
Differential Expression of Dickkopf 1 and Periostin in Mouse Strains with High and Low Bone Mass
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Mice, Materials, and Methods
2.1. Animals
2.2. Sample Preparation
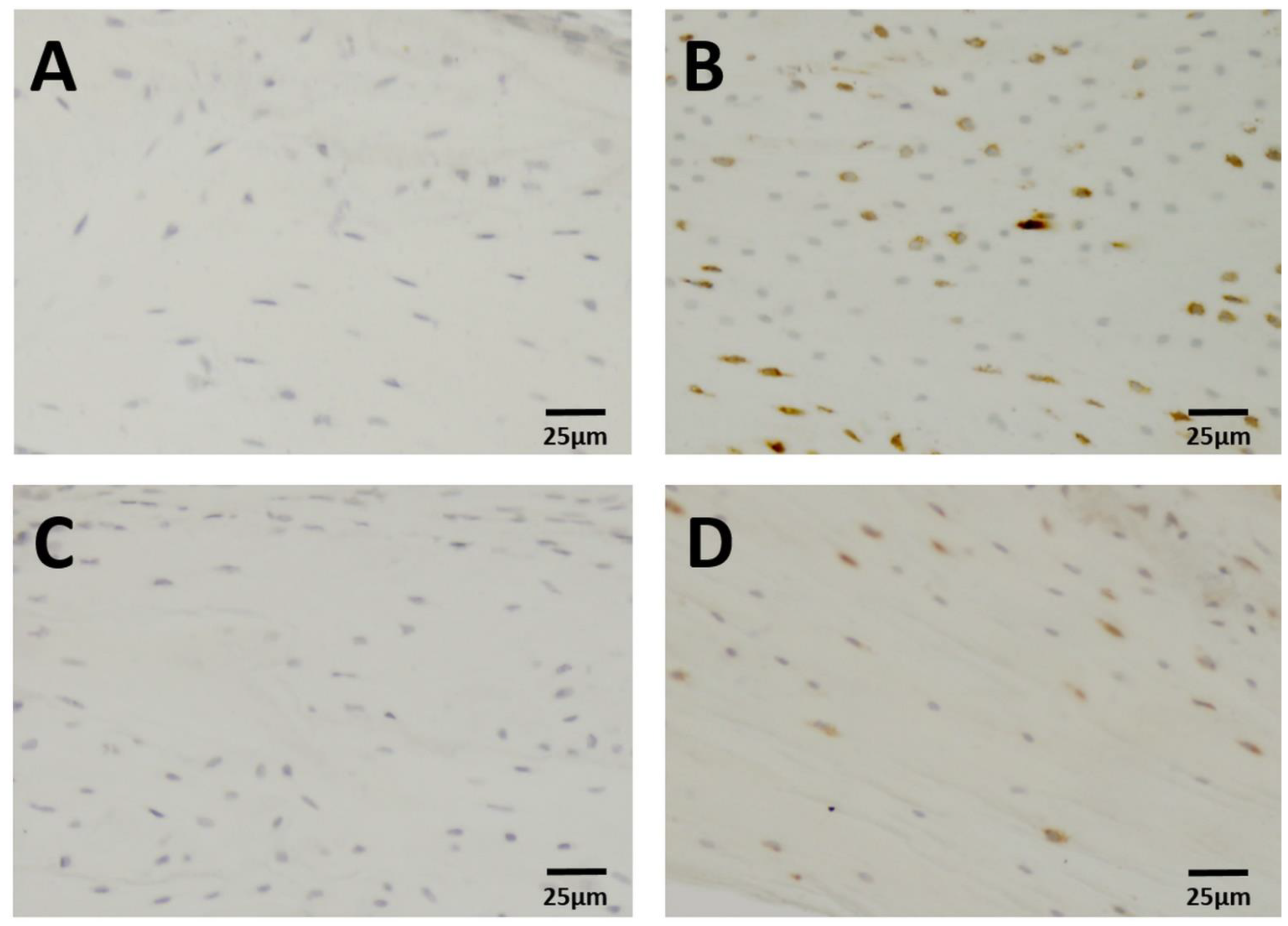
2.3. Immunohistochemistry
2.4. Histomorphometry
2.5. Statistical Analysis
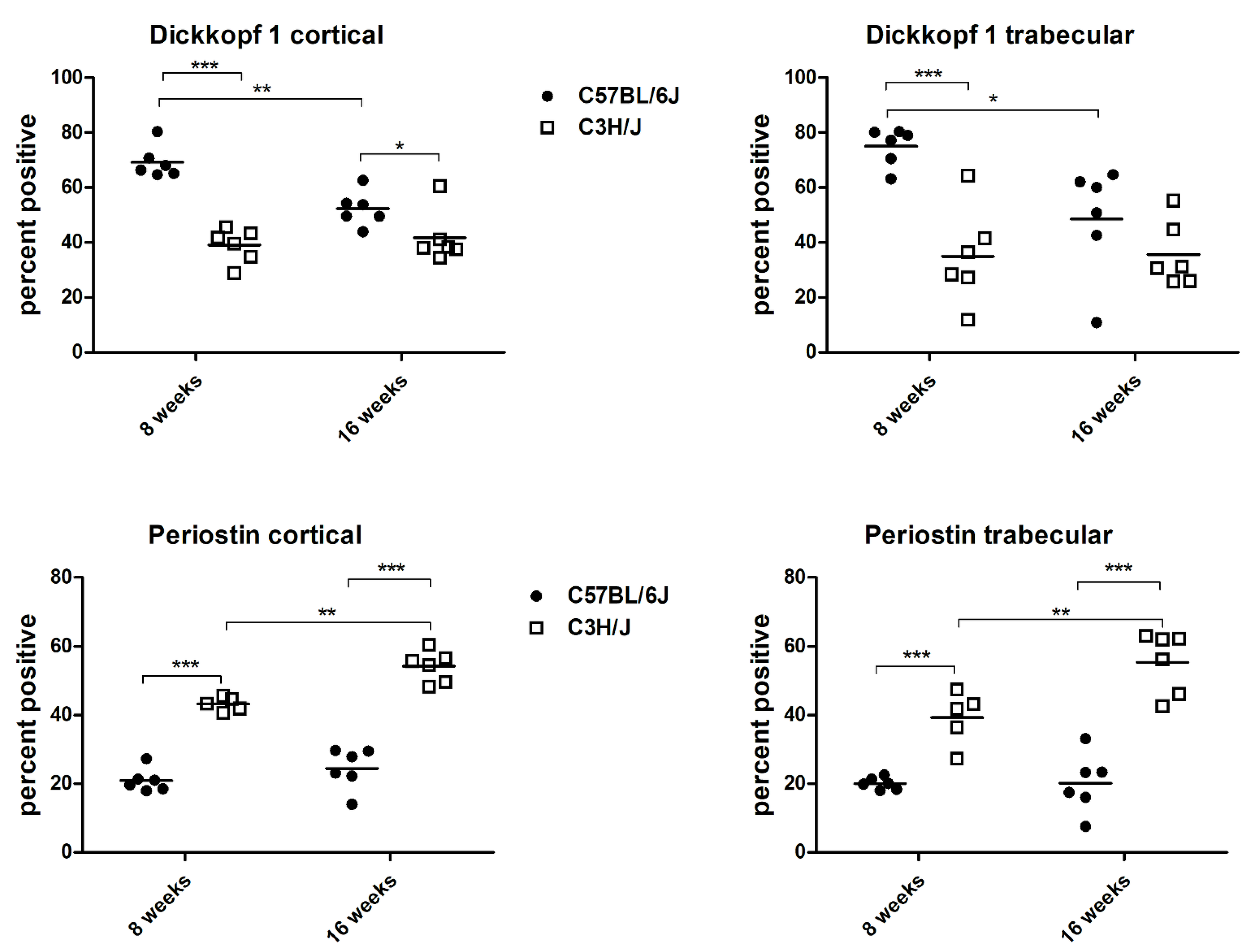
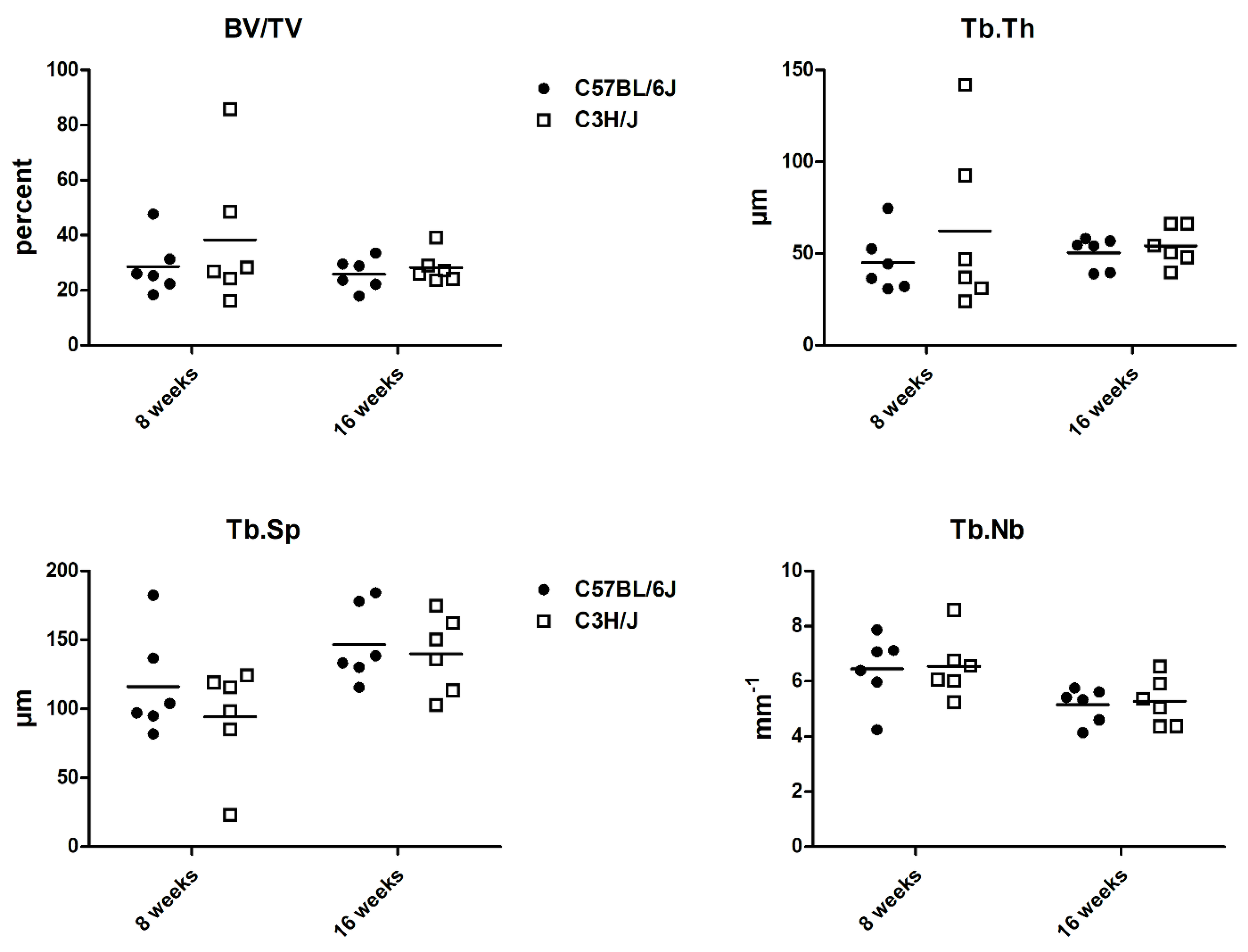
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonewald, L.F. The role of the osteocyte in bone and non-bone disease. Endocrinol. Metab. Clin. N. Am. 2017, 46, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saphir-Koren, R.; Livshits, G. Osteocyte control of bone remodeling: Is sclerostin a key molecular coordinator of the balanced bone resorption- formation cyles? Osteoporos. Int. 2014, 25, 2685–2700. [Google Scholar] [CrossRef] [PubMed]
- Rossini, M.; Gatti, D.; Adami, S. Involvement of WNT/β-catenin signaling in the treatment of osteoporosis. Calcif. Tissue Int. 2013, 93, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Gong, S.; Wang, R.; Liu, S.; Wang, B.; Chen, G.; Gong, T.; Xu, W. Knockdown of POSTN inhibits osteogenic differentiation of mesenchymal stem cells from patients with steroid-induced osteoporosis. Front. Cell Dev. Biol. 2020, 8, 606289. [Google Scholar] [CrossRef]
- Rodelo-Haad, C.; Santamaria, R.; Muñoz-Castañeda, J.R.; Pendón-Ruiz de Mier, M.V.; Martin-Malo, A.; Rodriguez, M. FGF23, biomarker or target? Toxins 2019, 11, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellido, T. Osteocyte-driven bone remodeling. Calcif. Tissue Int. 2014, 94, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papageorgiou, M.; Föger-Samwald, U.; Wahl, K.; Kerschan-Schindl, K.; Pietschmann, P. Age- and strain-related differences in bone microstructure and body composition during development in inbred male mouse strains. Calcif. Tissue Int. 2020, 106, 431–443. [Google Scholar] [CrossRef] [Green Version]
