

Cultivars Resistance Assay for Maize Late Wilt Disease


Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. The Maize Varieties Selection and Experimental Design
2.2. M. maydis Origin and Growth Conditions
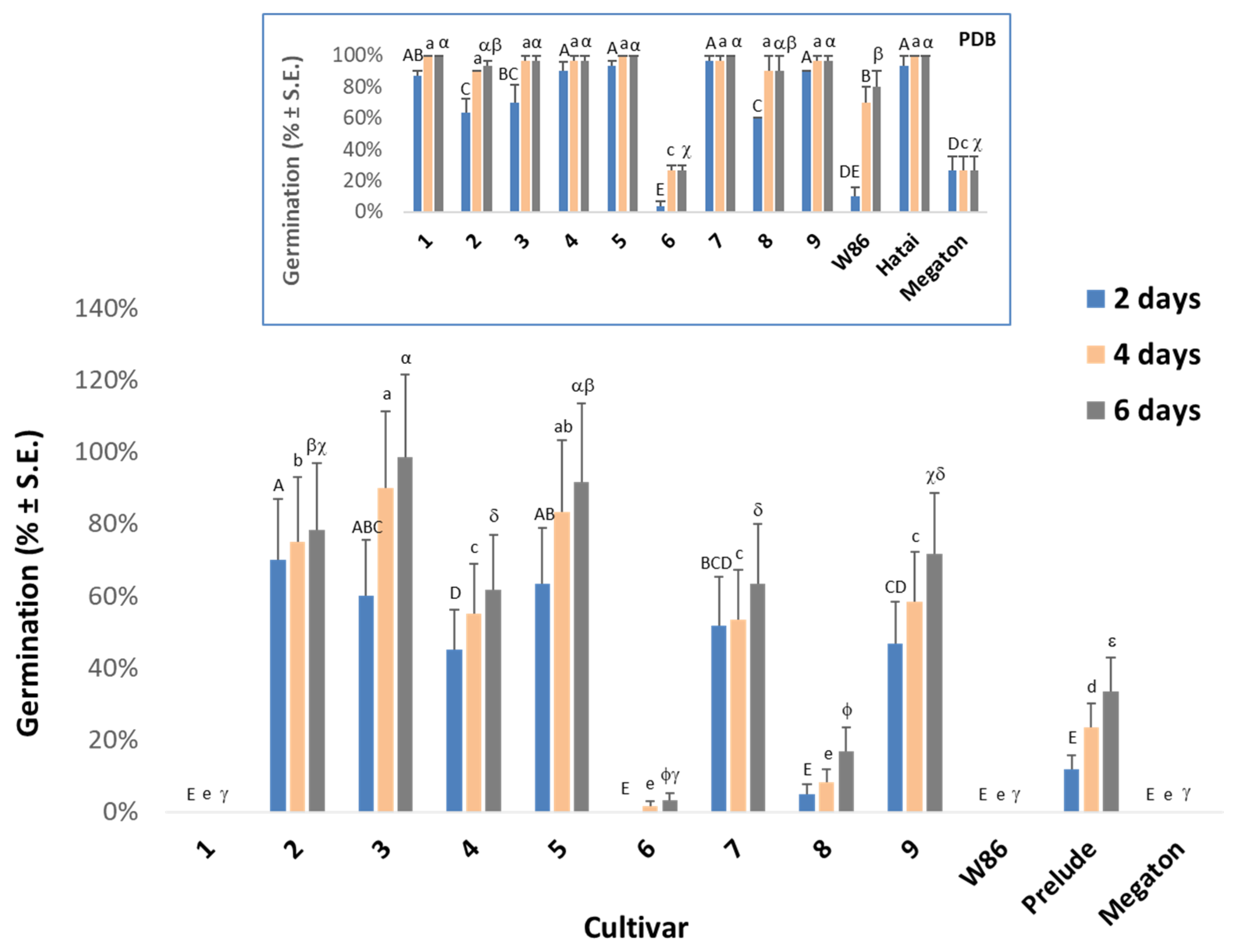
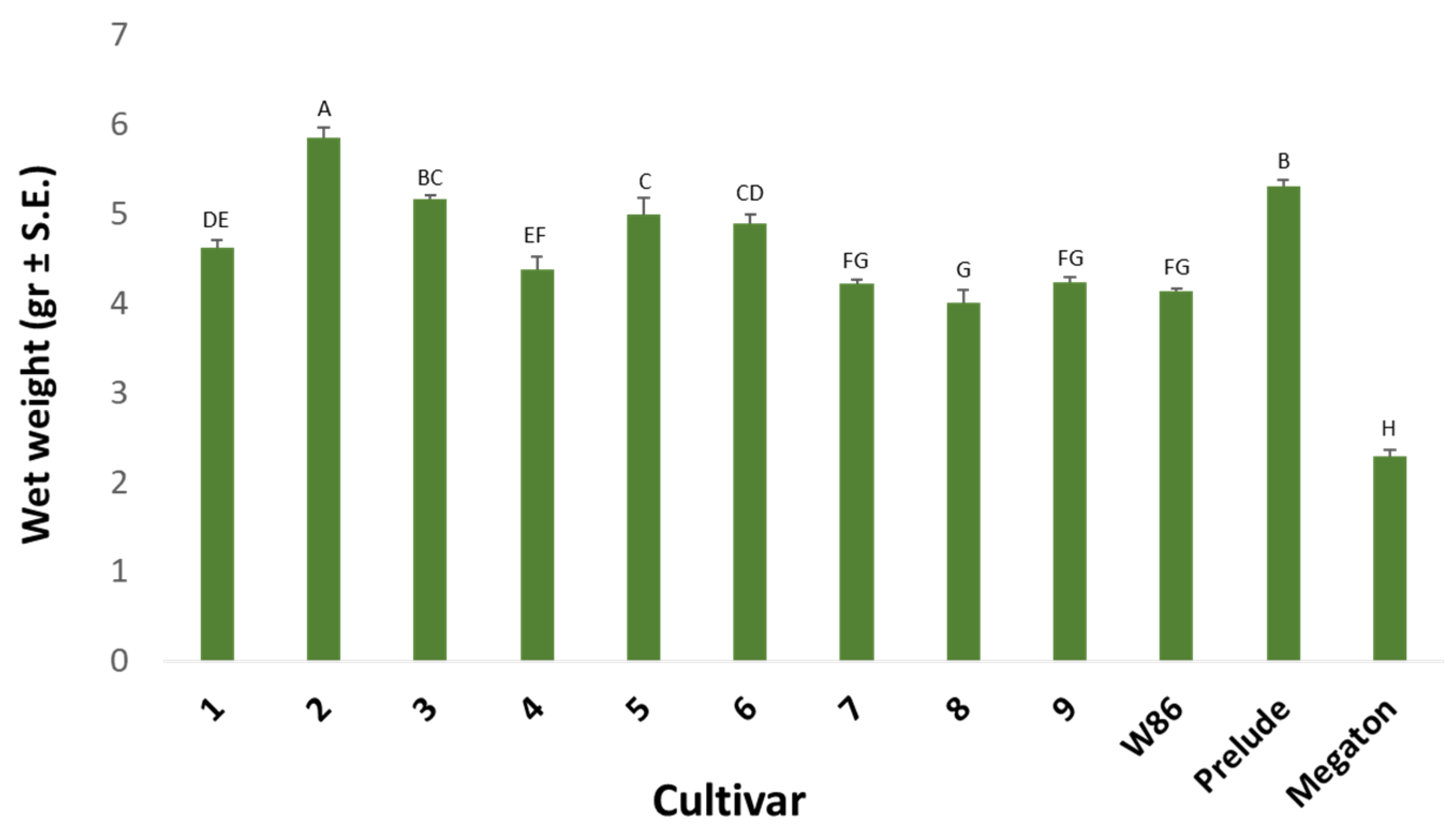
2.3. In Vitro Seed Test
2.4. Pots Assay under Semi-Field Conditions throughout an Entire Growing Season
2.4.1. The Experiment Conditions
2.4.2. Growth and Disease Estimation
2.5. Molecular Real-Time PCR Diagnostic
2.5.1. DNA Extraction
2.5.2. qPCR Technique
- The A200 primers amplify the segment-specific to M. maydis:
- The primers for the COX gene (codes for the enzyme cytochrome oxidase, the last enzyme in the cellular respiratory chain in the mitochondria) that is used as a housekeeping gene to normalize M. maydis DNA amount according to the ΔCt model:
2.6. Statistical Analysis
3. Results
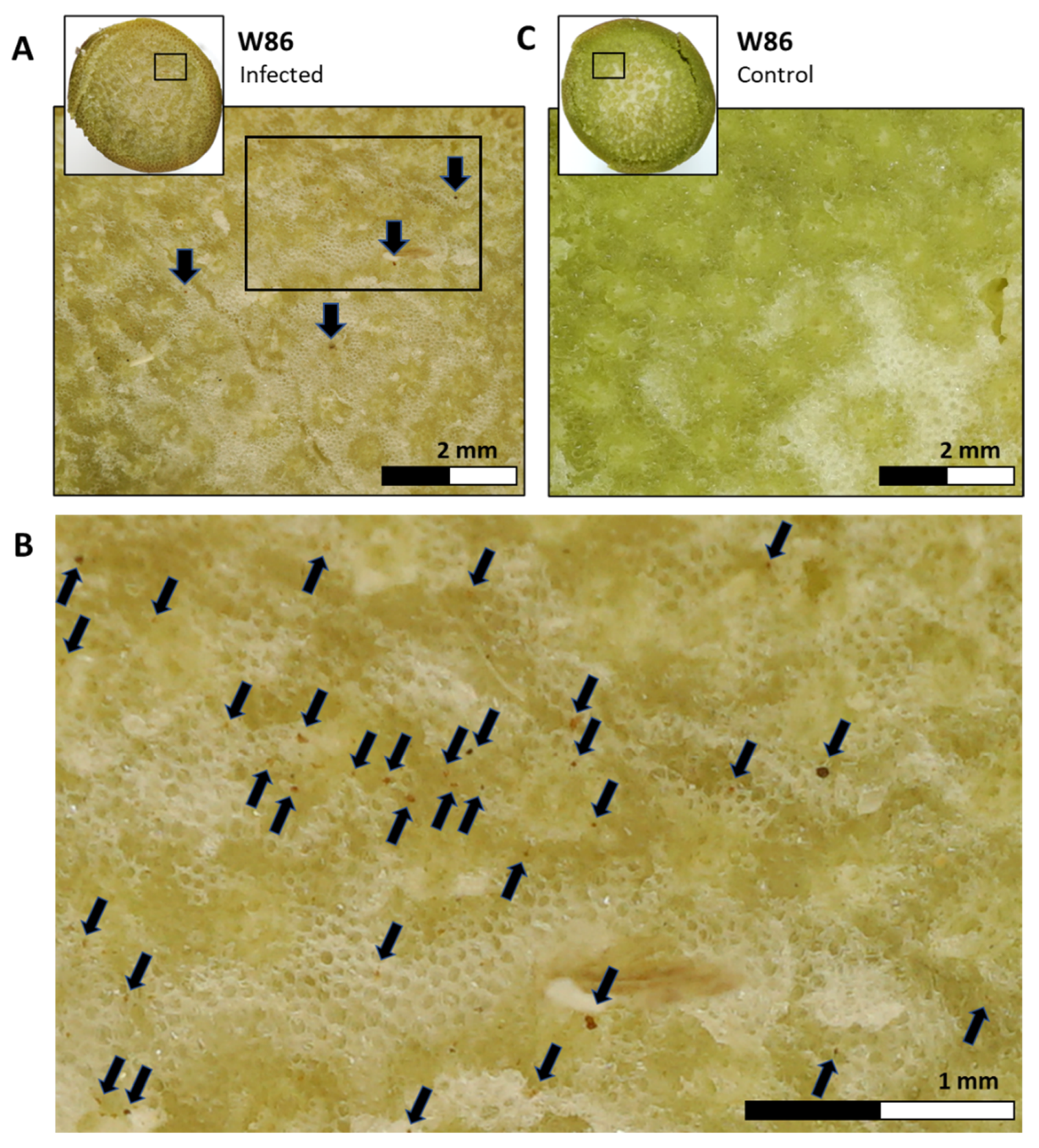
3.1. In Vitro Seed Test
3.2. Pot Assay under Semi-Field Conditions throughout an Entire Growing Season
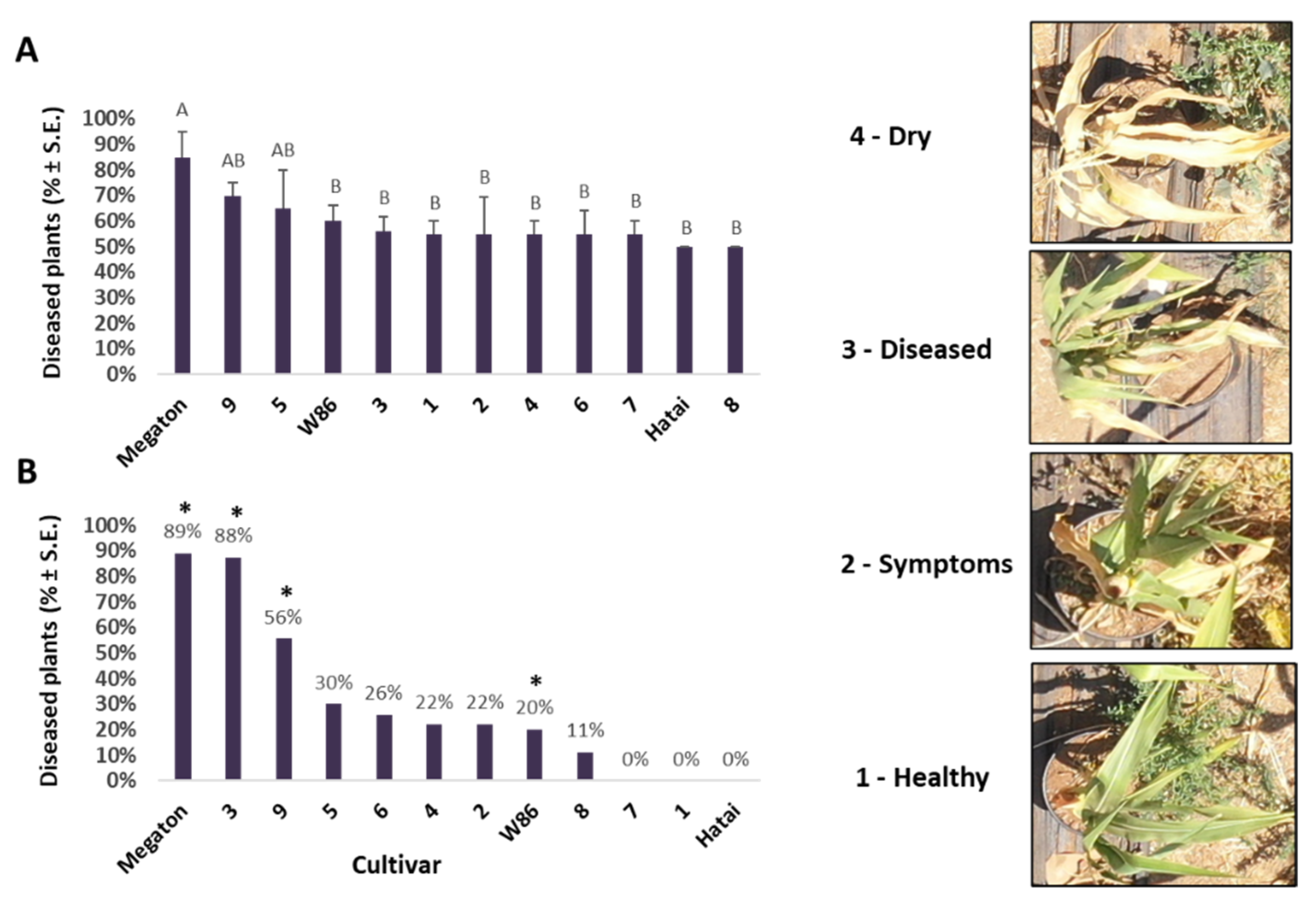
3.2.1. Aboveground Appearance Rate Estimate
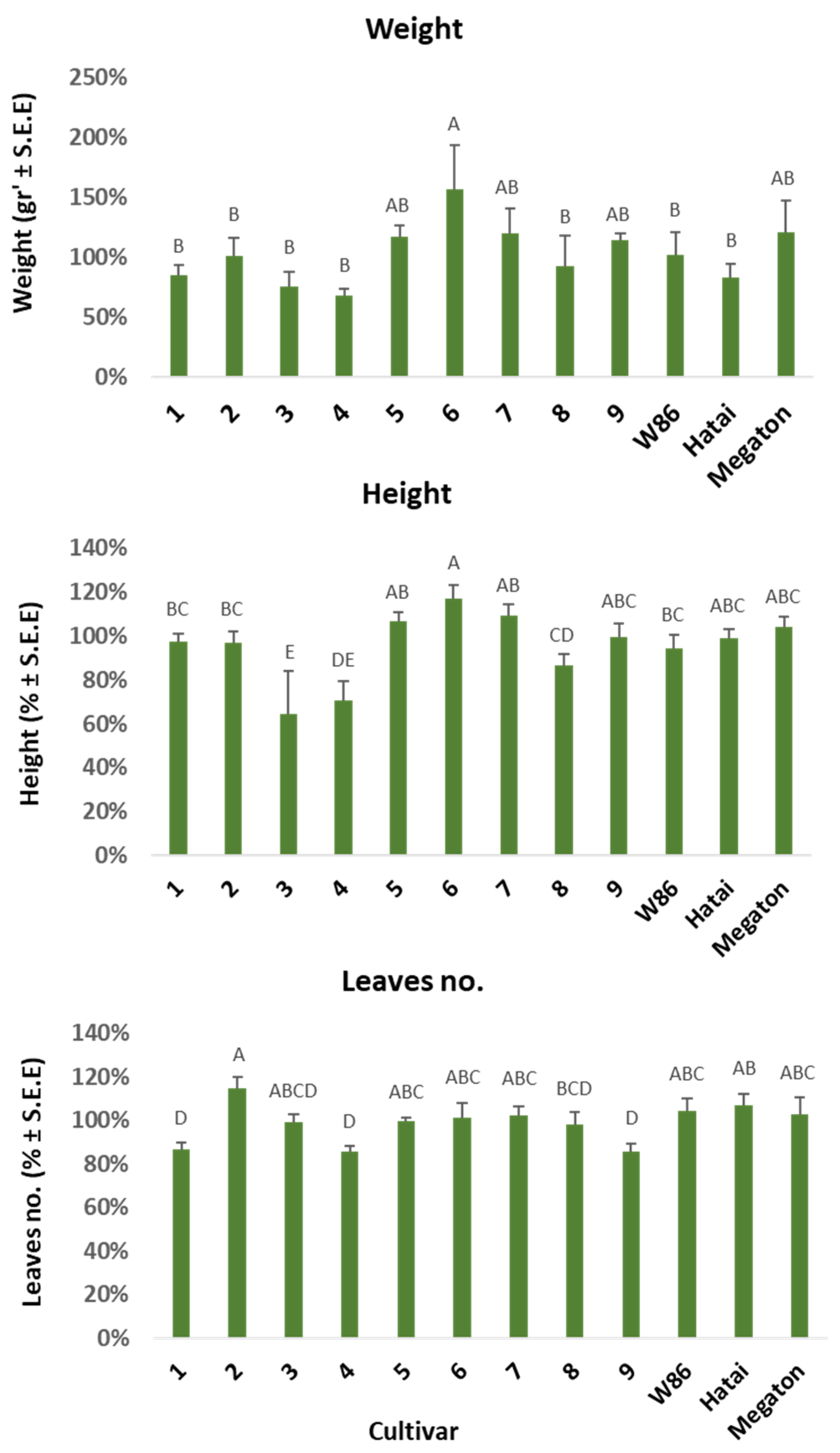
3.2.2. Growth Indices on Day 50 of Growth
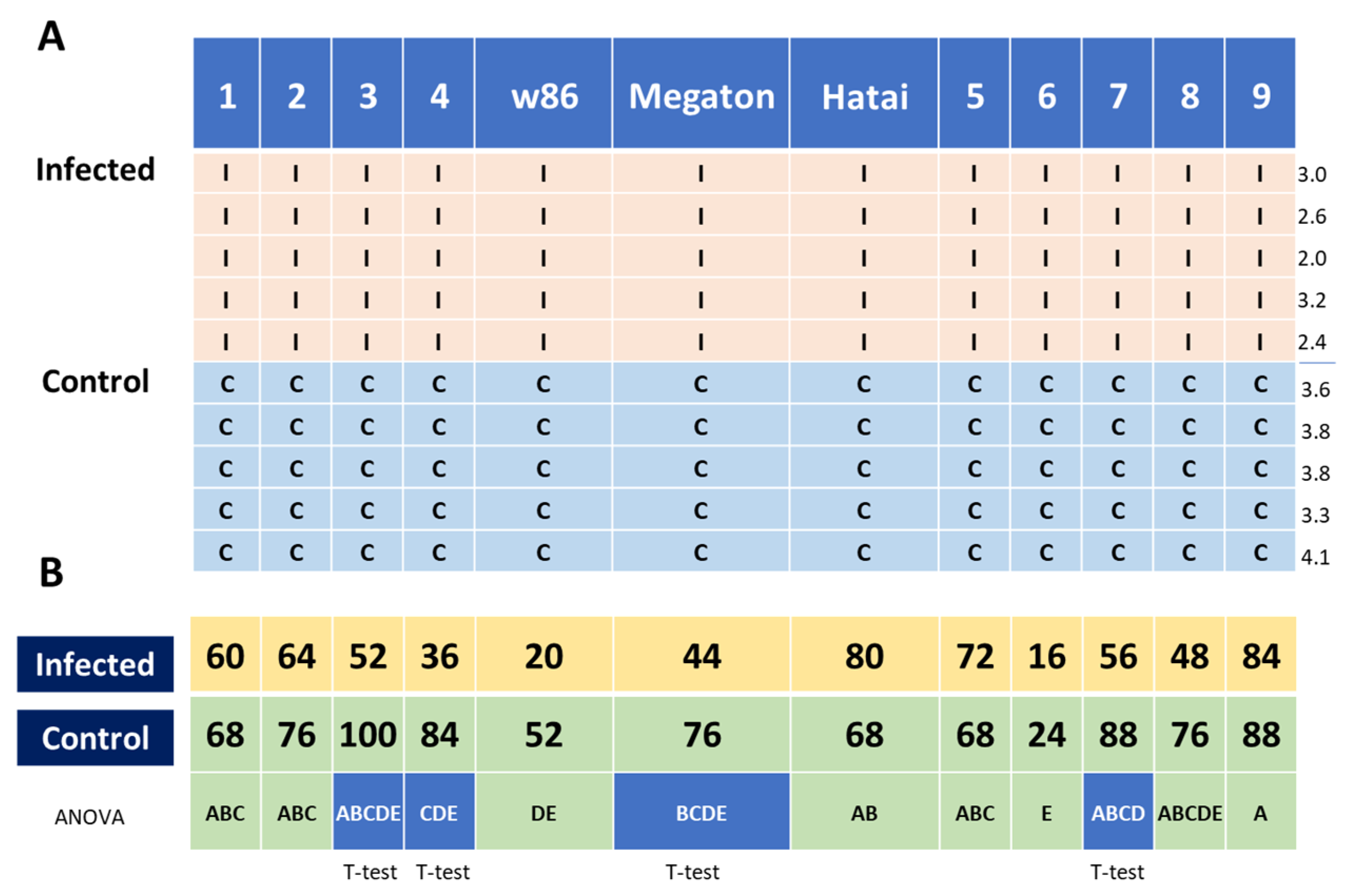
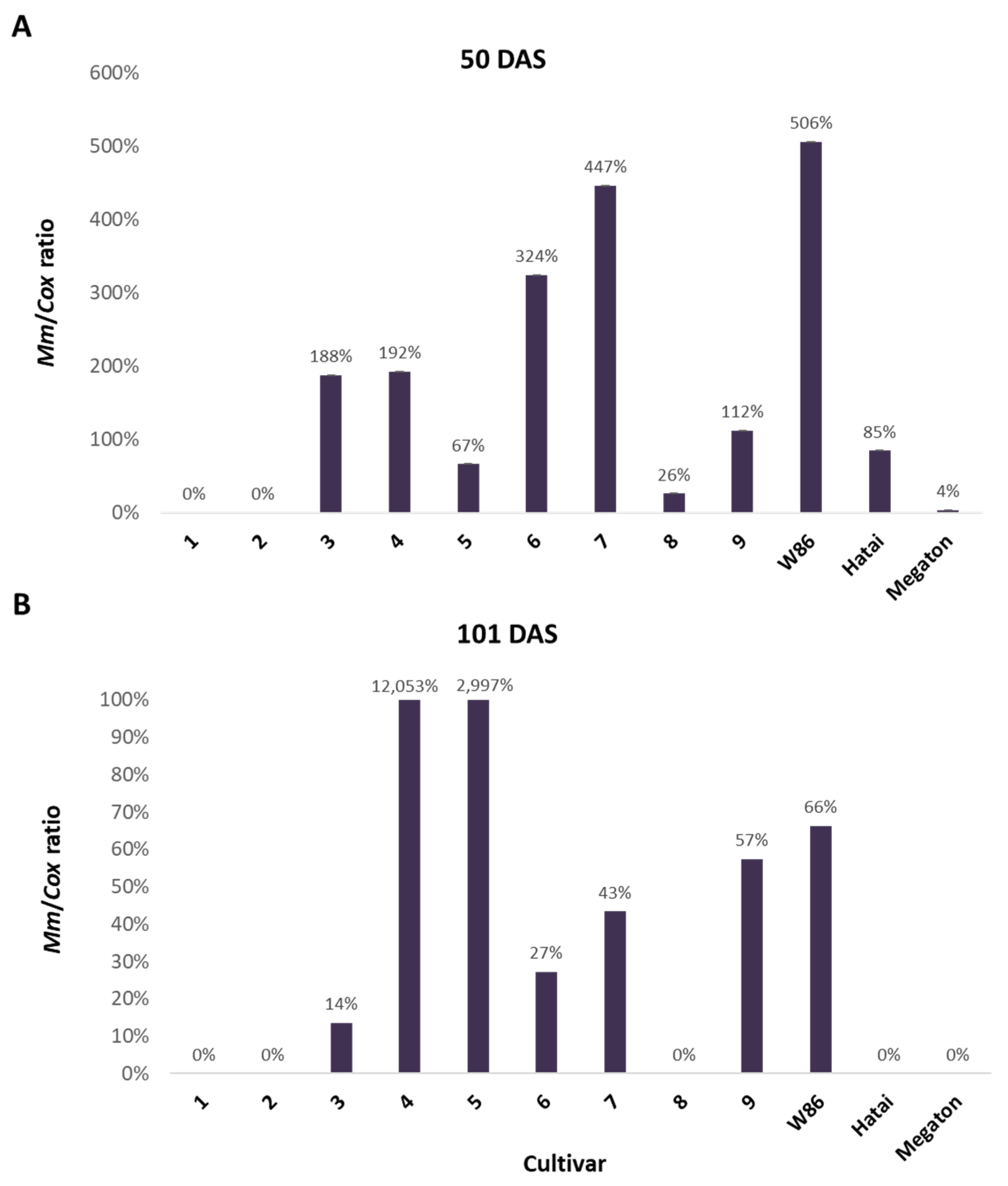
3.2.3. Infection Level throughout the Growing Season
3.2.4. Growth Indices and Dehydration Level at the Harvest
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klaubauf, S.; Tharreau, D.; Fournier, E.; Groenewald, J.Z.; Crous, P.W.; de Vries, R.P.; Lebrun, M.H. Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Stud. Mycol. 2014, 79, 85–120. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Restrepo, M.; Groenewald, J.; Elliott, M.; Canning, G.; McMillan, V.; Crous, P.W. Take-all or nothing. Stud. Mycol. 2016, 83, 19–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.-B.; Hyde, K.D.; Yang, E.-F.; Kakumyan, P.; Bahkali, A.H.; Elgorban, A.M.; Karunarathna, S.C.; Phookamsak, R.; Lumyong, S. Morphological and phylogenetic appraisal of Ophioceras (Ophioceraceae, Magnaporthales). PLoS ONE 2021, 16, e0253853. [Google Scholar] [CrossRef] [PubMed]
- Gams, W. Phialophora and some similar morphologically little-differentiated anamorphs of divergent ascomycetes. Stud. Mycol. 2000, 45, 187–200. [Google Scholar]
- Ward, E.; Bateman, G.L. Comparison of Gaeumannomyces- and Phialophora-like fungal pathogens from maize and other plants using DNA methods. New Phytol. 1999, 141, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Saleh, A.A.; Leslie, J.F. Cephalosporium maydis is a distinct species in the Gaeumannomyces-Harpophora species complex. Mycologia 2004, 96, 1294–1305. [Google Scholar] [CrossRef]
