiol. Pharmacol. 2004, 55, 503–517. [Google Scholar]
- Pellegrin, M.; Berthelot, A.; Houdayer, C.; Gaume, V.; Deckert, V.; Laurant, P. New insights into the vascular mechanisms underlying the beneficial effect of swimming training on the endothelial vasodilator function in apolipoprotein E-deficient mice. Atherosclerosis 2007, 190, 35–42. [Google Scholar] [CrossRef]
- Liao, C.C.; Chen, Y.W.; Jeng, T.L.; Li, C.R.; Kuo, C.F. Consumption of Purple Sweet Potato Affects Post-Translational Modification of Plasma Proteins in Hamsters. J. Agric. Food Chem. 2013, 61, 12450–12458. [Google Scholar] [CrossRef]
- Uen, Y.H.; Lin, K.Y.; Sun, D.P.; Liao, C.C.; Hsieh, M.S.; Huang, Y.K.; Chen, Y.W.; Huang, P.H.; Chen, W.J.; Tai, C.C.; et al. Comparative proteomics, network analysis and post-translational modification identification reveal differential profiles of plasma Con A-bound glycoprotein biomarkers in gastric cancer. J. Proteom. 2013, 27, 197–213. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Speidl, W.S.; Kastl, S.P.; Huber, K.; Wojta, J. Complement in atherosclerosis: Friend or foe? J. Thromb. Haemost. 2011, 9, 428–440. [Google Scholar] [CrossRef]
- Froelicher, V.F. Animal studies of effect of chronic exercise on the heart and atherosclerosis: A review. Am. Heart J. 1972, 84, 496–506. [Google Scholar] [CrossRef]
- Kramsch, D.M.; Aspen, A.J.; Abramowitz, B.M.; Kreimendahl, T.; Hood, W.B., Jr. Reduction of coronary atherosclerosis by moderate conditioning exercise in monkeys on an atherogenic diet. N. Engl. J. Med. 1981, 305, 1483–1489. [Google Scholar] [CrossRef]
- Shephard, R.J.; Balady, G.J. Exercise as cardiovascular therapy. Circulation 1999, 99, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Jakic, B.; Carlsson, M.; Buszko, M.; Cappellano, G.; Ploner, C.; Onestingel, E.; Foti, M.; Hackl, H.; Demetz, E.; Dietrich, H.; et al. The effects of aerobic exercise and diet on atherosclerosis in young and aged ApoE-/- and wild-type mice. Gerontology 2019, 65, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Cho, J.Y.; Kim, W.K. Anti-inflammation effect of exercise and Korean red ginseng in aging model rats with diet-induced atherosclerosis. Nutr. Res. Pract. 2014, 8, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Okabe, T.A.; Shimada, K.; Hattori, M.; Murayama, T.; Yokode, M.; Kita, T.; Kishimoto, C. Swimming reduces the severity of atherosclerosis in apolipoprotein E deficient mice by antioxidant effects. Cardiovasc. Res. 2007, 74, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Szostak, J.; Miguet-Alfonsi, C.; Berthelot, A.; Laurant, P. Training-induced anti-atherosclerotic effects are associated with increased vascular PPAR gamma expression in apolipoprotein E-deficient mice. Acta Physiol. 2016, 216, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Amiri-Dashatan, N.; Koushki, M.; Abbaszadeh, H.A.; Rostami-Nejad, M.; Rezaei-Tavirani, M. Proteomics applications in health: Biomarker and drug discovery and food industry. Iran. J. Pharm. Res. 2018, 17, 1523–1536. [Google Scholar]
- Previtali, E.; Bucciarelli, P.; Passamonti, S.M.; Martinelli, I. Risk factors for venous and arterial thrombosis. Blood Transfus. 2011, 9, 120. [Google Scholar] [PubMed]
- Oksjoki, R.; Kovanen, P.T.; Meri, S.; Pentikainen, M.O. Function and regulation of the complement system in cardiovascular diseases. Front. Biosci. 2007, 12, 4696–4708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, T.H.; Cortini, A.; Carassiti, D.; Boyle, J.J.; Haskard, D.O.; Botto, M. The alternative pathway is critical for pathogenic complement activation in endotoxin- and diet-induced atherosclerosis in low-density lipoprotein receptor-deficient mice. Circulation 2010, 122, 1948–1956. [Google Scholar] [CrossRef]
