
Paramecium bursaria as a Potential Tool for Evaluation of Microplastics Toxicity
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Microplastic Characterizations
2.3. Toxic Effects of MPs Exposure to P. bursaria
2.4. Flow Cytometric and Fluorescent Microscopic Analysis of MPs Uptake and Accumulation in P. bursaria
2.5. Effects of MPs Exposure on P. bursaria Locomotion Activity, Avoidance Behaviors, Prey Pursuit, and Predator Evasion Ability
2.6. Examination of Oxidative Stress Status of P. bursaria following MPs Exposure
2.7. Statistics
3. Results
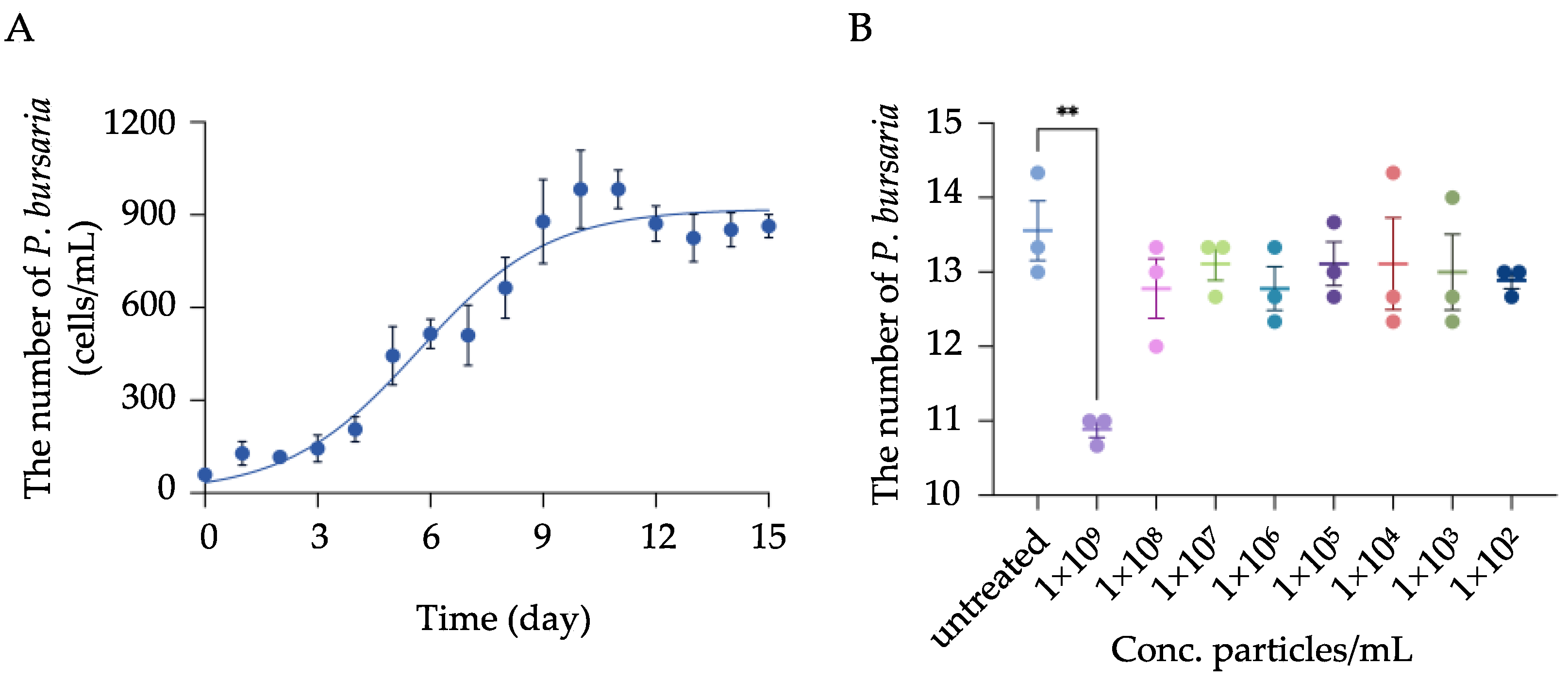
3.1. Physicochemical Characterization of MPs and Determination of Toxic Concentration of MPs on P. bursaria Growth
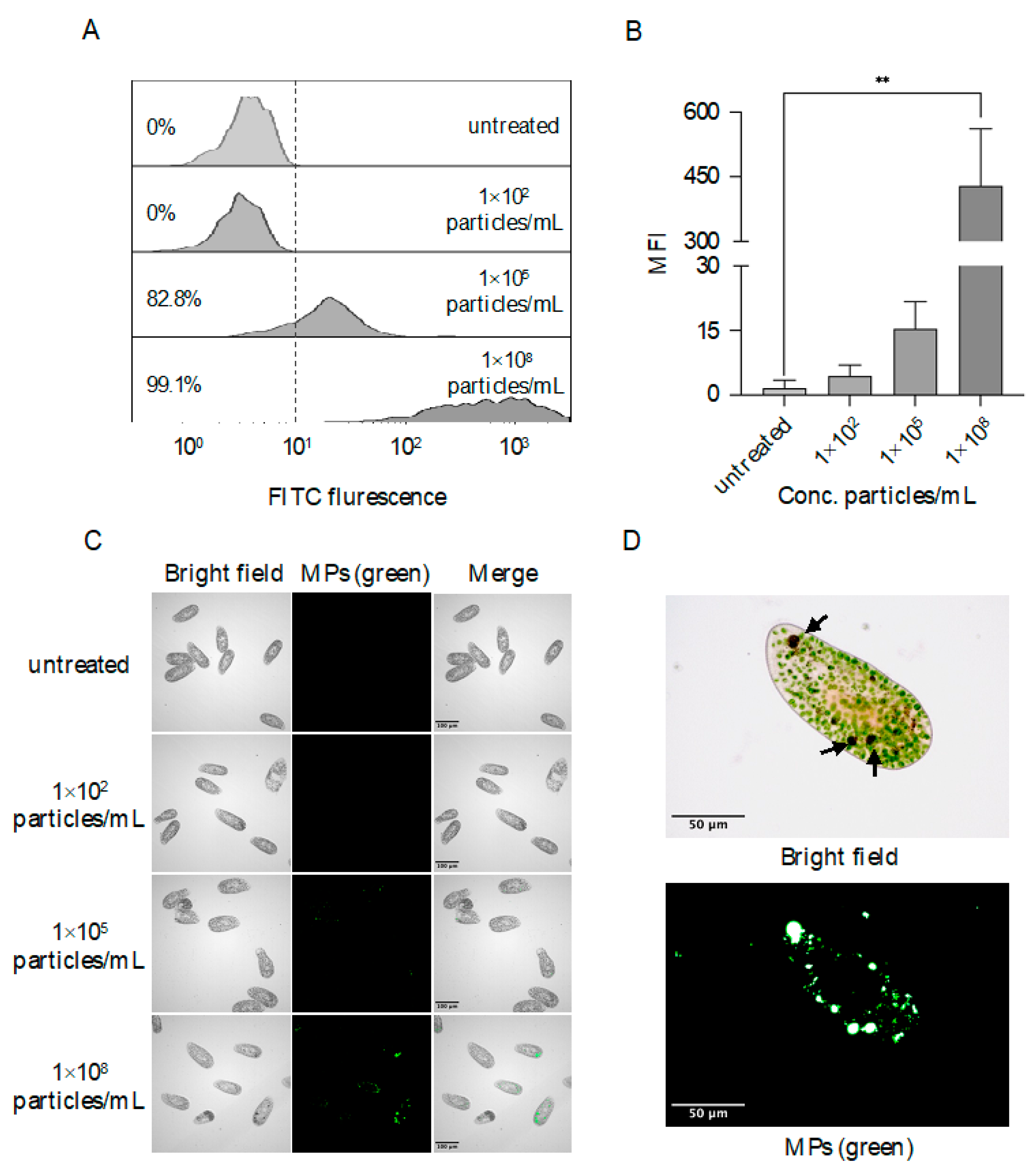
3.2. Accumulation of MPs in Food Vacuoles
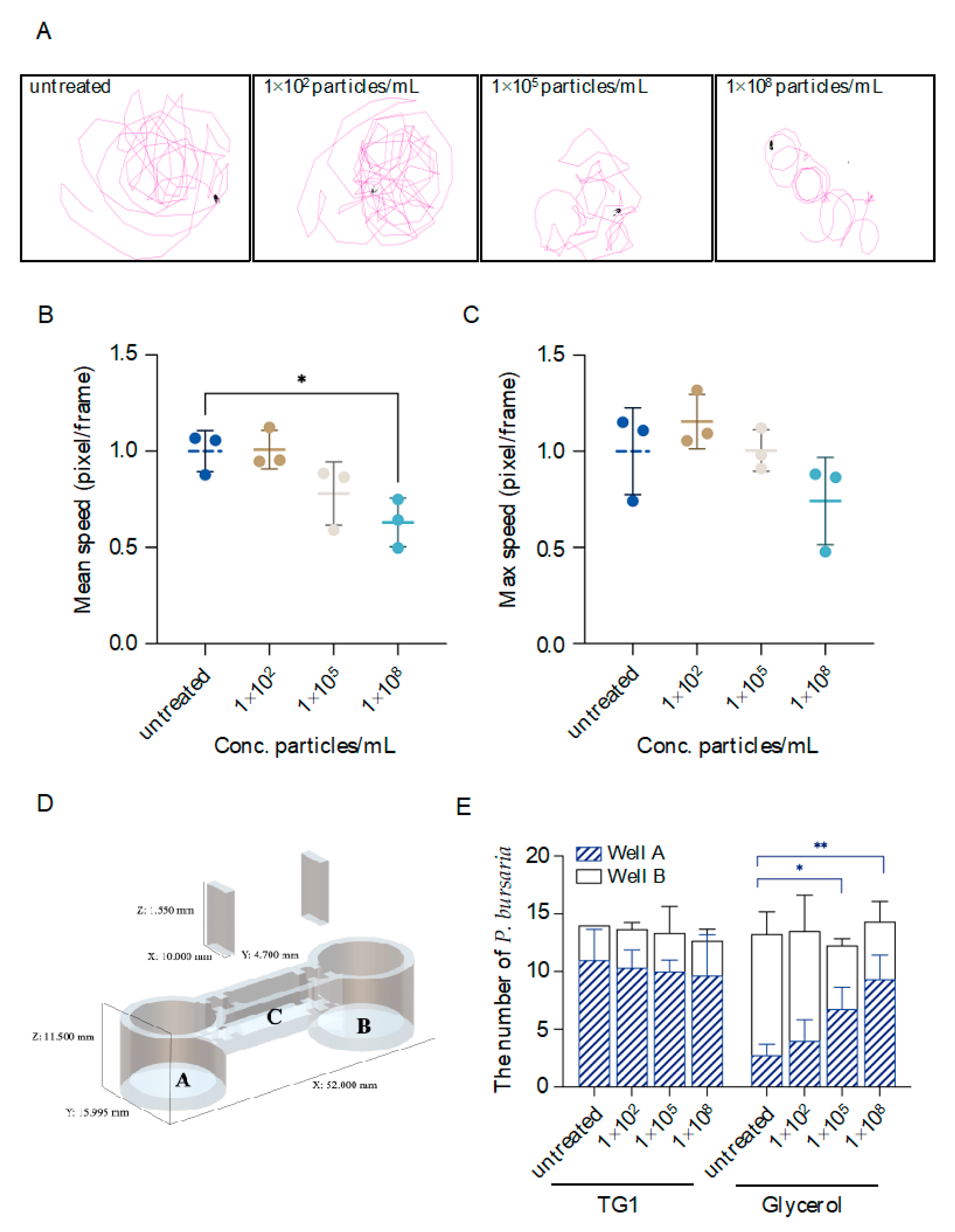
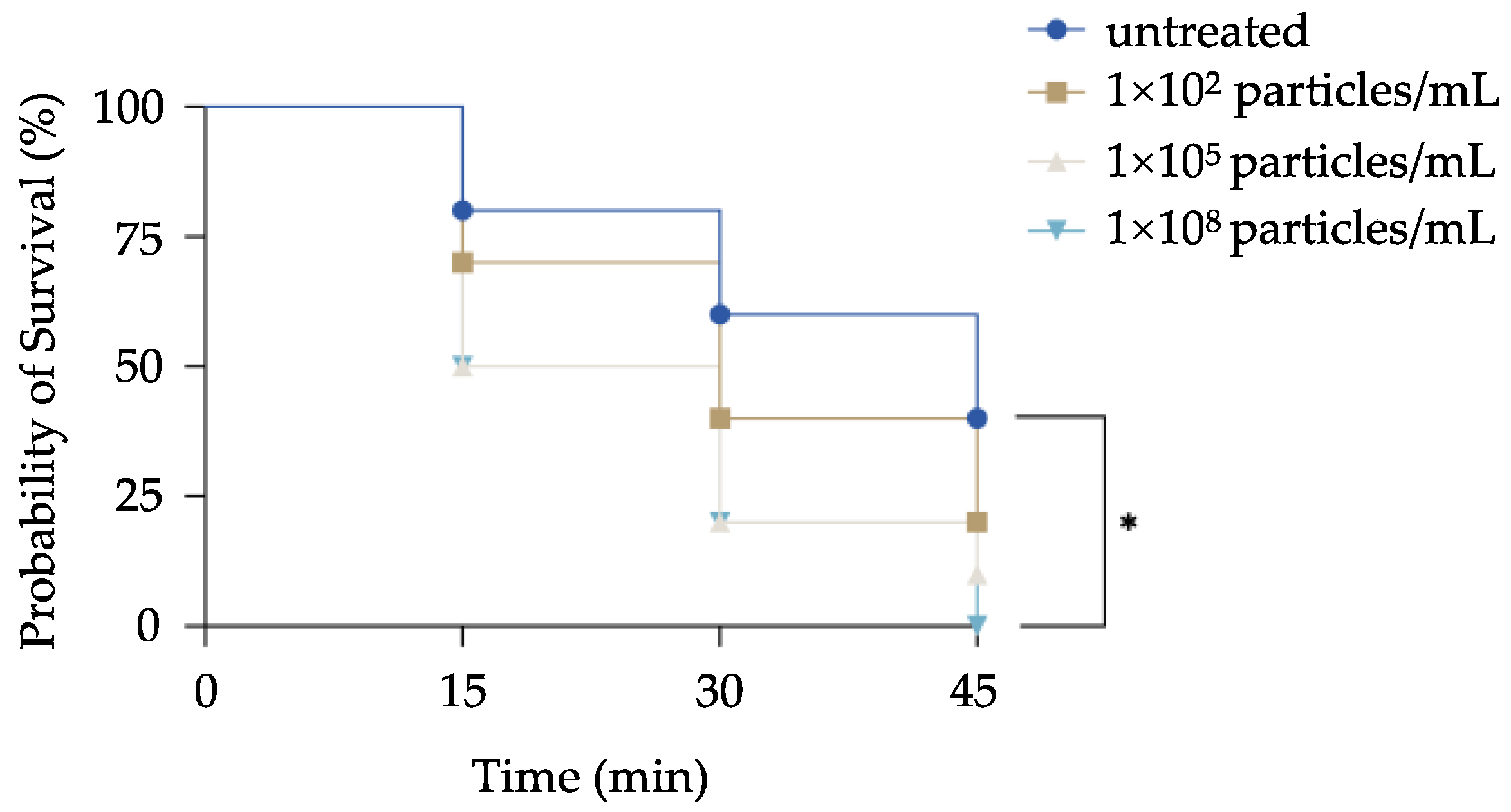
3.3. MPs-Exposed P. bursaria Damage on Locomotion Activity, Prey Pursuit, and Avoidance Behaviors
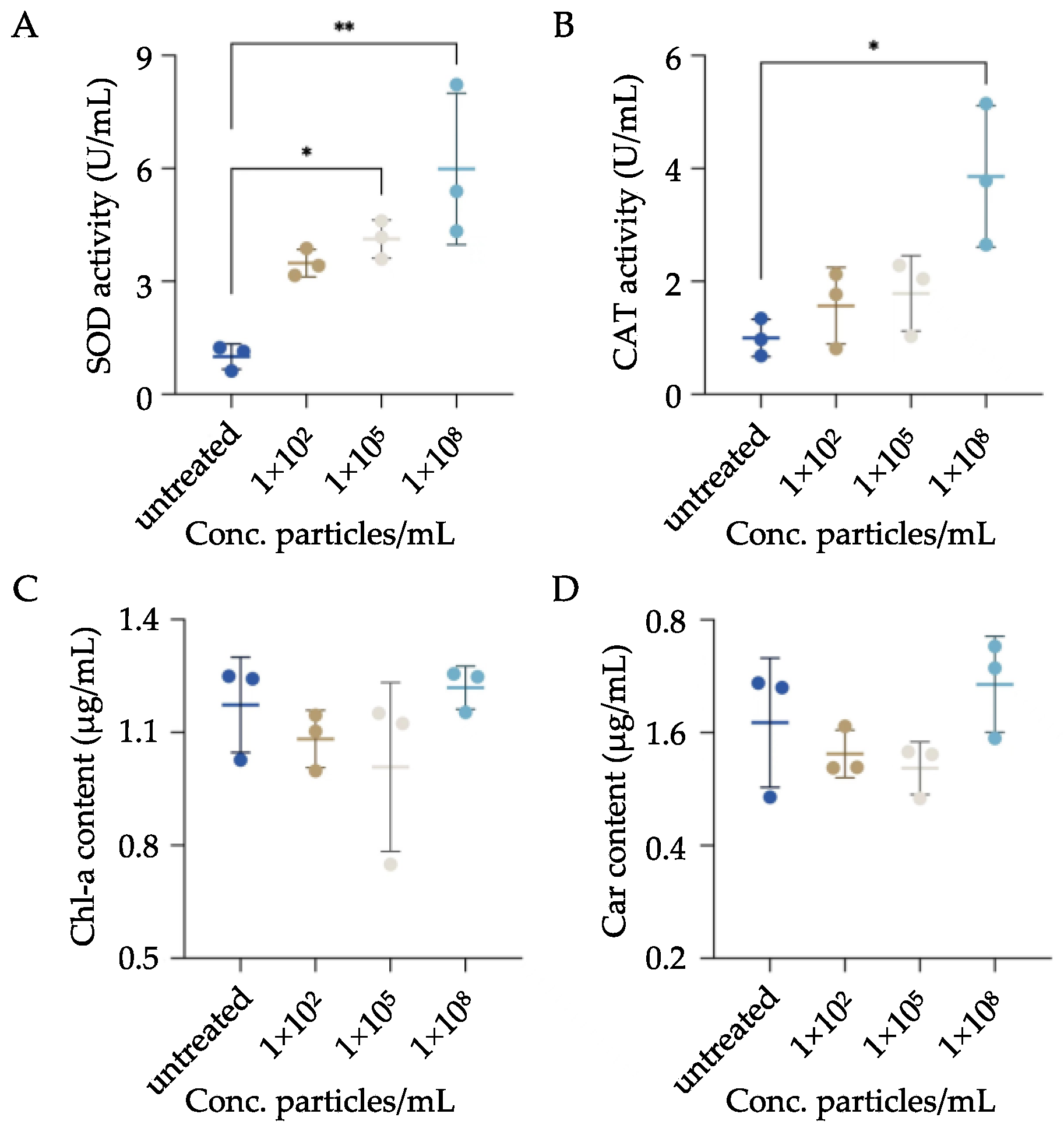
3.4. Impacts of MPs Exposure on the Oxidative Stress Status and Endosymbiont of P. bursaria
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Provencher, J.F.; Ammendolia, J.; Rochman, C.M.; Mallory, M.L. Assessing Plastic Debris in Aquatic Food Webs: What We Know and Don’t Know about Uptake and Trophic Transfer. Environ. Rev. 2019, 27, 304–317. [Google Scholar] [CrossRef] [Green Version]
- Gall, S.C.; Thompson, R.C. The Impact of Debris on Marine Life. Mar. Pollut. Bull. 2015, 92, 170–179. [Google Scholar] [CrossRef]
- Law, K.L.; Thompson, R.C. Microplastics in the Seas. Science 2014, 345, 144–145. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Redondo-Hasselerharm, P.E.; Nor, N.H.M.; de Ruijter, V.N.; Mintenig, S.M.; Kooi, M. Risk Assessment of Microplastic Particles. Nat. Rev. Mater. 2022, 7, 138–152. [Google Scholar] [CrossRef]
- Nelms, S.E.; Galloway, T.S.; Godley, B.J.; Jarvis, D.S.; Lindeque, P.K. Investigating Microplastic Trophic Transfer in Marine Top Predators. Environ. Pollut. 2018, 238, 999–1007. [Google Scholar] [CrossRef]