- Samra, A.S.; Sabet, K.A.; Hingorani, M.K. Late wilt disease of maize caused by Cephalosporium maydis. Phytopathology 1963, 53, 402–406. [Google Scholar]
- Degani, O.; Chen, A.; Dor, S.; Orlov-Levin, V.; Jacob, M.; Shoshani, G.; Rabinovitz, O. Remote evaluation of maize cultivars susceptibility to late wilt disease caused by Magnaporthiopsis maydis. J. Plant Pathol. 2022, 104, 509–525. [Google Scholar] [CrossRef]
- Elshahawy, I.E.; Khattab, A.E.-N.A. Endophyte Chaetomium globosum improves the growth of maize plants and induces their resistance to late wilt disease. J. Plant Dis. Prot. 2022, 129, 1125–1144. [Google Scholar] [CrossRef]
- El-Shenawy, A.; Hassan, M.; Abo El-Haress, S.; Abd-Elaziz, M. Assessment of combining ability in some newly maize inbred lines for grain yield and late wilt resistance. J. Plant Prod. 2022, 13, 45–48. [Google Scholar] [CrossRef]
- El-Shabrawy, E. Use silica nanoparticles in controlling late wilt disease in maize caused by Harpophora maydis. Egypt. J. of Appl. Sci. 2021, 36, 1–19. [Google Scholar] [CrossRef]
- Agag, S.H.; Sabry, A.M.; EL-Samman, M.G.; Mostafa, M.H. Pathological and molecular characterization of Magnaporthiopsis maydis isolates causing late wilt in maize. Egypt. J. Phytopathol. 2021, 49, 1–9. [Google Scholar] [CrossRef]
- Kamara, M.M.; Ghazy, N.A.; Mansour, E.; Elsharkawy, M.M.; Kheir, A.M.S.; Ibrahim, K.M. Molecular Genetic Diversity and Line × Tester Analysis for Resistance to Late Wilt Disease and Grain Yield in Maize. Agronomy 2021, 11, 898. [Google Scholar] [CrossRef]
- Rakesh, B.; Gangappa, E.; Gandhi, S.; Gowda, R.P.V.; Swamy, S.D.; Ramesh, S.; Hemareddy, H.B. Mapping genomic regions controlling resistance to late wilt disease caused by Harpophora maydis in maize (Zea mays L.). Euphytica 2022, 218, 101. [Google Scholar] [CrossRef]
- Gazala, P.; Gangappa, E.; Ramesh, S.; Swamy, D. Comparative Breeding potential of two crosses for response to late wilt disease (LWD) in maize (Zea mays L.). Genet. Resour. Crop Evol. 2021, 68, 1751–1757. [Google Scholar] [CrossRef]
- Sunitha, N.; Gangappa, E.; Gowda, R.V.; Ramesh, S.; Biradar, S.; Swamy, D.; Hemareddy, H. Assessment of Impact of Late Wilt Caused by Harpophora maydis (Samra, Sabet and Hing) on Grain Yield and its Attributing Traits in Maize (Zea mays L.). Mysore J. Agric. Sci. 2020, 54, 30–36. [Google Scholar]
