
In Silico Study of Mangostin Compounds and Its Derivatives as Inhibitors of α-Glucosidase Enzymes for Anti-Diabetic Studies

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ligand Creation
2.2. Molecular Docking
2.3. Molecular Dynamics Simulation
3. Results
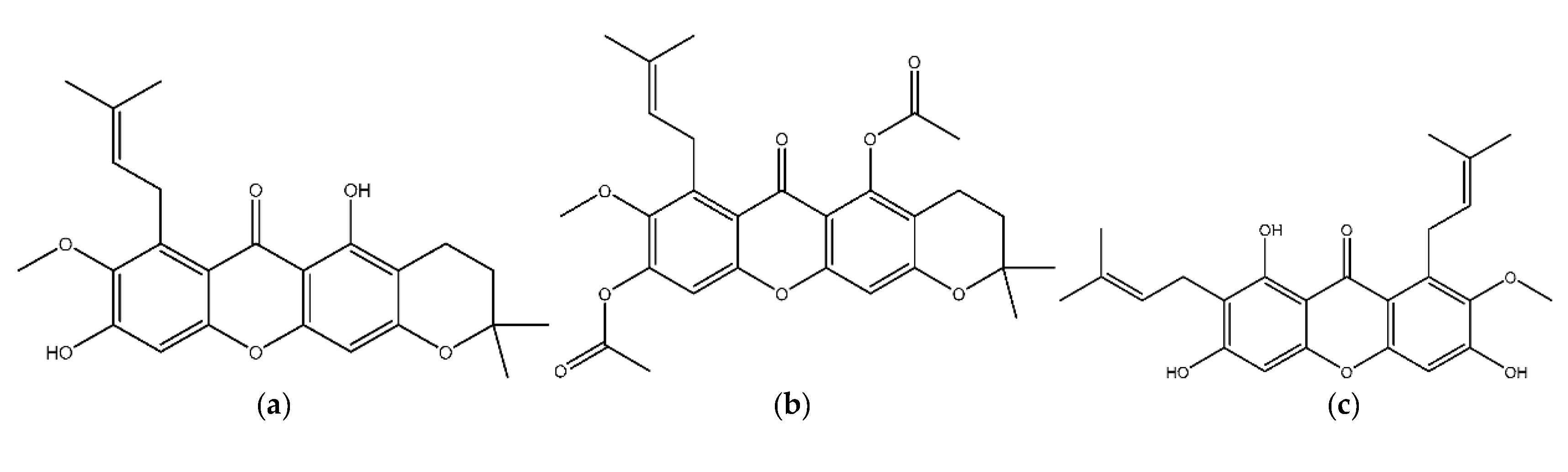
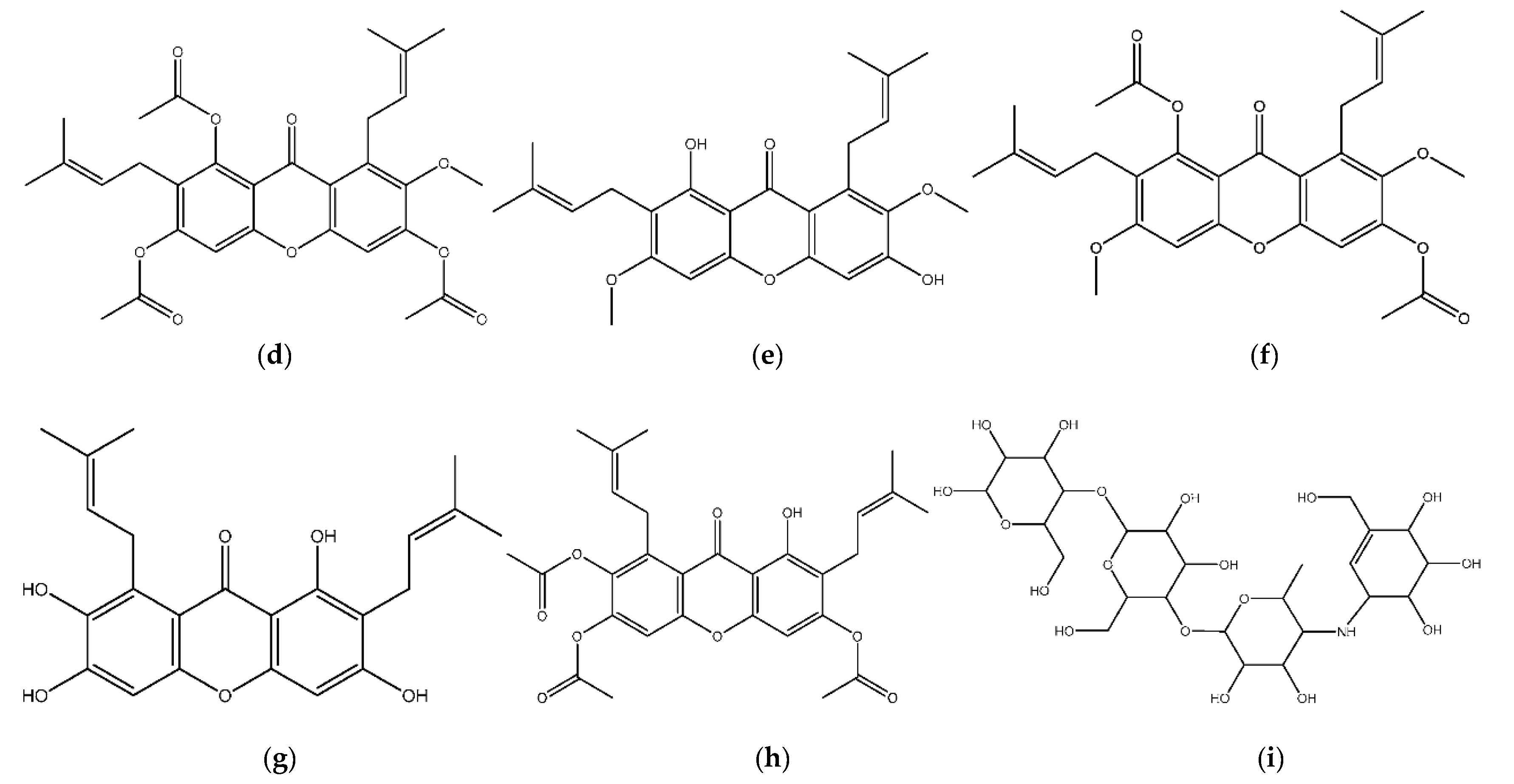
3.1. Structure Preparation
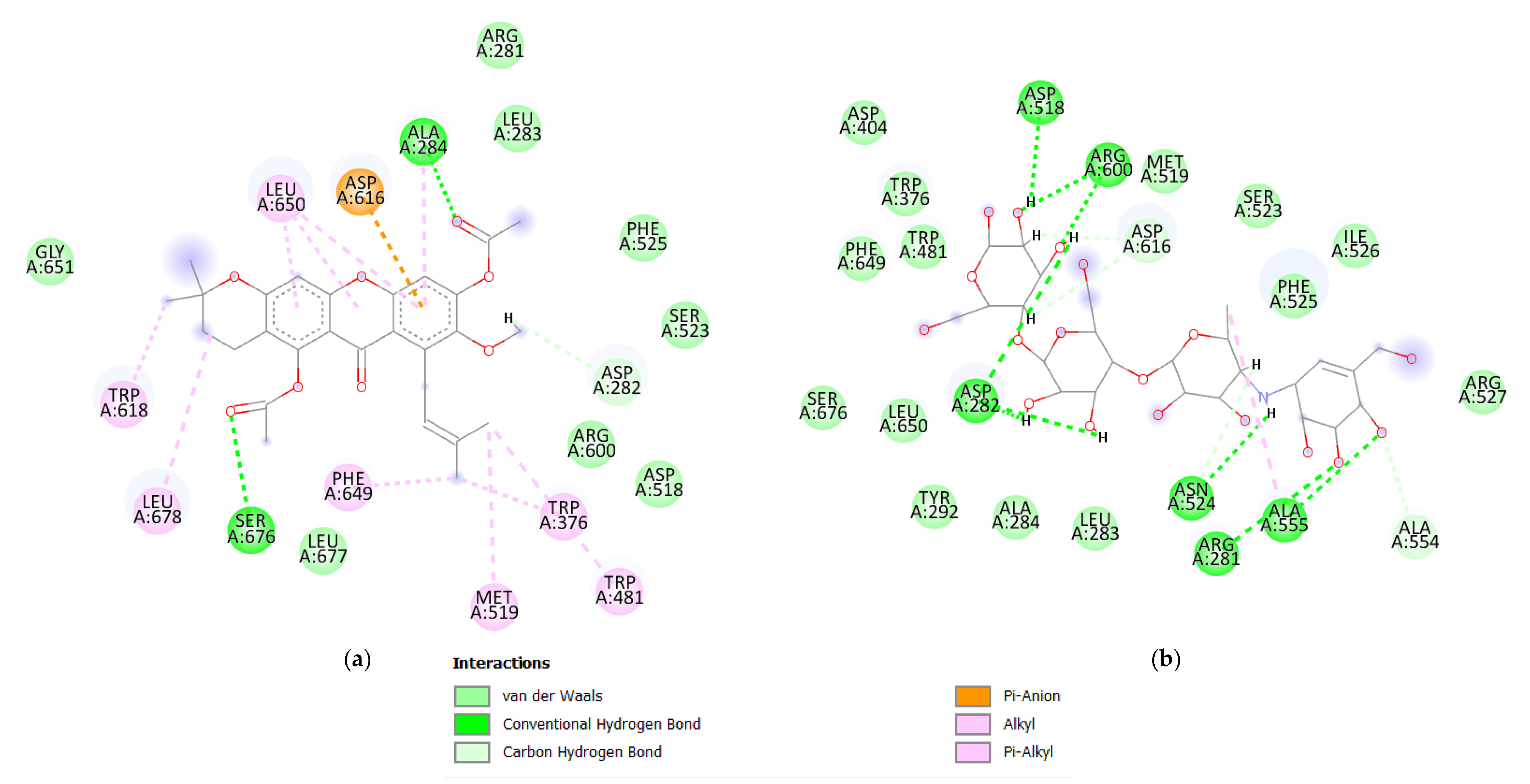
3.2. Molecular Docking
3.3. Molecular Dynamics Simulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandra, R.A.I.; Sriwidodo; Diantini, A.; Maksum, I.P. Restriction Enzymes ApaI Analysis to Find A3243G Mutation in Indonesia Diabetes Mellitus Type II Patients. J. Med. Bioeng. 2015, 4, 492–496. [Google Scholar] [CrossRef] [Green Version]
- IDF. IDF Diabetes Atlas, 10th ed.; IDF: Brussels, Belgium, 2021. [Google Scholar]
- Yani, F.; Bellatasie, R.; Fauziah, F. Antidiabetic Potential of G. Mangostana Extract and α -Mangostin Compounds from Mangosteen (Garcinia mangostana Linn.). EAS J. Pharm. Pharmacol. 2021, 3, 94–105. [Google Scholar] [CrossRef]
- McIver, L.A.; Charles, P.; Tripp, J. Acarbose; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Mahmudah, R.; Adnyana, I.; Sukandar, E. Molecular Docking Studies of α-Mangostin, γ-Mangostin, and