- Tiede-Lewis, L.M.; Xie, Y.; Hulbert, M.A.; Campos, R.; Dallas, M.R.; Dusevich, V.; Bonewald, L.F.; Dallas, S.L. Degeneration of the osteocyte network in the C57BL/6J mouse model of aging. Aging 2017, 9, 2190–2208. [Google Scholar] [CrossRef] [Green Version]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Min. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef] [Green Version]
- Colditz, J.; Thiele, S.; Baschant, U.; Niehrs, C.; Bonewald, L.F.; Hofbauer, L.C.; Rauner, M. Postnatal skeletal deletion of dickkopf-1 increases bone formation and bone volume in male and female mice, despite increased sclerostin expression. J. Bone Min. Res. 2018, 33, 1698–1707. [Google Scholar] [CrossRef]
- Morse, A.; Cheng, T.L.; Schindeler, A.; McDonald, M.M.; Mohanty, S.T.; Kneissel, M.; Kramer, I.; Little, D.G. DKK1 KO mice treated with sclerostin antibody have additional increases in bone volume. Calcif. Tissue Int. 2018, 103, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Tsourdi, E.; Colditz, J.; Lademann, F.; Rijntjes, E.; Köhrle, J.; Niehrs, C.; Hofbauer, L.C.; Rauner, M. The Role of Dickkopf-1 in Thyroid Hormone–Induced changes of bone remodeling in male mice. Endocrinology 2019, 160, 664–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goes, P.; Dutra, C.; Lösser, L.; Hofbauer, L.C.; Rauner, M.; Thiele, S. Loss of Dkk-1 in Osteocytes Mitigates Alveolar Bone Loss in Mice with Periodontitis. Front. Immunol. 