- Biradar, M.; Gangappa, E.; Ramesh, S.; Sowjanya, P.R.; Sunitha, N.; Parveen, G.; Sowmya, H.; Suma, K. Association Between GCA and Per Se Performance of Parents and Hybrids for Grain Yield, its Attributing Traits and Late Wilt Disease (Harpophora maydis) Resistance in Maize (Zea mays L.). Int. J. Curr. Microbiol. App. Sci 2020, 9, 2560–2570. [Google Scholar] [CrossRef]
- Tej, R.; Rodríguez-Mallol, C.; Rodríguez-Arcos, R.; Karray-Bouraoui, N.; Molinero-Ruiz, L. Inhibitory effect of Lycium europaeum extracts on phytopathogenic soil-borne fungi and the reduction of late wilt in maize. Eur. J. Plant Pathol. 2018, 152, 249–265. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Bustos, C.M.; Testi, L.; García-Carneros, A.B.; Molinero-Ruiz, L. Geographic distribution and aggressiveness of Harpophora maydis in the Iberian peninsula, and thermal detection of maize late wilt. Eur. J. Plant Pathol. 2015, 144, 383–397. [Google Scholar] [CrossRef]
- García-Carneros, A.; Girón, I.; Molinero-Ruiz, L. Aggressiveness of Cephalosporium maydis causing late wilt of maize in Spain. Commun. Agric. Appl. Biol. Sci. 2011, 77, 173–179. [Google Scholar]
- Molinero-Ruiz, M.L.; Melero-Vara, J.M.; Mateos, A. Cephalosporium maydis, the cause of late wilt in maize, a pathogen new to Portugal and Spain. Plant Dis. 2010, 94, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patanita, M.; Campos, M.D.; Félix, M.d.R.; Carvalho, M.; Brito, I. EEffect of tillage system and cover crop on maize mycorrhization and presence of Magnaporthiopsis maydis. Biology 2020, 9, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degani, O.; Movshowitz, D.; Dor, S.; Meerson, A.; Goldblat, Y.; Rabinovitz, O. Evaluating Azoxystrobin seed coating against maize late wilt disease using a sensitive qPCR-based method. Plant Dis. 2019, 103, 238–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samra, A.; Sabet, K.; Kamel, M. Further studies on the effect of field conditions and cultural practices on infection with stalk-rot complex of maize. Arab Re-public of Egypt, Min. of Agriculture. Plant Prot. Dept. Bull 1971. no. 2. Available online: http://library.wur.nl/webquery/clc/385177 (accessed on 16 November 2022).
- Payak, M.; Sharma, R. Research on Diseases of Maize; Indian Council of Agricultural Research: New Delhi, India, 1978; p. 228. [Google Scholar]
- Degani, O. A review: Late wilt of maize—The pathogen, the disease, current status and future perspective. J. Fungi 2021, 7, 989. [Google Scholar] [CrossRef]
- Bergstrom, G.; Leslie, F.J.; Huber, D.; Lipps, P.; Warren, H.; Esker, P.; Grau, C.; Botratynski, T.; Bulluck, R.; Floyd, J.; et al. Recovery Plan for Late Wilt of Corn Caused by Harpophora Maydis Syn. Cephalosporium Maydis; National Plant Disease Recovery System (NPDRS): Washington, DC, USA, 2008; p. 24. [Google Scholar]