- Jacob, H.; Besson, M.; Swarzenski, P.W.; Lecchini, D.; Metian, M. Effects of Virgin Micro- and Nanoplastics on Fish: Trends, Meta-Analysis, and Perspectives. Environ. Sci. Technol. 2020, 54, 4733–4745. [Google Scholar] [CrossRef]
- Jaikumar, G.; Brun, N.R.; Vijver, M.G.; Bosker, T. Reproductive Toxicity of Primary and Secondary Microplastics to Three Cladocerans during Chronic Exposure. Environ. Pollut. 2019, 249, 638–646. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.F.; Xu, W.J.; Gu, Q.J.; Chen, Y.T.; Zhou, Q.M.; Zhang, L.; Gu, L.; Huang, Y.; Lyu, K.; Yang, Z. Small-Sized Microplastics Negatively Affect Rotifers: Changes in the Key Life-History Traits and Rotifer–Phaeocystis Population Dynamics. Environ. Sci. Technol. 2019, 53, 9241–9251. [Google Scholar] [CrossRef]
- Wang, X.; Liu, L.L.Q.Q.; Zheng, H.; Wang, M.X.; Fu, Y.X.; Luo, X.X.; Li, F.M.; Wang, Z.Y. Polystyrene Microplastics Impaired the Feeding and Swimming Behavior of Mysid Shrimp Neomysis japonica. Mar. Pollut. Bull. 2020, 150, 110660. [Google Scholar] [CrossRef]
- Qiao, R.X.; Sheng, C.; Lu, Y.F.; Zhang, Y.; Ren, H.Q.; Lemos, B. Microplastics Induce Intestinal Inflammation, Oxidative Stress, and Disorders of Metabolome and Microbiome in Zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef]
- Jeong, C.B.; Won, E.J.; Kang, H.M.; Lee, M.C.; Hwang, D.S.; Hwang, U.K.; Zhou, B.; Souissi, S.; Lee, S.J.; Lee, J.S. Microplastic Size-Dependent Toxicity, Oxidative Stress Induction, and p-JNK and p-p38 Activation in the Monogonont Rotifer (Brachionus koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef] [PubMed]
- Nanninga, G.B.; Horswill, C.; Lane, S.M.; Manica, A.; Briffa, M. Microplastic Exposure Increases Predictability of Predator Avoidance Strategies in Hermit Crabs. J. Hazard. Mater. Lett. 2020, 1, 100005. [Google Scholar] [CrossRef]
