
Characterization of Acinetobacter baumannii Isolated from Raw Milk

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Milk Sampling
2.2. Isolation of Acinetobacter spp.
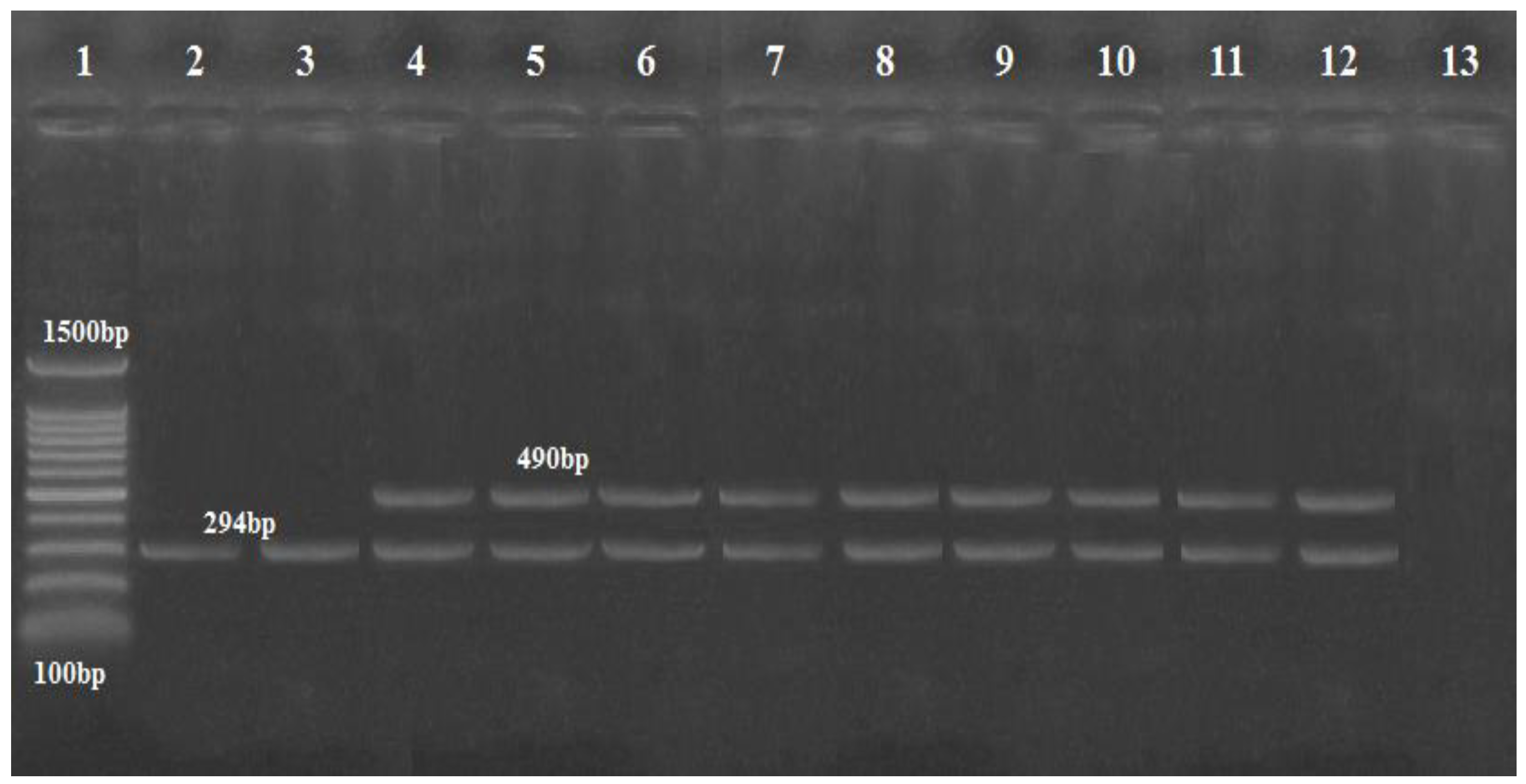
2.3. Genotypic Identification of A. baumannii
2.4. Antimicrobial Susceptibility Testing
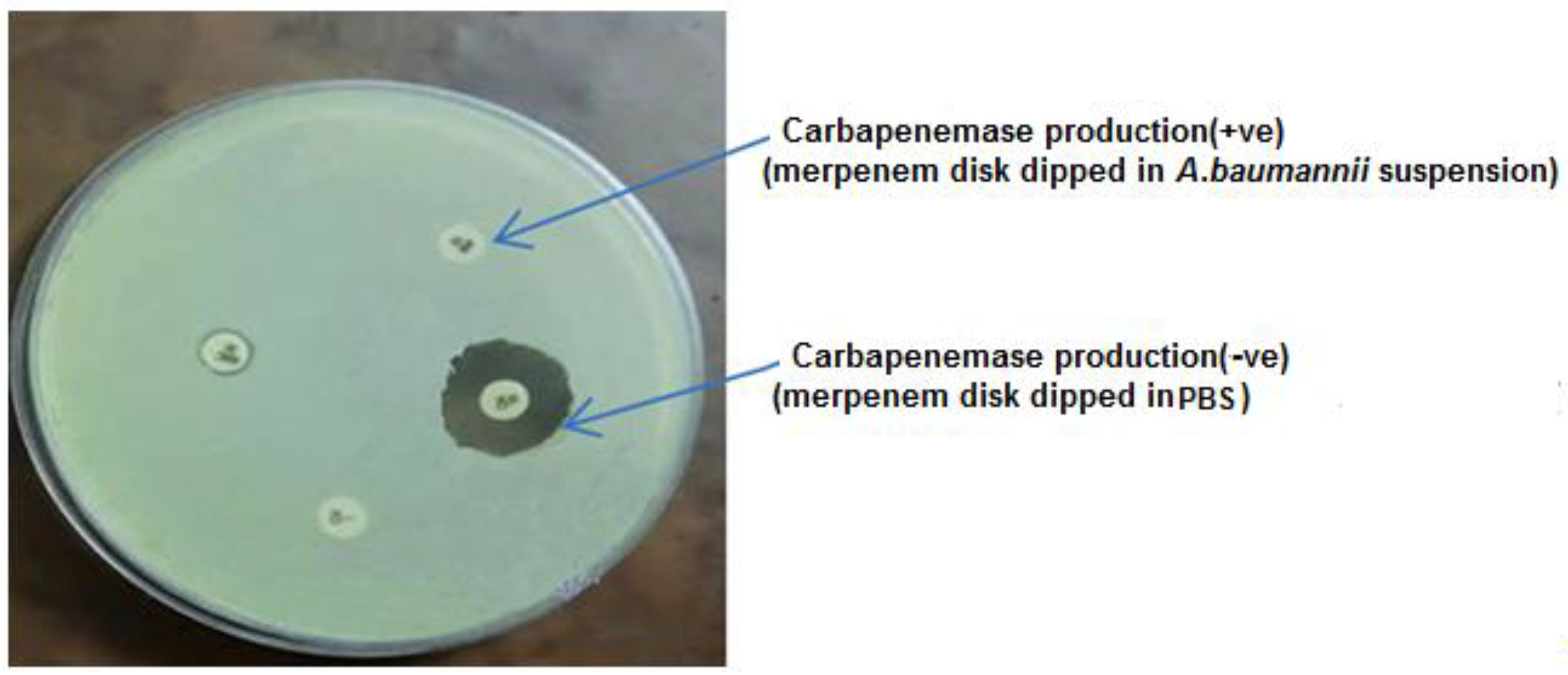
2.5. Screening of Metallo-β-lactamases Production Using the Disk Method
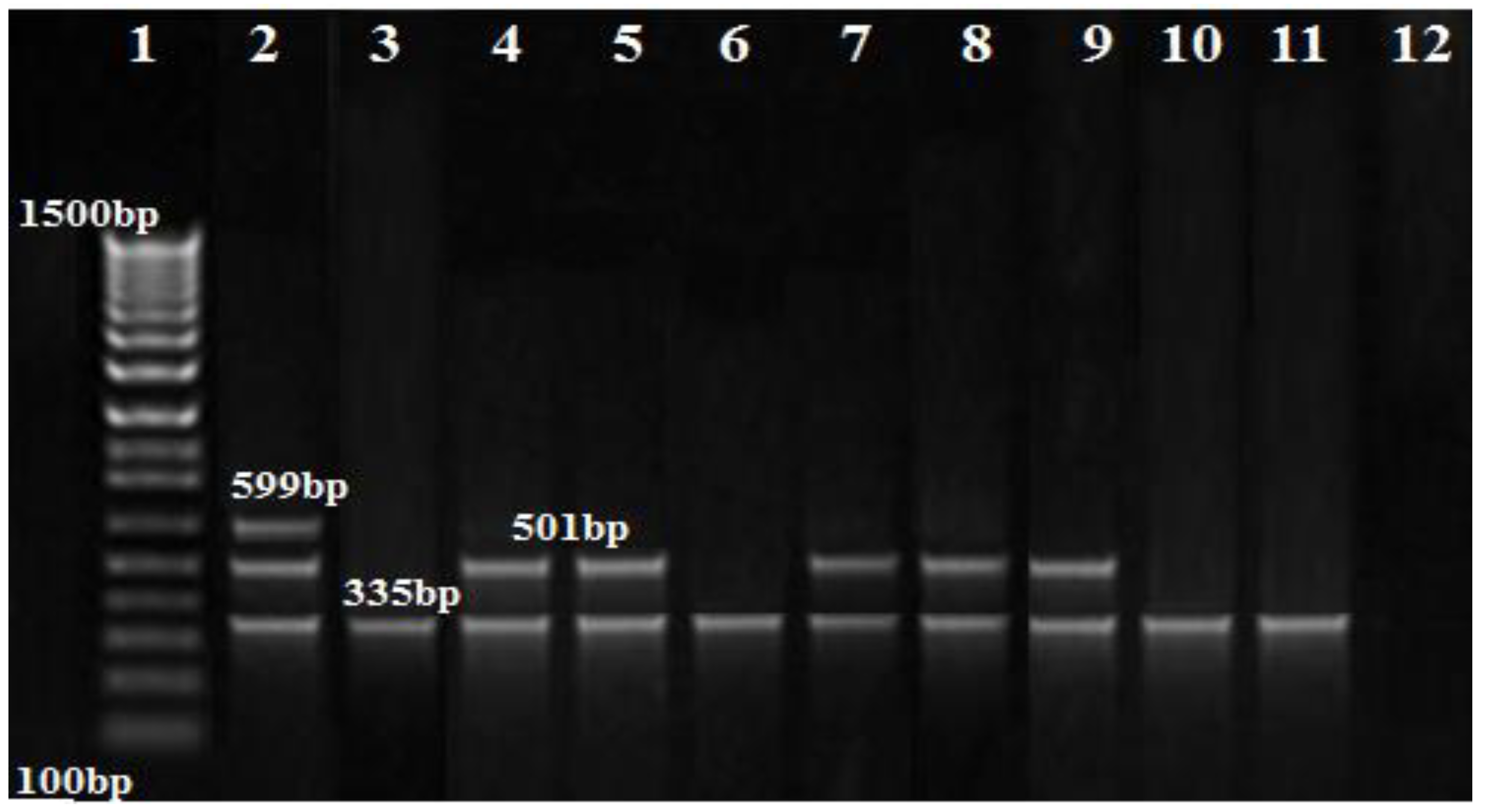
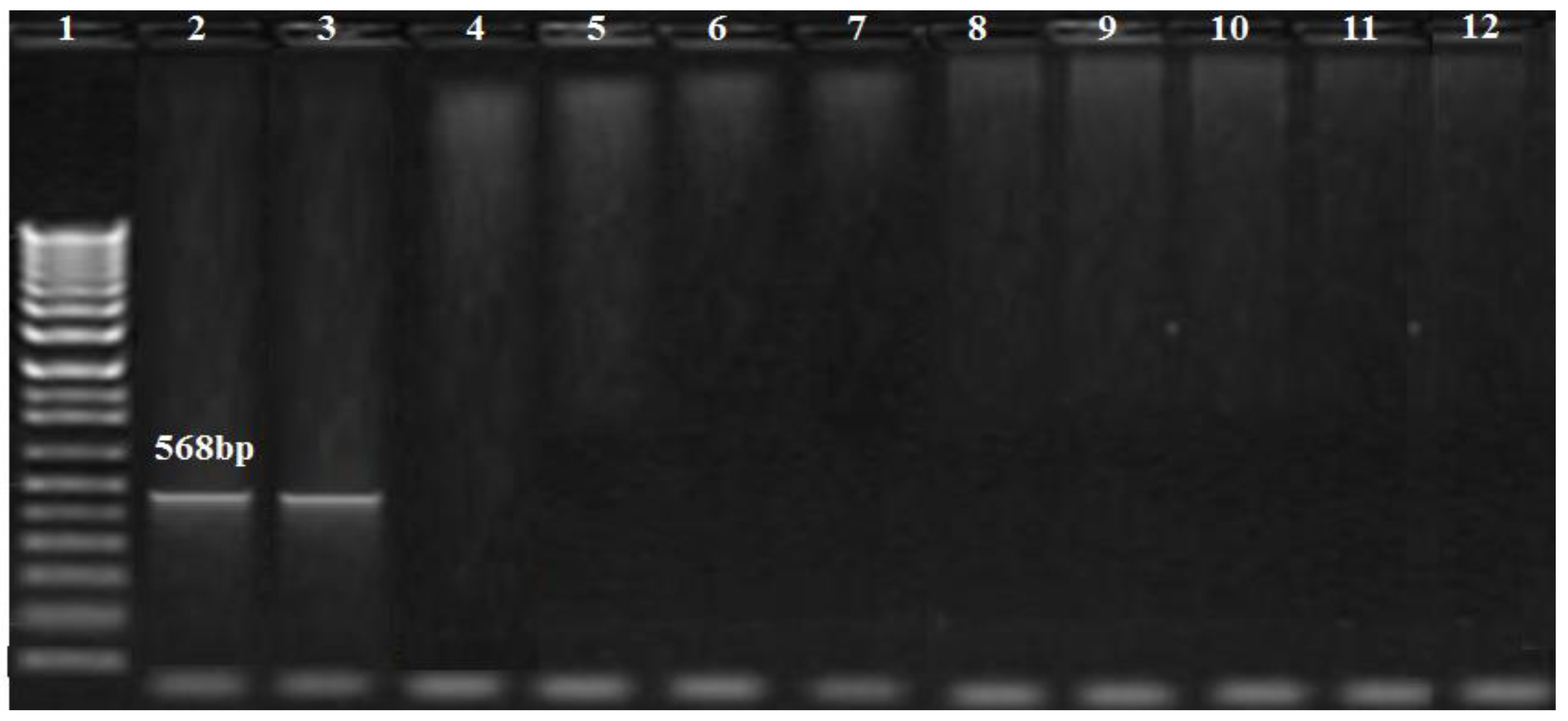
2.6. Detection of Metallo-β-lactamasesgenes
2.7. PCR Positive Control
2.8. Qualitative and Quantatine Methods for the Detction of Biofilm Formation
2.8.1. Congo Red Agar
2.8.2. Microtiter Plate Technique
3. Results
3.1. Results of the Isolation and Identification of A. baumannii
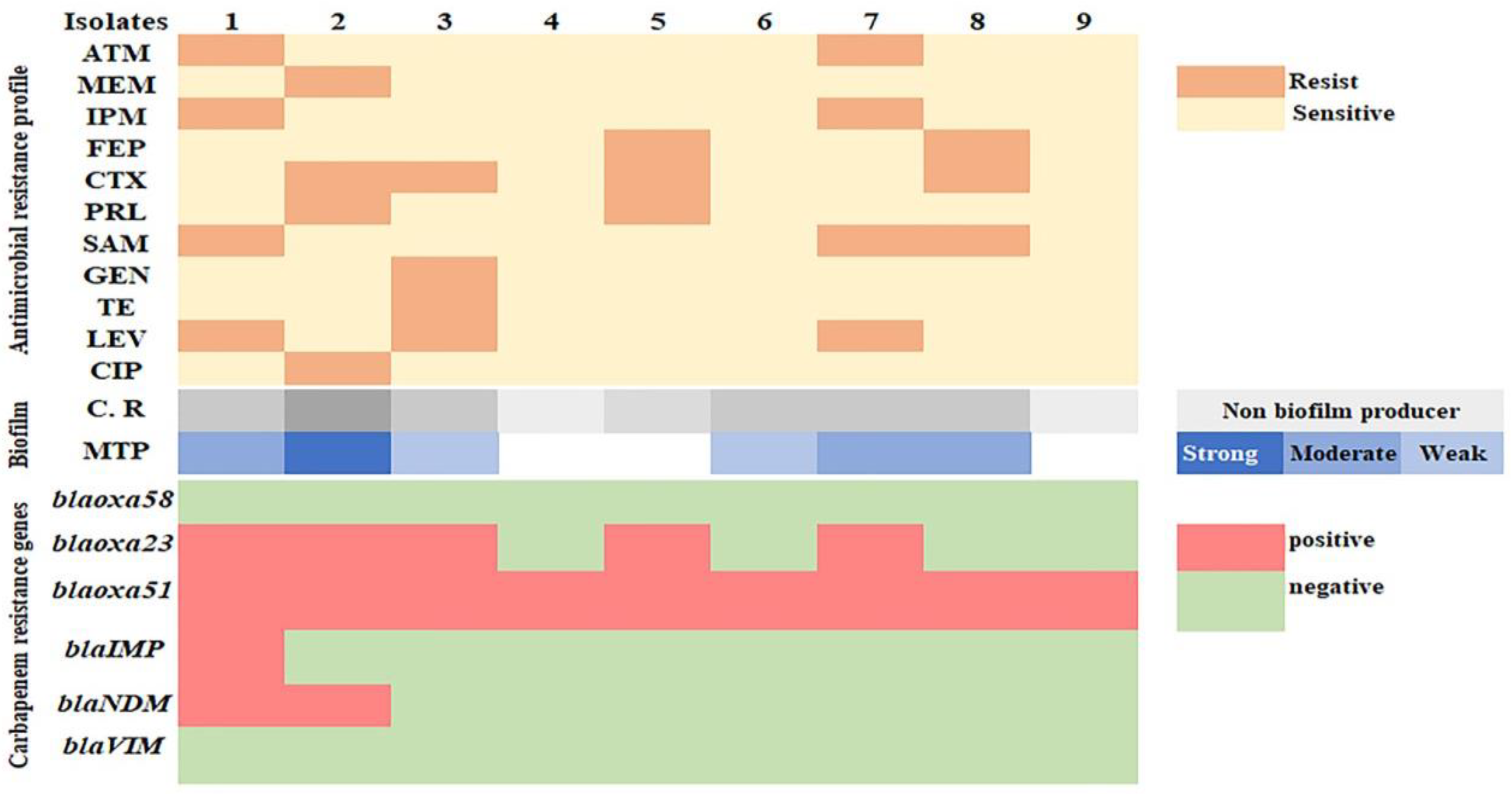
3.2. Antimicrobial Susceptibility Profile and MBL Production of A. baumannii Isolates
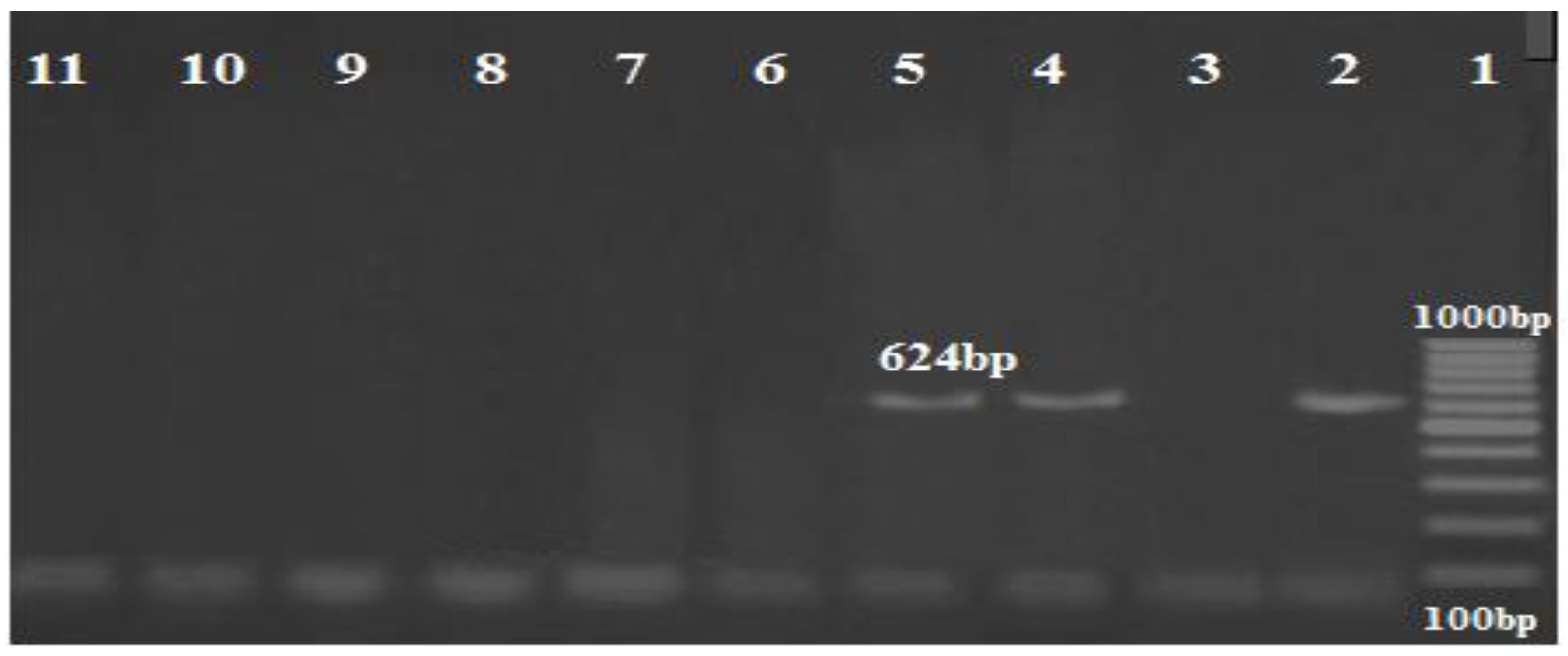
3.3. PCR Results of the Carbapenem Resistance Genes
3.4. Biofilm Formation of A. baumannii Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliver, S.P.; Boor, K.J.; Murphy, S.C.; Murinda, S.E. Food Safety Hazards Associated with Consumption of Raw Milk. Foodborne Pathog. Dis. 2009, 6, 793–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurung, M.; Nam, H.M.; Tamang, M.D.; Chae, M.H.; Jang, G.C.; Jung, S.C.; Lim, S.K. Prevalence and Antimicrobial Susceptibility of Acinetobacter from Raw Bulk Tank Milk in Korea. J. Dairy Sci. 2013, 96, 1997–2002. [Google Scholar] [CrossRef] [PubMed]
- Zucali, M.; Bava, L.; Tamburini, A.; Brasca, M.; Vanoni, L.; Sandrucci, A. Effects of Season, Milking Routine and Cow Cleanliness on Bacterial and Somatic Cell Counts of Bulk Tank Milk. J. Dairy Res. 2011, 78, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Kurcik-Trajkovska, B. Acinetobacter spp.—A Serious Enemy Threatening Hospitals Worldwide. Maced. J. Med. Sci. 2009, 2, 157–162. [Google Scholar] [CrossRef]
- Bergogne-Bérézin, E.; Towner, K.J. Acinetobacter spp. as Nosocomial Pathogens: Microbiological, Clinical, and Epidemiological Features. Clin. Microbiol. Rev. 1996, 9, 148–165. [Google Scholar] [CrossRef]
- Doughari, H.J.; Ndakidemi, P.A.; Human, I.S.; Benade, S. The Ecology, Biology and Pathogenesis of Acinetobacter spp.: An Overview. Microbes Environ. 2011, 26, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Jain, R.; Danziger, L.H. Multidrug-Resistant Acinetobacter Infections: An Emerging Challenge to Clinicians. Ann. Pharmacother. 2004, 38, 1449–1459. [Google Scholar] [CrossRef]