Xanthone on Peroxisome Proliferator-Activated Receptor Gamma Diphenyl Peptidase-4 Enzyme, and Aldose Reductase Enzyme as an Antidiabetic Drug Candidate. J. Adv. Pharm. Technol. Res. 2021, 12, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Simamora, A.; Santoso, A.W.; Timotius, K.H. α-Glucosidase Inhibitory Effect of Fermented Fruit Juice of Morinda Citrifolia l and Combination Effect with Acarbose. Curr. Res. Nutr. Food Sci. 2019, 7, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Djeujo, F.M.; Francesconi, V.; Gonella, M.; Ragazzi, E.; Tonelli, M.; Froldi, G. Anti-α-Glucosidase and Antiglycation Activities of α-Mangostin and New Xanthenone Derivatives: Enzymatic Kinetics and Mechanistic Insights through in Vitro Studies. Molecules 2022, 27, 547. [Google Scholar] [CrossRef]
- Chen, S.P.; Lin, S.R.; Chen, T.H.; Ng, H.S.; Yim, H.S.; Leong, M.K.; Weng, C.F. Mangosteen Xanthone γ-Mangostin Exerts Lowering Blood Glucose Effect with Potentiating Insulin Sensitivity through the Mediation of AMPK/PPARγ. Biomed. Pharmacother. 2021, 144, 112333. [Google Scholar] [CrossRef]
- Kodiha, M.; Rassi, J.G.; Brown, C.M.; Stochaj, U. Localization of AMP Kinase Is Regulated by Stress, Cell Density, and Signaling through the MEK→ERK1/2 Pathway. Am. J. Physiol.-Cell Physiol. 2007, 293, 1427–1436. [Google Scholar] [CrossRef]