- Niziolek, M.; Bicka, M.; Osinka, A.; Samsel, Z.; Sekretarska, J.; Poprzeczko, M.; Bazan, R.; Fabczak, H.; Joachimiak, E.; Wloga, D. PCD Genes—From Patients to Model Organisms and Back to Humans. Int. J. Mol. Sci. 2022, 23, 1749. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.M.; Xia, H.M.; Xu, Y.N.; Xin, N.X.; Liu, J.; Zhang, S.H. Anesthetic Action of Volatile Anesthetics by Using Paramecium as a Model. J. Huazhong Univ. Sci. Technol. Med. Sci. 2012, 32, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Van Houten, J. Paramecium Biology. Results and Problems in Cell Differentiation. In Evo-Devo: Non-model Species in Cell and Developmental Biology; Tworzydlo, W., Bilinski, S., Eds.; Springer: Cham, Switzerland, 2019; Volume 68, pp. 291–318. [Google Scholar]
- Petrishchev, N.N.; Papayan, G.V.; Chistyakova, L.V.; Struy, A.V.; Faizullina, D.R. Effect of Photobiomodulation by Red and Infrared Laser Radiation on Motility of Paramecium Caudatum. J. Evol. Biochem. Physiol. 2018, 54, 457–464. [Google Scholar] [CrossRef]
- DeLong, J.P.; Lyon, S. Temperature Alters the Shape of Predator–Prey Cycles through Effects on Underlying Mechanisms. PeerJ 2020, 8, e9377. [Google Scholar] [CrossRef]
- Brette, R. Integrative Neuroscience of Paramecium, a “Swimming Neuron”. eNeuro 2021, 8, 0018–0021. [Google Scholar] [CrossRef]
- Arnaiz, O.; Meyer, E.; Sperling, L. Paramecium DB 2019: Integrating Genomic Data across the Genus for Functional and Evolutionary Biology. Nucleic Acids Res. 2020, 48, D599–D605. [Google Scholar]
- Bulannga, R.B.; Schmidt, S. Uptake and Accumulation of Microplastic Particles by Two Freshwater Ciliates Isolated from a Local River in South Africa. Environ. Res. 2022, 204, 112123. [Google Scholar] [CrossRef]
- Feng, J.X.; Zhao, H.S.; Gong, X.Y.; Xia, M.C.; Cai, L.S.; Yao, H.; Zhao, X.; Yan, Z.H.; Li, Z.P.; Nie, H.G.; et al. In Situ Identification and Spatial Mapping of Microplastic Standards in Paramecia by Secondary-Ion Mass Spectrometry Imaging. Anal. Chem. 2021, 93, 5521–5528. [Google Scholar] [CrossRef]
- Karakashian, M.W. Symbiosis in Paramecium Bursaria. Symp. Soc. Exp. Biol. 1975, 29, 145–173. [Google Scholar]
- Flores, E.; Thompson, L.; Sirisaengtaksin, N.; Nguyen, A.T.; Ballard, A.; Krachler, A.M. Using the Protozoan Paramecium Caudatum as a Vehicle for Food-Borne Infections in Zebrafish Larvae. J. Vis. Exp. 2019, 143, e58949. [Google Scholar] [CrossRef] [PubMed]