2019, 10, 2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramli, R.R.; Chin, K.Y. A review of the potential application of osteocyte-related biomarkers, fibroblast growth factor-23, sclerostin, and dickkopf-1 in predicting osteoporosis and fractures. Diagnostics 2020, 10, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueland, T.; Stilgren, L.; Bollerslev, J. Bone Matrix Levels of Dickkopf and Sclerostin are Positively Correlated with Bone Mass and Strength in Postmenopausal Osteoporosis. Int. J. Mol. Sci. 2019, 20, 2896. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Dong, Z.; Hui, Z.; Aifei, W.; Lianfu, D.; Youjia, X. Bone sclerostin and dickkopf-related protein-1 are positively correlated with bone mineral density, bone microarchitecture, and bone strength in postmenopausal osteoporosis. BMC Musculoskelet. Disord. 2021, 22, 480. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Joiner, D.M.; Oyserman, S.M.; Sharma, P.; Goldstein, S.A.; He, X.; Hauschka, P.V. Bone mass is inversely proportional to Dkk1 levels in mice. Bone 2007, 41, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Zhang, Z.; Yu, Y.; Chu, H.Y.; Yu, S.; Yao, S.; Zhang, G.; Zhang, B.-T. Drug discovery of DKK1 inhibitors. Front. Pharmcol. 2022, 13, 847387. [Google Scholar] [CrossRef]
- Li, X.; Ominsky, M.S.; Niu, Q.-T.; Sun, N.; Daugherty, B.; D’Agostin, D.; Kurahara, C.; Gao, Y.; Cao, J.; Gong, J.; et al. Targeted deletion of the sclerostin gene in mice results in increased bone formation and bone strength. J. Bone Min. Res. 2008, 23, 860–869. [Google Scholar] [CrossRef]
- Cosman, F.; Crittenden, D.B.; Adachi, J.D.; Binkley, N.; Czerwinski, E.; Ferrari, S.; Hofbauer, L.C.; Lau, E.; Lewiecki, E.M.; Miyauchi, A.; et al. Romosozumab treatment in postmenopausal women with osteoporosis. N. Engl. J. Med. 2016, 375, 1532–1543. [Google Scholar] [CrossRef]