- Webster, C.; Towner, K.J.; Humphreys, H. Survival of Acinetobacter on Three Clinically Related Inanimate Surfaces. Infect. Control Hosp. Epidemiol. 2000, 21, 246. [Google Scholar] [CrossRef] [Green Version]
- Gallego, L.; Towner, K.J. Carriage of Class 1 Integrons and Antibiotic Resistance in Clinical Isolates of Acinetobacter baumannii from Northern Spain. J. Med. Microbiol. 2001, 50, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Lucey, J.A. Raw Milk Consumption: Risks and Benefits. Nutr. Today 2015, 50, 189–193. [Google Scholar] [CrossRef]
- EL-Shinawy, S.A.; EL-Kholy, A.D.; Meshref, A.; Sharkawy, S. Mycological evaluation of milk and some milk products in Beni-suef city. Assiut Vet. Med. J. 2018, 64, 117–122. [Google Scholar]
- Brooks, J.C.; Martinez, B.; Stratton, J.; Bianchini, A.; Krokstrom, R.; Hutkins, R. Survey of raw milk cheeses for microbiological quality and prevalence of foodborne pathogens. Food Microbiol. 2012, 31, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and Pathophysiological Overview of Acinetobacter Infections: A Century of Challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzer, H.J.F.; Rolling, T.; Schmiedel, S.; Klupp, E.M.; Lange, C.; Seifert, H. Severe Community-Acquired Bloodstream Infection with Acinetobacter ursingii in Person Who Injects Drugs. Emerg. Infect. Dis. 2016, 22, 134–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalheira, A.; Silva, J.; Teixeira, P. Acinetobacter spp. in Food and Drinking Water—A Review. Food Microbiol. 2021, 95, 103675. [Google Scholar] [CrossRef]
- Tiku, V. Acinetobacter baumannii: Virulence Strategies and Host Defense Mechanisms. DNA Cell Biol. 2022, 41, 43–48. [Google Scholar] [CrossRef]
- Logan, L.K.; Gandra, S.; Trett, A.; Weinstein, R.A.; Laxminarayan, R. Acinetobacter baumannii Resistance Trends in Children in the United States, 1999–2012. J. Pediatr. Infect. Dis. Soc. 2019, 8, 136–142. [Google Scholar] [CrossRef]
- Abbott, I.; Cerqueira, G.M.; Bhuiyan, S.; Peleg, A.Y. Carbapenem Resistance in Acinetobacter baumannii: Laboratory Challenges, Mechanistic Insights and Therapeutic Strategies. Expert Rev. Anti-Infect. Ther. 2013, 11, 395–409. [Google Scholar] [CrossRef]