- Venkateswara Rao, J.; Srikanth, K.; Arepalli, S.K.; Gunda, V.G. Toxic Effects of Acephate on Paramecium Caudatum with Special Emphasis on Morphology, Behaviour, and Generation Time. Pestic. Biochem. Physiol. 2006, 86, 131–137. [Google Scholar] [CrossRef]
- Kryuchkova, M.; Danilushkina, A.; Lvov, Y.; Fakhrullin, R. Evaluation of Toxicity of Nanoclays and Graphene Oxide in Vivo: A Paramecium Caudatum Study. Environ. Sci. Nano 2016, 3, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Ershov, D.; Phan, M.S.; Pylvänäinen, J.W.; Rigaud, S.U.; Le Blanc, L.; Charles-Orszag, A.; Conway, J.; Laine, R.F.; Roy, N.H.; Bonazzi, D.; et al. TrackMate 7: Integrating State-of-the-Art Segmentation Algorithms into Tracking Pipelines. Nat. Methods 2022, 19, 829–832. [Google Scholar] [CrossRef]
- Jairo Kavana, N. Culture of Cyclops for Use the First Intermediate Host in Experimental Life Cycle of Spirometra Species. In Issues and Developments in Medicine and Medical: An Experimental Research; Karaman, R., Ed.; B P International: Hooghly, WB, India, 2022; Volume 9, pp. 81–85. [Google Scholar]
- Shi, X.H.; Zou, D.H.; Hu, S.S.; Mai, G.M.; Ma, Z.L.; Li, G. Photosynthetic Characteristics of Three Cohabitated Macroalgae in the Daya Bay, and Their Responses to Temperature Rises. Plants 2021, 10, 2441. [Google Scholar] [CrossRef]
- Botterell, Z.L.R.; Beaumont, N.; Dorrington, T.; Steinke, M.; Thompson, R.C.; Lindeque, P.K. Bioavailability and Effects of Microplastics on Marine Zooplankton: A Review. Environ. Pollut. 2019, 245, 98–110. [Google Scholar] [CrossRef]
- Du, J.; Xu, S.D.; Zhou, Q.W.; Li, H.X.; Fu, L.; Tang, J.H.; Wang, Y.Y.; Peng, X.; Xu, Y.T.; Du, X.P. A Review of Microplastics in the Aquatic Environmental: Distribution, Transport, Ecotoxicology, and Toxicological Mechanisms. Environ. Sci. Pollut. Res. 2020, 27, 11494–11505. [Google Scholar] [CrossRef]
- Franzellitti, S.; Canesi, L.; Auguste, M.; Wathsala, R.H.G.R.; Fabbri, E. Microplastic Exposure and Effects in Aquatic Organisms: A Physiological Perspective. Environ. Toxicol. Pharmacol. 2019, 68, 37–51. [Google Scholar] [CrossRef]