- Cosman, F.; Crittenden, D.B.; Ferrari, S.; Khan, A.; Lane, N.E.; Lippuner, K.; Matsumoto, T.; Milmont, C.E.; Libanati, C.; Grauer, A. FRAME Study: The foundation effect of building bone with 1 year of romosozumab leads to continued lower fracture risk after transition to denosumab. J. Bone Min. Res. 2018, 33, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Patsch, J.M.; Kohler, T.; Berzlanovich, A.; Muschitz, C.; Bieglmayr, C.; Roschger, P.; Resch, H.; Pietschmann, P. Trabecular bone microstructure and local gene expression in iliac crest biopsies of men with idiopathic osteoporosis. J. Bone Min. Res. 2011, 26, 1584–1592. [Google Scholar] [CrossRef] [PubMed]
- Coulson, J.; Bagley, L.; Barnouin, Y.; Bradburn, S.; Butler-Browne, G.; Gapeyeva, H.; Hogrel, J.-Y.; Maden-Wilkinson, T.; Maier, A.B.; Meskers, C.; et al. Circulating levels of dickkopf-1, osteoprotegerin and sclerostin are higher in old compared with young men and women and positively associated with whole-body bone mineral density in older adults. Osteoporos. Int. 2017, 28, 2683–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Büttner, A.; Albiol, L.; Julien, C.; Thiele, T.; Figge, C.; Kramer, I.; Kneissel, M.; Duda, G.N.; Checa, S.; et al. Cortical bone adaptation to a moderate level of mechanical loading in male Sost deficient mice. Sci. Rep. 2020, 10, 22299. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, N.; Standley, K.N.; Bianchi, E.N.; Stadelmann, V.; Foti, M.; Conway, S.J.; Ferrari, S.L. The matricellular protein periostin is required for sost inhibition and the anabolic response to mechanical loading and physical activity. J. Biol. Chem. 2009, 284, 35939–35950. [Google Scholar] [CrossRef] [Green Version]
- Gerbaix, M.; Vico, L.; Ferrari, S.L.; Bonnet, N. Periostin expression contributes to cortical bone loss during unloading. Bone 2015, 71, 94–100. [Google Scholar] [CrossRef]
- Walsh, J.S.; Gossiel, F.; Scott, J.R.; Paggiosi, M.A.; Eastell, R. Effect of age and gender on serum periostin: Relationship to cortical measures, bone turnover and hormones. Bone 2017, 99, 8–13. [Google Scholar] [CrossRef]