- Johal, L.; Huber, D.M.; Martyn, R. Late wilt of corn (maize) pathway analysis: Intentional introduction of Cephalosporium maydis. In Pathways Analysis for the Introduction to the U.S. of Plant Pathogens of Economic Importance; Purdue University: West Lafayette, IN, USA, 2004. [Google Scholar]
- Michail, S.H.; Abou-Elseoud, M.S.; Nour Eldin, M.S. Seed health testing of corn for Cephalosporium maydis. Acta Phytopathol. Entomol. Hung. 1999, 34, 35–42. [Google Scholar]
- Sabet, K.A.; Samra, A.S.; Mansour, I.M. Saprophytic behaviour of Cephalosporium maydis and C. acremonium. Ann. Appl. Biol. 1970, 66, 265–271. [Google Scholar] [CrossRef]
- Sahab, A.F.; Osman, A.R.; Soleman, N.K.; Mikhail, M.S. Studies on root-rot of lupin in Egypt and its control. Egypt. J. Phytopathol. 1985, 17, 23–35. [Google Scholar]
- Sabet, K.; Samra, A.; Mansour, I. Interaction between Fusarium oxysporum f. vasinfectum and Cephalosporium maydis on cotton and maize. Ann. Appl. Biol. 1966, 58, 93–101. [Google Scholar] [CrossRef]
- Elshahawy, I.E.; Abd El-Wahed, M.S. Suppression of Cephalosporium maydis by the resistance inducer beta-sitosterol. Eur. J. Plant Pathol. 2022, 163, 673–693. [Google Scholar] [CrossRef]
- Hassan, E.O.; Shoala, T.; Attia, A.M.F.; Badr, O.A.M.; Mahmoud, S.Y.M.; Farrag, E.S.H.; EL-Fiki, I.A.I. Chitosan and nano-chitosan for management of Harpophora maydis: Approaches for investigating antifungal activity, pathogenicity, maize-resistant lines, and molecular diagnosis of plant infection. J. Fungi 2022, 8, 509. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Kader, M.; Khalil, M.; El-Mougy, N. Efficacy of fungicide alternatives against late wilt disease of maize and their influence on plant morphogenesis and yield characters. Hell. Plant Prot. J. 2022, 15, 57–71. [Google Scholar] [CrossRef]
- Doleib, N.M.; Farfour, S.A.; Al-shakankery, F.M.; Ammar, M.; Hamouda, R.A. Antifungal activates of cyanobacteria and some marine algae against Cephalosporium maydis, the cause of maize late wilt disease in vitro. Biosci. Res. 2021, 18, 536–543. [Google Scholar]
- Degani, O. Control strategies to cope with late wilt of maize. Pathogens 2022, 11, 13. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S.; Chen, A.; Orlov-Levin, V.; Stolov-Yosef, A.; Regev, D.; Rabinovitz, O. Molecular Tracking and Remote Sensing to Evaluate New Chemical Treatments Against the Maize Late Wilt Disease Causal Agent, Magnaporthiopsis maydis. J. Fungi 2020, 6, 54. [Google Scholar] [CrossRef]