- Sridharan, S.; Kumar, M.; Bolan, N.S.; Singh, L.; Kumar, S.; Kumar, R.; You, S. Are Microplastics Destabilizing the Global Network of Terrestrial and Aquatic Ecosystem Services? Environ. Res. 2021, 198, 111243. [Google Scholar] [CrossRef]
- Chen, C.C.; Shi, Y.H.; Zhu, Y.J.; Zeng, J.M.; Qian, W.; Zhou, S.; Ma, J.; Pan, K.; Jiang, Y.L.; Tao, Y.; et al. Combined Toxicity of Polystyrene Microplastics and Ammonium Perfluorooctanoate to Daphnia magna: Mediation of Intestinal Blockage. Water Res. 2022, 219, 118536. [Google Scholar] [CrossRef] [PubMed]
- Aljaibachi, R.; Callaghan, A. Impact of Polystyrene Microplastics on Daphnia magna Mortality and Reproduction in Relation to Food Availability. PeerJ 2018, 6, e4601. [Google Scholar] [CrossRef]
- Bhagat, J.; Zang, L.; Nishimura, N.; Shimada, Y. Zebrafish: An Emerging Model to Study Microplastic and Nanoplastic Toxicity. Sci. Total Environ. 2020, 728, 138707. [Google Scholar] [CrossRef] [PubMed]
- Mayne, R.; Whiting, J.G.H.; Wheway, G.; Melhuish, C.; Adamatzky, A. Particle Sorting by Paramecium Cilia Arrays. Biosystems 2017, 156–157, 46–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayukai, T. Discriminate Feeding of the Calanoid Copepod Acartia Clausi in Mixtures of Phytoplankton and Inert Particles. Mar. Biol. 1987, 94, 579–587. [Google Scholar] [CrossRef]
- DeMott, W.R. Discrimination Between Algae and Detritus by Freshwater and Marine Zooplankton. Bullet. Mar. Sci. 1988, 43, 486–499. [Google Scholar]
- Frost, B.W. Feeding Behavior of Calanus Pacificus in Mixtures of Food Particles. Limnol. Oceanogr. 1977, 22, 472–491. [Google Scholar] [CrossRef]
- Kodama, Y.; Fujishima, M. Localization of Perialgal Vacuoles beneath the Host Cell Surface Is Not a Prerequisite Phenomenon for Protection from the Host’s Lysosomal Fusion in the Ciliate Paramecium Bursaria. Protist 2009, 160, 319–329. [Google Scholar] [CrossRef]
- Kodama, Y.; Fujishima, M. Timing of Perialgal Vacuole Membrane Differentiation from Digestive Vacuole Membrane in Infection of Symbiotic Algae Chlorella Vulgaris of the Ciliate Paramecium Bursaria. Protist 2009, 160, 65–74. [Google Scholar] [CrossRef]