- Bonnet, N.; Conway, S.J.; Ferrari, S.L. Regulation of beta catenin signaling and parathyroid hormone anabolic effects in bone by the matricellular protein periostin. Proc. Natl. Acad. Sci. USA 2012, 109, 15048–15053. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, X.; Wang, X.; Miao, P.; Liu, J.; Li, C.; Li, D.; Zhou, W.; Jin, Z.; Cao, M. Periostin mediates oestrogen-induced osteogenic differentiation of bone marrow stromal cells in ovariectomised rats. BioMed Res. Int. 2020, 2020, 9405909. [Google Scholar] [CrossRef]
- Sirikul, W.; Siri-Angkul, N.; Chattipakorn, N.; Chattipakorn, S.C. Fibroblast growth factor 23 and osteoporosis: Evidence from bench to bedside. Int. J. Mol. Sci. 2022, 23, 2500. [Google Scholar] [CrossRef]
- Whyte, M.P.; Obrecht, S.E.; Finnegan, P.M.; Jones, J.L.; Podgornik, M.N.; McAlister, W.H.; Mumm, S. Osteoprotegerin deficiency and juvenile Paget’s disease. New Engl. J. Med. 2002, 347, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Nava-Valdivia, C.A.; Ponce-Guarneros, J.M.; Saldaña-Cruz, A.M.; Corona-Sanchez, E.G.; Ramirez-Villafaña, M.; Perez-Guerrero, E.E.; Murillo-Saich, J.D.; Contreras-Haro, B.; Vazquez-Villegas, M.L.; Gonzalez-Ponce, F.; et al. Assessment of serum sRANKL, sRANKL/OPG ratio, and other bone turnover markers with the estimated 10-year risk of major and hip osteoporotic fractures in rheumatoid arthritis: A cross-sectional study. BioMed Res. Int. 2021, 2021, 5567666. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, M.; Pitel, K.S.; Bruinsma, E.S.; Monroe, D.G.; Hawse, J.R. TIEG an destrogen modulate SOST expression in the murine skeleton. J. Cell. Physiol. 2018, 233, 3540–3551. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variable | C57BL/6J—8 we | C3H/J—8 we | C57BL/6J—16 we | C3H/J—16 we |