- Avila-Adame, C.; Koller, W. Characterization of spontaneous mutants of Magnaporthe grisea expressing stable resistance to the Qo-inhibiting fungicide azoxystrobin. Curr. Genet. 2003, 42, 332–338. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining biocontrol agents with chemical fungicides for integrated plant fungal disease control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef]
- Abdel-Hamid, M.S.; Abdel-Momeim, M.F.; El-Shafey, H.A.; El-Deeb, S.T. Biological control of late-wilt disease of maize caused by Cephalosporium maydis. Agric. Res. Rev. 1981, 59, 253–260. [Google Scholar]
- El-Assiuty, E.M.; El-Hamahmy, A.A.; El-Sharkawy, A.Y. Bacillus subtilis, Pseudomonas fluorescens and Verticillium tricorpus as biological agents against late-wilt of maize. Egypt. J. Appl. Sci. 1991, 6, 8245–8829. [Google Scholar]
- El-Mehalowy, A.A.; Hassanein, N.M.; Khater, H.M.; Daram El-Din, E.A.; Youssef, Y.A. Influence of maize root colonization by rhizosphere actinomycetes and yeast fungi on plant growth and on the biological control of late wilt disease. Inter. J. Agric. Biol. 2004, 6, 599–605. [Google Scholar]
- Ferreira, F.V.; Musumeci, M.A. Trichoderma as biological control agent: Scope and prospects to improve efficacy. World J. Microbiol. Biotechnol. 2021, 37, 90. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, M.; El-Moghazy, S.; Mehesen, A.A. Biological control of maize late wilt disease caused by Cephalosporium maydis. J. Agric. Res. Kafrelsheikh Univ. 2009, 35, 1–19. [Google Scholar]
- El-Gazzar, N.; El-Bakery, A.M.; Ata, A.A. Influence of some bioagents and chitosan nanoparticles on controlling maize late wilt and improving plants characteristics. Egypt. J. Phytopathol. 2018, 46, 243–264. [Google Scholar] [CrossRef]
- Hamza, A.M.; El-Kot, G.; El-Moghazy, S. Non-traditional methods for controlling maize late wilt disease caused by Cephalosporium maydis. Egypt. J. Biol. Pest Control. 2013, 23, 87–93. [Google Scholar]
- Mosa, H.; Motawei, A.; El-Aal, A.A. Nitrogen fertilization influence on combining ability for grain yield and resistance to late wilt disease in maize. J. Agric. Res. Kafrelsheikh Univ. 2010, 36, 278–291. [Google Scholar]
- El-Moghazy, S.; Shalaby, M. Effects of some aqueous plant extracts and sulphur compounds on the control of maize late wilt disease caused by Cephalosporium maydis. J. Agric. Res. Tanta Univ 2006, 32, 758–775. [Google Scholar]