- Tyagi, S.; Gupta, P.; Saini, A.; Kaushal, C.; Sharma, S. The Peroxisome Proliferator-Activated Receptor: A Family of Nuclear Receptors Role in Various Diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236–240. [Google Scholar] [CrossRef]
- Smith, M.E.; Dion, G. Morton the mouth, salivary glands and oesophagus. In The Digestive System; Churchill Livingstone Elsevier: Edinburgh, UK, 2010; pp. 19–38. [Google Scholar]
- Tomasik, P.; Horton, D. Enzymatic Conversions of Starch, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; Volume 68, ISBN 9780123965233. [Google Scholar]
- Prabhakar, P.K.; Doble, M. Mechanism of Action of Natural Products Used in the Treatment of Diabetes Mellitus. Chin. J. Integr. Med. 2011, 17, 563–574. [Google Scholar] [CrossRef]
- Husen, S.A.; Salamun; Ansori, A.N.M.; Susilo, R.J.K.; Hayaza, S.; Winarni, D. The Effect of Alpha-Mangostin in Glucose Level, Cholesterol Level, and Diameter of the Islets of Langerhans of STZ-Induced Diabetic Mice. In Proceedings of the 2nd International Conference Postgraduate School (ICPS 2018), Surabaya, Indonesia, 10–11 July 2018; pp. 561–566. [Google Scholar] [CrossRef]