|---|---|---|---|---|
| DKK1 cortical | 4121/6024 | 3893/9888 | 2124/4031 | 1624/4004 |
| DKK1 cortical % | 69 ± 6 | 39 ± 6 *** | 52 ± 6 | 42 ± 9 * |
| DKK 1 trabecular | 2357/3173 | 1312/3838 | 775/1614 | 738/2147 |
| DKK 1 trabecular % | 75 ± 7 | 35 ± 18 ** | 49 ± 20 | 36 ± 12 |
| SOST cortical | 1257/2620 | 702/2347 | 1170/2572 | 862/2337 |
| SOST cortical % | 49 ± 9 | 30 ± 8 * | 45 ± 8 | 39 ± 12 |
| PSTN cortical | 789/3702 | 1789/4160 | 858/3455 | 1939/3622 |
| PSTN cortical % | 21 ± 3 | 43 ± 2 *** | 24 ± 6 | 54 ± 5 *** |
| PSTN trabecular | 327/1653 | 645/1779 | 327/1688 | 926/1691 |
| PSTN trabecular % | 20 ± 2 | 39 ± 8 *** | 20 ± 9 | 55 ± 9 ** |
| FGF23 cortical | 1182/2307 | 1289/2387 | 1010/2462 | 1344/2430 |
| FGF23 cortical % | 54 ± 16 | 54 ± 5 | 42 ± 13 | 55 ± 8 |
| OPG cortical | 1938/3443 | 2758/5265 | 1888/3598 | 1681/3051 |
| OPG cortical % | 58.88 ± 8.77 | 54.66 ± 14.04 | 53.51 ± 7.88 | 55.73 ± 10.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerschan-Schindl, K.; Schramek, V.; Butylina, M.; Föger-Samwald, U.; Pietschmann, P. Differential Expression of Dickkopf 1 and Periostin in Mouse Strains with High and Low Bone Mass. Biology 2022, 11, 1840. https://doi.org/10.3390/biology11121840
Kerschan-Schindl K, Schramek V, Butylina M, Föger-Samwald U, Pietschmann P. Differential Expression of Dickkopf 1 and Periostin in Mouse Strains with High and Low Bone Mass. Biology. 2022; 11(12):1840. https://doi.org/10.3390/biology11121840
Chicago/Turabian StyleKerschan-Schindl, Katharina, Victoria Schramek, Maria Butylina, Ursula Föger-Samwald, and Peter Pietschmann. 2022. "Differential Expression of Dickkopf 1 and Periostin in Mouse Strains with High and Low Bone Mass" Biology 11, no. 12: 1840. https://doi.org/10.3390/biology11121840