- Elshahawy, I.E.; El-Sayed, A.E.-K.B. Maximizing the efficacy of Trichoderma to control Cephalosporium maydis, causing maize late wilt disease, using freshwater microalgae extracts. Egypt. J. Biol. Pest Control. 2018, 28, 48. [Google Scholar] [CrossRef] [Green Version]
- Sunitha, N.; Gangappa, E.; Gowda, R.V.; Ramesh, S.; Biradar, S.; Swamy, D.; Hemareddy, H. Discovery of genomic regions associated with resistance to late wilt disease caused by Harpophora Maydis (Samra, Sabet and Hing) in maize (Zea mays L.). J. Appl. Genet. 2021, 63, 185–197. [Google Scholar] [CrossRef]
- Kumar, K.A.; Gangappa, E.; Ramesh, S.; Swamy, D. Identification of high yielding inbred lines resistant to late wilt disease caused by Harpophora maydis in maize. Electron. J. Plant Breed. 2021, 12, 151–158. [Google Scholar] [CrossRef]
- Sunitha, N.; Gangappa, E.; Gowda, R.V.; Ramesh, S.; Swamy, S.D.; Hemareddy, H. Effectiveness of one cycle of phenotype-based backcross breeding for resistance to late wilt disease in maize (Zea mays L.). Mysore J. Agric. Sci. 2020, 54, 47–50. [Google Scholar]
- Farahat, G.A.; Barakat, E.H.; El-Bana, M. Effects of late wilt disease on infection development of ear rot disease, phenolic compounds, trypsin and α-amylase inhibitors of some maize hybrids grains and quality characteristics of fortified cookies. Middle East J. Agric. Res. 2020, 9, 515–532. [Google Scholar]
- Zeller, K.A.; Ismael, A.M.; El-Assiuty, E.M.; Fahmy, Z.M.; Bekheet, F.M.; Leslie, J.F. Relative competitiveness and virulence of four clonal lineages of Cephalosporium maydis from Egypt Toward greenhouse-grown maize. Plant Dis. 2002, 86, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Shafey, H.A.; El-Shorbagy, F.A.; Khalil, I.I.; El-Assiuty, E.M. Additional sources of resistance to the late-wilt disease of maize caused by Cephalosporium maydis. Agric. Res. Rev. Egypt. 1988, 66, 221–230. [Google Scholar]
- Ghazy, N.; El-Gremi, S.; Belal, E.-S. Chemical and Histological Differences of Corn (Zea mays L.) Responsive to Harpophora maydis Infection. Environ. Biodivers. Soil Secur. 2017, 1, 3–7. [Google Scholar] [CrossRef]
- Degani, O.; Regev, D.; Dor, S.; Rabinovitz, O. Soil Bioassay for Detecting Magnaporthiopsis maydis Infestation Using a Hyper Susceptible Maize Hybrid. J. Fungi 2020, 6, 107. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S.; Movshovitz, D.; Rabinovitz, O. Methods for studying Magnaporthiopsis maydis, the maize late wilt causal agent. Agronomy 2019, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [Green Version]