- Sawa, T.; Kooguchi, K.; Moriyama, K. Molecular Diversity of Extended-Spectrum β-Lactamases and Carbapenemases, and Antimicrobial Resistance. J. Intensive Care 2020, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Bonomo, R.A. β-Lactamases: A Focus on Current Challenges. Cold Spring Harb. Perspect. Med. 2017, 7, a025239. [Google Scholar] [CrossRef]
- Eze, E.C.; Chenia, H.Y.; El Zowalaty, M.E. Acinetobacter baumannii Biofilms: Effects of Physicochemical Factors, Virulence, Antibiotic Resistance Determinants, Gene Regulation, and Future Antimicrobial Treatments. Infect. Drug Resist. 2018, 11, 2277. [Google Scholar] [CrossRef] [PubMed]
- Babapour, E.; Haddadi, A.; Mirnejad, R.; Angaji, S.-A.; Amirmozafari, N. Biofilm Formation in Clinical Isolates of Nosocomial Acinetobacter baumannii and Its Relationship with Multidrug Resistance. Asian Pac. J. Trop. Biomed. 2016, 6, 528–533. [Google Scholar] [CrossRef] [Green Version]
- Bardbari, A.M.; Arabestani, M.R.; Karami, M.; Keramat, F.; Alikhani, M.Y.; Bagheri, K.P. Correlation between Ability of Biofilm Formation with Their Responsible Genes and MDR Patterns in Clinical and Environmental Acinetobacter baumannii Isolates. Microb. Pathog. 2017, 108, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Yu, C.-P.; Fernández, G.E.; Hu, Z. Interactions of Nanosilver with Escherichia Coli Cells in Planktonic and Biofilm Cultures. Water Res. 2010, 44, 6095–6103. [Google Scholar] [CrossRef]
- Constantiniu, S.; Romaniuc, A.; Iancu, L.S.; Filimon, R.; Taraşi, I. Cultural and Biochemical Characteristics of Acinetobacter spp. Strains Isolated from Hospital Units. J. Prev. Med. 2004, 12, 35–42. [Google Scholar]
- La Scola, B.; Gundi, V.A.K.B.; Khamis, A.; Raoult, D. Sequencing of the RpoB Gene and Flanking Spacers for Molecular Identification of Acinetobacter species. J. Clin. Microbiol. 2006, 44, 827–832. [Google Scholar] [CrossRef] [Green Version]
- Barry, T.; Colleran, G.; Glennon, M.; Dunican, L.K.; Gannon, F. The 16s/23s Ribosomal Spacer Region as a Target for DNA Probes to Identify Eubacteria. PCR Methods Appl. 1991, 1, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Wisplinghoff, H.; Krut, O.; Seifert, H. A PCR-Based Method to Differentiate between Acinetobacter baumannii and Acinetobacter Genomic Species 13TU. Clin. Microbiol. Infect. 2007, 13, 1199–1201. [Google Scholar] [CrossRef] [Green Version]
- M100-S25; Performance Standards for Antimicrobial Susceptibility Testing. Twenty-Fifth Informational Supplement; CLSI: Berwyn, PA, USA, 2015.