- Wong, B.B.M.; Candolin, U. Behavioral Responses to Changing Environments. Behavl. Ecol. 2015, 26, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Ivantsova, E.; Souders, C.L.; Martyniuk, C.J. The Agrochemical S-Metolachlor Disrupts Molecular Mediators and Morphology of the Swim Bladder: Implications for Locomotor Activity in Zebrafish (Danio Rerio). Ecotoxicol. Environ. Saf. 2021, 208, 111641. [Google Scholar] [CrossRef] [PubMed]
- Barboza, L.G.A.; Vieira, L.R.; Guilhermino, L. Single and Combined Effects of Microplastics and Mercury on Juveniles of the European Seabass (Dicentrarchus Labrax): Changes in Behavioural Responses and Reduction of Swimming Velocity and Resistance Time. Environ. Pollut. 2018, 236, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; An, Y.J. Edible Size of Polyethylene Microplastics and Their Effects on Springtail Behavior. Environ. Pollut. 2020, 266, 115255. [Google Scholar] [CrossRef]
- Wang, W.L.; Zhang, J.L.; Qiu, Z.Q.; Cui, Z.Y.; Li, N.Q.; Li, X.; Wang, Y.W.; Zhang, H.X.; Zhao, C.Y. Effects of Polyethylene Microplastics on Cell Membranes: A Combined Study of Experiments and Molecular Dynamics Simulations. J. Hazard. Mater. 2022, 429, 128323. [Google Scholar] [CrossRef]
- Uguen, M.; Nicastro, K.R.; Zardi, G.I.; Gaudron, S.M.; Spilmont, N.; Akoueson, F.; Duflos, G.; Seuront, L. Microplastic Leachates Disrupt the Chemotactic and Chemokinetic Behaviours of an Ecosystem Engineer (Mytilus Edulis). Chemosphere 2022, 306, 135425. [Google Scholar] [CrossRef]
- Atema, J. Chemoreception in the Sea: Adaptations of Chemoreceptors and Behaviour to Aquatic Stimulus Conditions. Symp. Soc. Exp. Biol. 1985, 39, 387–423. [Google Scholar]
- Kirsch, D.R. Freshwater Gastropods as an Important Group for Studying the Impact of Inter- and Intra-Specific Chemical Communication on Aquatic Community Dynamics. Aquat. Ecol. 2022, 56, 361–375. [Google Scholar] [CrossRef]