- El-Gremi, S.M.A.; Belal, E.B.A.; Ghazy, N.A. Cephalosporium maydis as affected by maize root exudates and role of the fungal metabolites in pathogenesis. J. Agric. Sci. Mansoura Univ. 2007, 32, 7605–7615. [Google Scholar] [CrossRef]
- Galal, O.A.; Aboulila, A.A.; Motawei, A.; Galal, A. Biochemical and molecular diversity and their relationship to late wilt disease resistance in yellow maize inbred lines. Egypt. J. Genet. Cytol. 2018, 47, 203–227. [Google Scholar]
- Degani, O.; Drori, R.; Goldblat, Y. Plant growth hormones suppress the development of Harpophora maydis, the cause of late wilt in maize. Physiol. Mol. Biol. Plants 2015, 21, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Shofman, G.; Bahouth, M.; Degani, O. Aggressive strains of the late wilt fungus of corn exist in Israel in mixed populations and can specialize in disrupting growth or plant health. Fungal Biol. 2022, 126, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elrazik, A.; Darweish, F.A.; Rushdi, M.H. The role of certain oxidative enzymes, catalase, and beta-glucosidase on virulence of Cephalosporium maydis. Zent. Bakteriol. Parasitenkd. Infekt. Und Hyg. 1978, 133, 240–244. [Google Scholar] [CrossRef]
- Degani, O.; Rabinovitz, O.; Becher, P.; Gordani, A.; Chen, A. Trichoderma longibrachiatum and Trichoderma asperellum confer growth promotion and protection against late wilt disease in the field. J. Fungi 2021, 7, 444. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Bustos, C.; López-Bernal, A.; Testi, L.; Molinero-Ruiz, L. Environmental and irrigation conditions can mask the effect of Magnaporthiopsis maydis on growth and productivity of maize. Plant Pathol. 2019, 68, 1555–1564. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Cultivar | Type | Seed Company | Supply Company | Degree of LWD Sensitivity a |
|---|---|---|---|---|---|
| 1 | KXB9571 | Fodder | KWS, Einbeck, Lower Saxony, Germany | CTS Group, Tel Aviv, Israel | N/A |
| 2 | KAC0572 | Fodder | KWS, Einbeck, Lower Saxony, Germany | CTS Group, Tel Aviv, Israel | N/A |
| 3 | KES Elektro | Fodder | KWS, Einbeck, Lower Saxony, Germany | CTS Group, Tel Aviv, Israel | N/A |
| 4 | Leonidos | Fodder | KWS, Einbeck, Lower Saxony, Germany | CTS Group, Tel Aviv, Israel | N/A |
| 5 | Calumet | Fodder | KWS, Einbeck, Lower Saxony, Germany | CTS Group, Tel Aviv, Israel | N/A |
| 6 | Laurca | Fodder | KWS, Einbeck, Lower Saxony, Germany | CTS Group, Tel Aviv, Israel | Resistant [8] |
| 7 | Colossus | Fodder | Semillas Fitó, Barcelona, Spain | Tarsis Inc., Petach Tikva, Israel | Sensitive [8,26,59] |
| 8 | Hatai | Fodder | Semillas Fitó, Barcelona, Spain | Tarsis Inc., Petach Tikva, Israel | Highly resistant [8,58] |