- EUCAST. Antimicrob. Susceptibility Testing. Break. Tables Interpret. MICs Zo. Diameters. Version 5.0. 2015. Available online: http//www.eucast.org/clinical_breakpoints (accessed on 31 August 2015).
- Ayandele, A.A.; Oladipo, E.K.; Oyebisi, O.; Kaka, M.O. Prevalence of Multi-antibiotic Resistant Escherichia coli and Klebsiella species Obtained from a Tertiary Medical Institution in Oyo State, Nigeria. Qatar Med. J. 2020, 2020, 9. [Google Scholar] [CrossRef]
- van der Zwaluw, K.; de Haan, A.; Pluister, G.N.; Bootsma, H.J.; de Neeling, A.J.; Schouls, L.M. The carbapenem inactivation method (CIM), a simple and low-cost alternative for the Carba NP test to assess phenotypic carbapenemase activity in gram-negative rods. PLoS ONE 2015, 10, e0123690. [Google Scholar] [CrossRef] [Green Version]
- Shoja, S.; Moosavian, M.; Rostami, S.; Farahani, A.; Peymani, A.; Ahmadi, K.; Ebrahimifard, N. Dissemination of Carbapenem-Resistant Acinetobacter baumannii in Patients with Burn Injuries. J. Chin. Med. Assoc. 2017, 80, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Kazi, M.; Khot, R.; Shetty, A.; Rodrigues, C. Rapid Detection of the Commonly Encountered Carbapenemases (New Delhi Metallo-β-Lactamase, OXA-48/181) Directly from Various Clinical Samples Using Multiplex Real-Time Polymerase Chain Reaction Assay. Indian J. Med. Microbiol. 2018, 36, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New Method for Detecting Slime Production by Coagulase Negative Staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Hedayati, S.; Eftekhar, F.; Hosseini, S.M. Biofilm Formation by Bacteria Isolated from Intravenous Catheters. J. Med Bacteriol. 2014, 3, 26–31. [Google Scholar]
- Melo, D.H.; Souza, B.V.; Nascimento, J.S.; Amorim, A.M.B.; Medeiros, L.M.; Mattoso, J.M.V. A Reddish Problem: Antibiotic-Resistant Serratia marcescens in Dairy Food Commercialized in Rio de Janeiro. Int. Food Res. J. 2018, 25, 880–883. [Google Scholar]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of Staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Almasaudi, S.B. Acinetobacter spp. as Nosocomial Pathogens: Epidemiology and Resistance Features. Saudi J. Biol. Sci. 2018, 25, 586–596. [Google Scholar] [CrossRef] [Green Version]