- Reynolds, G.D.; Vance, R.P. C-reactive protein immunohistochemical localization in normal and atherosclerotic human aortas. Arch. Pathol. Lab. Med. 1987, 111, 265–269. [Google Scholar]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Meng, W.; Hughes, A.; Patterson, C.C.; Belton, C.; Kamaruddin, M.S.; Horan, P.G.; Kee, F.; McKeown, P.P. Genetic variants of Complement factor H gene are not associated with premature coronary heart disease: A family-based study in the Irish population. BMC. Med. Genet. 2007, 8, 62. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.A.M.; Dantas, P.M.S.; Dos Santos, I.K.; Dantas, M.; da Silva, D.C.P.; Cabral, B.G.A.T.; Guerra, R.O.; Júnior, G.B.C. Effect of acute and chronic aerobic exercise on immunological markers: A Systematic Review. Front. Physiol. 2020, 10, 1602. [Google Scholar] [CrossRef] [PubMed]
- Santos, V.C.; Levada-Pires, A.C.; Alves, S.R.; Pithon-Curi, T.C.; Curi, R.; Cury-Boaventura, M.F. Changes in lymphocyte and neutrophil function induced by a marathon race. Cell Biochem. Funct. 2013, 31, 237–243. [Google Scholar] [CrossRef]
- Wolach, B.; Eliakim, A.; Gavrieli, R.; Kodesh, E.; Yarom, Y.; Schlesinger, M.; Falk, B. Aspects of leukocyte function and the complement system following aerobic exercise in young female gymnasts. J. Med. Sci. Sports 1998, 8, 91–97. [Google Scholar] [CrossRef]
- Amara, U.; Rittirsch, D.; Flierl, M.; Bruckner, U.; Klos, A.; Gebhard, F.; Lambris, J.D.; Huber-Lang, M. Interaction between the coagulation and complement system. Adv. Exp. Med. Biol. 2008, 68–76. [Google Scholar]
- Huber-Lang, M.; Sarma, J.V.; Zetoune, F.S.; Rittirsch, D.; Neff, T.A.; McGuire, S.R.; Lambris, J.D.; Warner, R.L.; Flierl, M.A.; Hoesel, L.M.; et al. Generation of C5a in the absence of C3: A new complement activation pathway. Nat. Med. 2006, 12, 682–687. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Gene ID | Protein Name | Respective Fold Change | p-Value |
|---|---|---|---|---|
| COOA1_MOUSE | Q30D77 | Collagen alpha-1(XXIV) chain | - | - |
| HVM45_MOUSE | P01821 | Ig heavy chain V region MC101 | - | - |
| MYCD_MOUS | Q8VIM5 | Myocardin | - | - |
| CP250_MOUSE | Q60952 | Centrosome-associated protein CEP250 | - | - |
| LOXL1_MOUSE | P97873 | Lysyl oxidase homolog 1 | −7.289 | 0.0220 |
| DYHC1_MOUSE | Q9JHU4 | Cytoplasmic dynein 1 heavy chain 1 | −6.976 | 0.0232 |
| AMPL_MOUSE | Q9CPY7 | Cytosol aminopeptidase | −5.695 | 0.0125 |
| NRP1_MOUSE | P97333 | Neuropilin-1 | −5.124 | 0.0163 |
| BIP_MOUSE | P20029 | Endoplasmic reticulum chaperone BiP | −4.781 | 0.0008 |
| BHMT1_MOUSE | O35490 | Betaine–homocysteine S-methyltransferase 1 | −4.416 | 0.0157 |
| PLSL_MOUSE | Q61233 | Plastin-2 | −3.783 | 0.0186 |
| ACE_MOUSE | P09470 | Angiotensin-converting enzyme | −3.692 | 0.0209 |
| ALDOB_MOUSE | Q91Y97 | Fructose-bisphosphate aldolase B | −3.373 | 0.0220 |
| OBP1A_MOUSE | Q9D3H2 | Odorant-binding protein 1a | −3.369 | 0.0127 |
| SAP_MOUSE | Q61207 | Prosaposin | −3.103 | 0.0206 |
| RETNG_MOUSE | Q8K426 | Resistin-like gamma | −3.020 | 0.0096 |
| HSP7C_MOUSE | P63017 | Heat shock cognate 71 kDa protein | −3.000 | 0.0117 |
| HPT_MOUSE | Q61646 | Haptoglobin | −2.745 | 0.0012 |
| IGF1_MOUSE | P05017 | Insulin-like growth factor I | −2.576 | 0.0395 |
| PSA7_MOUSE | Q9Z2U0 | Proteasome subunit alpha type-7 | −2.451 | 0.0326 |
| CO2_MOUSE | P21180 | Complement C2 | −2.355 | 0.0091 |
| COL11_MOUSE | Q3SXB8 | Collectin-11 | −2.349 | 0.0378 |
| KPYM_MOUSE | P52480 | Pyruvate kinase PKM | −2.280 | 0.0093 |