| 9 | 32D99 | Fodder | Pioneer Hi-Bred International, Inc. Johnston, Iowa, USA | Gadot Agro, Kidron, Israel | Highly resistant [8,23] |
| 10 | P32W86 (W86) | Fodder | Pioneer Hi-Bred International, Inc. Johnston, Iowa, USA | Gadot Agro, Kidron, Israel | Sensitive [23] |
| 11 | Hatai | Fodder | Semillas Fitó, Barcelona, Spain | Tarsis Inc., Petach Tikva, Israel | Highly resistant [8,58] |
| 12 | Megaton | Sweet | Limagrain, Saint-Beauzire, Puy-de-Dôme, France | Hazera Seeds Ltd., Berurim MP Shikmim, Israel | Hypersensitive [58] |
| 13 | Prelude | Sweet | SRS—snowy river seeds | Green 2000, Bitan Aharon, Israel | Hypersensitive [23,58] |
| Cultivar | DiseasedPlants | Height | Plants’ Wet Weight | Leaves No. | Cobs Fresh Weight |
|---|---|---|---|---|---|
| 1 | 48% | 99% | 114% | 90% | 76% |
| 2 | 52% | 95% | 117% | 94% | 91% |
| 3 | 50% | 96% | 89% | 102% | 97% |
| 4 | 32% | 93% | 125% | 95% | 159% |
| 5 | 60% | 99% | 86% | 87% | 88% |
| 6 | 48% | 91% | 93% | 82% | 115% |
| 7 | 44% | 98% | 135% | 100% | 102% |
| 8 | 48% | 98% | 106% | 93% | 138% |
| 9 | 44% | 99% | 83% | 110% | 94% |
| w86 | 48% | 90% | 87% | 93% | 83% |
| Hatai | 44% | 94% | 124% | 108% | 128% |
| Megaton | 68% | 91% | 89% | 80% | 68% |
| Seed Assay | Pot Assay | Total Rank | |||||
|---|---|---|---|---|---|---|---|
| Seed germination 6 days | Sprouts emergence 6 DAS | qPCR 50 DAS | qPCR 101 DAS | Air symptoms 92 DAS | Ground symptoms 101 DAS | Growth indexes 101 DAS | Estimated resistance order |
| Hatai | Hatai | 1 | 1 | Hatai | 4 | 7 | 7 |
| 1 | 5 | 2 | 2 | 1 | Hatai | Hatai | Hatai |
| 5 | 9 | 8 | 8 | 7 | 7 | 2 | 4 |
| 7 | 1 | 5 | Hatai | 8 | 9 | 4 | 2 |
| 3 | 2 | Hatai | 3 | w86 | 1 | 8 | 1 |
| 4 | w86 | 9 | 6 | 2 | 6 | 9 | 8 |
| 9 | 8 | 3 | 7 | 4 | 8 | 1 | 9 |
| 2 | 6 | 4 | 9 | 6 | w86 | 5 | 3 |
| 8 | Meg | 6 | w86 | 5 | 3 | 3 | 5 |
| w86 | 7 | 7 | 5 | 9 | 2 | 6 | 6 |
| 6 | 3 | w86 | 4 | 3 | 5 | w86 | w86 |
| Meg | 4 | Meg | Meg | Meg | Meg | Meg | Meg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Degani, O.; Yifa, R.; Gordani, A.; Becher, P.; Chen, A. Cultivars Resistance Assay for Maize Late Wilt Disease. Biology 2022, 11, 1854. https://doi.org/10.3390/biology11121854
Degani O, Yifa R, Gordani A, Becher P, Chen A. Cultivars Resistance Assay for Maize Late Wilt Disease. Biology. 2022; 11(12):1854. https://doi.org/10.3390/biology11121854
Chicago/Turabian StyleDegani, Ofir, Ran Yifa, Asaf Gordani, Paz Becher, and Assaf Chen. 2022. "Cultivars Resistance Assay for Maize Late Wilt Disease" Biology 11, no. 12: 1854. https://doi.org/10.3390/biology11121854