- Ajao, A.O.; Robinson, G.; Lee, M.S.; Ranke, T.D.; Venezia, R.A.; Furuno, J.P.; Harris, A.D.; Johnson, J.K. Comparison of Culture Media for Detection of Acinetobacter baumannii in Surveillance Cultures of Critically-ill Patients. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 1425–1430. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.C.; Wei, Y.F.; Dijkshoorn, L.; Vaneechoutte, M.; Tang, C.T.; Chang, T.C. Species-Level Identification of Isolates of the Acinetobacter calcoaceticus-Acinetobacter baumannii Complex by Sequence Analysis of the 16S-23S RRNA Gene Spacer Region. J. Clin. Microbiol. 2005, 43, 1632–1639. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Nagao, M.; Matsumura, Y.; Hotta, G.; Matsushima, A.; Ito, Y.; Takakura, S.; Ichiyama, S. Regional Dissemination of Acinetobacter species Harbouring Metallo-β-Lactamase Genes in Japan. Clin. Microbiol. Infect. 2013, 19, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Maslunka, C.; Gifford, B.; Tucci, J.; Gürtler, V.; Seviour, R.J. Insertions or Deletions (Indels) in the Rrn 16S-23S rRNA Gene Internal Transcribed Spacer Region (ITS) Compromise the Typing and Identification of Strains within the Acinetobacter calcoaceticus-baumannii (Acb) Complex and Closely Related Members. PLoS ONE 2014, 9, e105390. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.G.; Lehmann, M.; Wisplinghoff, H.; Seifert, H. gyrB Multiplex PCR to Differentiate between Acinetobacter calcoaceticus and Acinetobacter Genomic Species 3. J. Clin. Microbiol. 2010, 48, 4592–4594. [Google Scholar] [CrossRef] [Green Version]
- Saad, N.M.; Amin, W.F.; Mostafa, S.M. Detection of Acinetobacter species in Milk and Some Dairy Products. Assiut Vet. Med. J. 2018, 64, 34–40. [Google Scholar]
- Jayarao, B.M.; Wang, L. A Study on the Prevalence of Gram-Negative Bacteria in Bulk Tank Milk. J. Dairy Sci. 1999, 82, 2620–2624. [Google Scholar] [CrossRef]
- Ndegwa, E.N.; Munyua, S.J.M.; Mulei, C.M. Prevalence of Microorganisms Associated with Udder Infections in Dairy Goats on Small-Scale Farms in Kenya. J. S. Afr. Vet. Assoc. 2001, 72, 97–98. [Google Scholar] [CrossRef] [PubMed]
- Lowe, M.; Ehlers, M.M.; Ismail, F.; Peirano, G.; Becker, P.J.; Pitout, J.D.D.; Kock, M.M. Acinetobacter baumannii: Epidemiological and Beta-Lactamase Data from Two Tertiary Academic Hospitals in Tshwane, South Africa. Front. Microbiol. 2018, 9, 1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.A.; Moon, H.W. In Vitro Activity of Antimicrobial Combination against Multidrug-Resistant Strains of Acinetobacter baumannii. Korean J. Lab. Med. 2005, 25, 312–316. [Google Scholar]
- Lee, K.; Yong, D.; Jeong, S.H.; Chong, Y. Multidrug-Resistant Acinetobacter Spp.: Increasingly Problematic Nosocomial Pathogens. Yonsei Med. J. 2011, 52, 879–89148. [Google Scholar] [CrossRef] [Green Version]