| LBP_MOUSE | Q61805 | Lipopolysaccharide-binding protein | −2.251 | 0.0029 |
| PROS_MOUSE | Q08761 | Vitamin K-dependent protein S | −1.901 | 0.0336 |
| PI16_MOUSE | Q9ET66 | Peptidase inhibitor 16 | −1.892 | 0.0270 |
| PPIA_MOUSE | P17742 | Peptidyl-prolyl cis-trans isomerase A | −1.787 | 0.0095 |
| TRFL_MOUSE | P08071 | Lactotransferrin | −1.771 | 0.0189 |
| HGFL_MOUSE | P26928 | Hepatocyte growth factor-like protein | −1.717 | 0.0059 |
| HABP2_MOUSE | Q8K0D2 | Hyaluronan-binding protein 2 | −1.684 | 0.0435 |
| A1AG2_MOUSE | P0736 | Alpha-1-acid glycoprotein 2 | −1.629 | 0.0198 |
| ZPI_MOUSE | Q8R121 | Protein Z-dependent protease inhibitor | −1.600 | 0.0222 |
| FA5_MOUSE | O88783 | Coagulation factor V | −1.582 | 0.0282 |
| A1AG1_MOUSE | Q60590 | Alpha-1-acid glycoprotein 1 | −1.542 | 0.0097 |
| ACTBL_MOUSE | Q8BFZ3 | Beta-actin-like protein 2 | −1.542 | 0.0029 |
| CO5_MOUSE | P06684 | Complement C5 | −1.454 | 0.0050 |
| HEMO_MOUSE | Q91 × 72 | Hemopexin | −1.448 | 0.0074 |
| CO8G_MOUSE | Q8VCG4 | Complement component C8 gamma chain | −1.391 | 0.0092 |
| PEDF_MOUSE | P97298 | Pigment epithelium-derived factor | −1.369 | 0.0078 |
| SAA4_MOUSE | P31532 | Serum amyloid A-4 protein | −1.320 | 0.0164 |
| ITIH4_MOUSE | A6 × 935 | Inter alpha-trypsin inhibitor, heavy chain 4 | −1.311 | 0.0134 |
| PGRP2_MOUSE | Q8VCS0 | N-acetylmuramoyl-L-alanine amidase | −1.304 | 0.0173 |
| AMBP_MOUSE | Q07456 | Protein AMBP | −1.304 | 0.0063 |
| ITIH3_MOUSE | Q61704 | Inter-alpha-trypsin inhibitor heavy chain H3 | −1.277 | 0.0389 |
| CRP_MOUSE | P14847 | C-reactive protein | −1.246 | 0.0003 |
| APOA1_MOUSE | Q00623 | Apolipoprotein A-I | −1.236 | 0.0093 |
| FA12_MOUSE | Q80YC5 | Coagulation factor XII | −1.194 | 0.0425 |
| ITIH2_MOUSE | Q61703 | Inter-alpha-trypsin inhibitor heavy chain H2 | −1.193 | 0.0103 |
| CFAH_MOUSE | P06909 | Complement factor H | −1.170 | 0.0094 |
| CO9_MOUSE | P06683 | Complement component C9 | −1.158 | 0.0229 |
| CFAB_MOUSE | P04186 | Complement factor B | −1.147 | 0.0139 |
| ITIH1_MOUSE | Q61702 | Inter-alpha-trypsin inhibitor heavy chain H1 | −1.144 | 0.0071 |
| FETUA_MOUSE | P29699 | Alpha-2-HS-glycoprotein | −1.141 | 0.0428 |
| FIBG_MOUSE | Q8VCM7 | Fibrinogen gamma chain | −1.128 | 0.0280 |
| SPA3G_MOUSE | Q5I2A0 | Serine protease inhibitor A3G | 1.269 | 0.0005 |
| P20D1_MOUSE | Q8C165 | N-fatty-acyl-amino acid synthase/hydrolase PM20D1 | 1.392 | 0.0353 |
| PROZ_MOUSE | Q9CQW3 | Vitamin K-dependent protein Z | 1.545 | 0.0461 |
| PRDX2_MOUSE | Q61171 | Peroxiredoxin-2 | 1.866 | 0.0113 |
| APOM_MOUSE | Q9Z1R3 | Apolipoprotein M | 1.911 | 0.0182 |
| HA1D_MOUSE | P01902 | H-2 class I histocompatibility antigen, K-D alpha chain | 2.176 | 0.0263 |
| HA1W_MOUSE | P03991 | H-2 class I histocompatibility antigen, K-W28 alpha chain | 2.178 | 0.0064 |
| CAMP_MOUSE | P51437 | Cathelicidin antimicrobial peptide | 3.047 | 0.0327 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, C.-C.; Xu, J.-W.; Huang, W.-C.; Chang, H.-C.; Tung, Y.-T. Plasma Proteomic Changes of Atherosclerosis after Exercise in ApoE Knockout Mice. Biology 2022, 11, 253. https://doi.org/10.3390/biology11020253
Liao C-C, Xu J-W, Huang W-C, Chang H-C, Tung Y-T. Plasma Proteomic Changes of Atherosclerosis after Exercise in ApoE Knockout Mice. Biology. 2022; 11(2):253. https://doi.org/10.3390/biology11020253
Chicago/Turabian StyleLiao, Chen-Chung, Jin-Wei Xu, Wen-Ching Huang, Hung-Chang Chang, and Yu-Tang Tung. 2022. "Plasma Proteomic Changes of Atherosclerosis after Exercise in ApoE Knockout Mice" Biology 11, no. 2: 253. https://doi.org/10.3390/biology11020253