- Gruzdev, G.A.; Karpukhina, O.V.; Inozemtsev, A.N.; Kamensky, A.A. Method for Primary Screening of Pharmaceuticals on the Paramecium Caudatum Eukaryotic Cell Model. Dokl. Biochem. Biophys. 2022, 502, 36–39. [Google Scholar] [CrossRef]
- Mayne, R.; Morgan, J.; Whiting, J.G.H.; Phillips, N.; Adamatzky, A. On Measuring Nanoparticle Toxicity and Clearance with Paramecium Caudatum. Sci. Rep. 2019, 9, 8957. [Google Scholar] [CrossRef] [Green Version]
- Sarvestani, A.N.; Shamloo, A.; Ahmadian, M.T. Simulation of Paramecium Chemotaxis Exposed to Calcium Gradients. Cell Biochem. Biophys. 2016, 74, 241–252. [Google Scholar] [CrossRef]
- Naitoh, Y. Control of the Orientation of Cilia by Adenosinetriphosphate, Calcium, and Zinc in Glycerol-Extracted Paramecium Caudatum. J. Gen. Physiol. 1969, 53, 517–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, M.S.; Van Houten, J. Ion Channels of Cilia: Paramecium as a Model. J. Eukaryot. Microbiol. 2022, 69, e12884. [Google Scholar] [CrossRef] [PubMed]
- Matés, J.M. Effects of Antioxidant Enzymes in the Molecular Control of Reactive Oxygen Species Toxicology. Toxicology 2000, 153, 83–104. [Google Scholar] [CrossRef]
- Kim, J.H.; Yu, Y.B.; Choi, J.H. Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Neurotoxicity in Fish Exposed to Microplastics: A Review. J. Hazard. Mater. 2021, 413, 125423. [Google Scholar] [CrossRef] [PubMed]
- Solomando, A.; Capó, X.; Alomar, C.; Álvarez, E.; Compa, M.; Valencia, J.M.; Pinya, S.; Deudero, S.; Sureda, A. Long-Term Exposure to Microplastics Induces Oxidative Stress and a pro-Inflammatory Response in the Gut of Sparus Aurata Linnaeus, 1758. Environ. Pollut. 2020, 266, 115295. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.H.; Sun, M.H.; Zhou, M.; Chang, Z.J.; Li, L. Polyvinyl Chloride Microplastics Induce Growth Inhibition and Oxidative Stress in Cyprinus Carpio Var. Larvae. Sci. Total Environ. 2020, 716, 136479. [Google Scholar] [CrossRef] [PubMed]
- Scandalios, J.G. The Rise of ROS. Trends Biochem. Sci. 2002, 27, 483–486. [Google Scholar] [CrossRef]
- Nishihara, N.; Horiike, S.; Takahashi, T.; Kosaka, T.; Shigenaka, Y.; Hosoya, H. Cloning and Characterization of Endosymbiotic Algae Isolated from Paramecium Bursaria. Protoplasma 1998, 203, 91–99. [Google Scholar] [CrossRef]