- Damier-Piolle, L.; Magnet, S.; Brémont, S.; Lambert, T.; Courvalin, P. AdeIJK, a Resistance-Nodulation-Cell Division Pump Effluxing Multiple Antibiotics in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2008, 52, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, R.; Dahiya, S.; Sayal, P. Evaluation of Multiple Antibiotic Resistance (MAR) Index and Doxycycline Susceptibility of Acinetobacter Species among Inpatients. Indian J. Microbiol. Res. 2016, 3, 299. [Google Scholar] [CrossRef]
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2016, 3, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Figueiredo, S.; Cattoir, V.; Carattoli, A.; Nordmann, P. Acinetobacter Radioresistens as a Silent Source of Carbapenem Resistance for Acinetobacter Spp. Antimicrob. Agents Chemother. 2008, 52, 1252–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saranathan, R.; Vasanth, V.; Vasanth, T.; Shabareesh, P.R.V.; Shashikala, P.; Devi, C.S.; Kalaivani, R.; Asir, J.; Sudhakar, P.; Prashanth, K. Emergence of Carbapenem Non-susceptible Multidrug Resistant Acinetobacter baumannii Strains of Clonal Complexes 103B and 92B Harboring OXA-type Carbapenemases and Metallo-β-lactamases in Southern India. Microbiol. Immunol. 2015, 59, 277–284. [Google Scholar] [CrossRef]
- Patrice, N.; Poirel, L.; Carrër, A.; Toleman, M.A.; Walsh, T.R. How to Detect NDM-1 Producers. J. Clin. Microbiol. 2011, 49, 718–721. [Google Scholar]
- Baquero, F.; Alvarez-Ortega, C.; Martinez, J.L. Ecology and evolution of antibiotic resistance. Environ. Microbiol. Rep. 2009, 1, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Turton, J.F.; Woodford, N.; Glover, J.; Yarde, S.; Kaufmann, M.E.; Pitt, T.L. Identification of Acinetobacter baumannii by Detection of the blaOXA-51-like Carbapenemase Gene Intrinsic to This Species. JCM J. Clin. Microbiol. 2006, 44, 2974–2976. [Google Scholar] [CrossRef] [Green Version]
- Marsik, F.J.; Nambiar, S. Review of Carbapenemases and AmpC-Beta Lactamases. Pediatr. Infect. Dis. J. 2011, 30, 1094–1095. [Google Scholar] [CrossRef]
- Chatterjee, S.; Datta, S.; Roy, S.; Ramanan, L.; Saha, A.; Viswanathan, R.; Som, T.; Basu, S. Carbapenem Resistance in Acinetobacter baumannii and Other Acinetobacter Spp. Causing Neonatal Sepsis: Focus on NDM-1 and Its Linkage to IS Aba125. Front. Microbiol. 2016, 7, 1126. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, N.; Agus, N.; Bozcal, E.; Uzel, A. Prevalence and Molecular Characterisation of Metallo-Beta-Lactamase Producing Strains of Imipenem-Resistant Pseudomonas aeruginosa in Turkey. Indian J. Med. Microbiol. 2014, 32, 349. [Google Scholar] [CrossRef]
- Pajand, O.; Rezaee, M.A.; Nahaei, M.R.; Mahdian, R.; Aghazadeh, M.; Soroush, M.H.; Tabrizi, M.S.; Hojabri, Z. Study of the Carbapenem Resistance Mechanisms in Clinical Isolates of Acinetobacter baumannii: Comparison of Burn and Non-Burn Strains. Burns 2013, 39, 1414–1419. [Google Scholar] [CrossRef]
- Malta, R.C.R.; Ramos, G.L.d.P.A.; dos Santos Nascimento, J. From Food to Hospital: We Need to Talk about Acinetobacter Spp. Germs 2020, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Azizi, O.; Shahcheraghi, F.; Salimizand, H.; Modarresi, F.; Shakibaie, M.R.; Mansouri, S.; Ramazanzadeh, R.; Badmasti, F.; Nikbin, V. Molecular Analysis and Expression of Bap Gene in Biofilm-Forming Multi-Drug-Resistant Acinetobacter baumannii. Rep. Biochem. Mol. Biol. 2016, 5, 62. [Google Scholar] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for Combating Bacterial Biofilms: A Focus on Anti-Biofilm Agents and Their Mechanisms of Action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [Green Version]
- Zarrilli, R.; Giannouli, M.; Tomasone, F.; Triassi, M.; Tsakris, A. Carbapenem Resistance in Acinetobacter baumannii: The Molecular Epidemic Features of an Emerging Problem in Health Care Facilities. J. Infect. Dev. Ctries. 2009, 3, 335–341. [Google Scholar] [CrossRef]
- Melo, P.D.C.; Ferreira, L.M.; Nader Filho, A.; Zafalon, L.F.; Vicente, H.I.G.; Souza, V.D. Comparison of methods for the detection of biofilm formation by Staphylococcus aureus isolated from bovine subclinical mastitis. Braz. J. Microbiol. 2013, 44, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Gaddy, J.A.; Actis, L.A. Regulation of Acinetobacter baumannii biofilm formation. Future Microbiol. 2009, 4, 273–278. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | rpoB Gene | 16S-23SrRNA Intergenic Space | |||
|---|---|---|---|---|---|
| Strain Name on Gene Bank | Spacer Region Zone 1 | Spacer Region Zone 2 | Identified Strains | Accession Number | Identified A. baumannii–A. calcoaceticus Complex |
| AcbaQHM_1 | OP326286 | OP326299 | A. baumannii | OP321175 | A. baumannii |
| AcbaQHM_2 | OP326287 | OP326300 | A. baumannii | OP321173 | A. baumannii |
| AcbaQHM_3 | OP326288 | OP326301 | A. baumannii | OP321177 | A. baumannii |