- Kodama, Y.; Suzuki, H.; Dohra, H.; Sugii, M.; Kitazume, T.; Yamaguchi, K.; Shigenobu, S.; Fujishima, M. Comparison of Gene Expression of Paramecium Bursaria with and without Chlorella Variabilis Symbionts. BMC Genom. 2014, 15, 183. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.H.; Liu, C.J.; Yu, Y.H.; Jhou, Y.T.; Fujishima, M.; Tsai, I.J.; Leu, J.Y. Genome Plasticity in Paramecium Bursaria Revealed by Population Genomics. BMC Biol. 2020, 18, 180. [Google Scholar] [CrossRef]
- Al-Ameeli, Z.T.; Al-Sammak, M.A.; DeLong, J.P.; Dunigan, D.D.; Van Etten, J.L. Catalysis of Chlorovirus Production by the Foraging of Bursaria Truncatella on Paramecia Bursaria Containing Endosymbiotic Algae. Microorganisms 2021, 9, 2170. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.Y.; Wang, Y.W.; Gong, Z.W.; Fan, X.P.; Ni, B. Toxic Effects of Copper Nanoparticles on Paramecium Bursaria–Chlorella Symbiotic System. Front. Microbiol. 2022, 13, 834208. [Google Scholar] [CrossRef] [PubMed]
- Stoecker, D.K.; Hansen, P.J.; Caron, D.A.; Mitra, A. Mixotrophy in the Marine Plankton. Annu. Rev. Mar. Sci. 2017, 9, 311–335. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Particles | Size, nm | Zeta-Potential, mV | PDI |
|---|---|---|---|
| Polystyrene microspheres | 1003 ± 24 | 38 ± 3 | 0.05 ± 0.016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Li, C.; Chen, X.; Li, Y.; Fei, C.; Chen, J. Paramecium bursaria as a Potential Tool for Evaluation of Microplastics Toxicity. Biology 2022, 11, 1852. https://doi.org/10.3390/biology11121852
Zhang J, Li C, Chen X, Li Y, Fei C, Chen J. Paramecium bursaria as a Potential Tool for Evaluation of Microplastics Toxicity. Biology. 2022; 11(12):1852. https://doi.org/10.3390/biology11121852
Chicago/Turabian StyleZhang, Jianhua, Changhong Li, Xiangrui Chen, Yunqian Li, Chenjie Fei, and Jiong Chen. 2022. "Paramecium bursaria as a Potential Tool for Evaluation of Microplastics Toxicity" Biology 11, no. 12: 1852. https://doi.org/10.3390/biology11121852