| AcbaQHM_4 | OP326289 | OP326302 | A. baumannii | OP321174 | A. baumannii |
| AcbaQHM_5 | OP326290 | OP326303 | A. baumannii | OP321176 | A. baumannii |
| AcbaQHM_6 | OP326291 | OP326304 | A. baumannii | OP321178 | A. baumannii |
| AcbaQHM_7 | OP326292 | OP326305 | A. baumannii | OP321258 | A. baumannii |
| AcbaQHM_8 | OP326293 | OP326306 | A. baumannii | OP321257 | A. baumannii |
| AcbaQHM_9 | OP326294 | OP326307 | A. baumannii | OP321259 | A. baumannii |
| AcbiQHM_10 | OP326295 | OP326308 | A. pittii | OP321256 | A. pittii |
| AcbiQHM_11 | OP326296 | OP326309 | A. pittii | OP321255 | A. pittii |
| ACspQHM_12 | OP326297 | OP326310 | A.oryzae | ------- | ----- |
| AcspQHM_13 | OP326298 | OP326311 | A.rudis | ------ | ------ |
| Antibiotic Classes | Antimicrobial | Susceptible Isolates | Resistant Isolates |
|---|---|---|---|
| Fluoroquinolones | Ciprofloxacin Levofloxacin | 8 (88.8%) 6 (66.6%) | 1 (11.1%) 3 (33.3%) |
| Tetracyclines | Tetracycline | 8 (88.8%) | 1 (11.1%) |
| Aminoglycosides | Amikacin Gentamicin | 9 (100%) 8 (88.8%) | 0 (0%) 1 (11.1%) |
| Penicillin | Ampicillin-sulbactam Piperacillin | 6 (66.6%) 7 (77.7%) | 3 (33.3%) 2 (22.2%) |
| Monobactams | Aztreonam | 7 (77.7%) | 2 (22.2%) |
| Cephalosporins | Cefotaxime Cefepime | 5 (55.5%) 7 (77.7%) | 4 (44.4%) 2 (22.2%) |
| Carbapenems | Imipenem Meropenem | 7 (77.7%) 7 (77.7%) | 2 (22.2%) 2 (22.2%) |
| Antibiotic Patterns | No. of Isolates | MAR Index |
|---|---|---|
| levofloxacin, ampicillin-sulbactam, imipenem, aztreonam | 2 | 0.33 |
| ciprofloxacin, piperacillin, cefotaxime, meropenem | 1 | 0.33 |
| ampicillin-sulbactam, cefotaxime, cefepime | 1 | 0.20 |
| levofloxacin, tetracyclins, gentamycin, cefotaxime | 1 | 0.33 |
| piperacillin, cefotaxime, cefepime, meropenem | 1 | 0.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, H.M.A.; Abd-Elhafeez, H.H.; Al-Jabr, O.A.; El-Zamkan, M.A. Characterization of Acinetobacter baumannii Isolated from Raw Milk. Biology 2022, 11, 1845. https://doi.org/10.3390/biology11121845
Mohamed HMA, Abd-Elhafeez HH, Al-Jabr OA, El-Zamkan MA. Characterization of Acinetobacter baumannii Isolated from Raw Milk. Biology. 2022; 11(12):1845. https://doi.org/10.3390/biology11121845
Chicago/Turabian StyleMohamed, Hams M. A., Hanan H. Abd-Elhafeez, Omar A. Al-Jabr, and Mona A. El-Zamkan. 2022. "Characterization of Acinetobacter baumannii Isolated from Raw Milk" Biology 11, no. 12: 1845. https://doi.org/10.3390/biology11121845