- Tholib Baladraf, T. Effect Jamblang Stem Barks, Mangosteen Peel, and Cinnamon to Blood Sugar Levels. J. La Lifesci 2021, 2, 29–36. [Google Scholar] [CrossRef]
- Rocky, E.; Rafsanjani, M.; Fadlan, A.; Ersam, T. Acetylation of γ-Mangostin Isolated from the Mangosteen Pericarp (Garcinia mangostana Linn.) and Their Antidiabetic Activity. IPTEK J. Proc. Ser. 2020, 127–131. [Google Scholar] [CrossRef]
- Prasetyanti, I.K.; Sukardiman. Suharjono Admet Prediction and in Silico Analysis of Mangostin Derivatives and Sinensetin on Maltase-Glucoamylase Target for Searching Anti-Diabetes Drug Candidates. Pharmacogn. J. 2021, 13, 883–889. [Google Scholar] [CrossRef]
- Roig-Zamboni, V.; Cobucci-Ponzano, B.; Iacono, R.; Ferrara, M.C.; Germany, S.; Bourne, Y.; Parenti, G.; Moracci, M.; Sulzenbacher, G. Structure of Human Lysosomal Acid α-Glucosidase-A Guide for the Treatment of Pompe Disease. Nat. Commun. 2017, 8, 1111. [Google Scholar] [CrossRef] [Green Version]
- Moretti, S. In Silico Experiments in Scientific Papers on Molecular Biology. Sci. Stud. 2011, 24, 23–42. [Google Scholar] [CrossRef]
- Valerio, L.G. Application of Advanced in Silico Methods for Predictive Modeling and Information Integration. Expert Opin. Drug Metab. Toxicol. 2012, 8, 395–398. [Google Scholar] [CrossRef]
- Ingólfsson, H.I.; Arnarez, C.; Periole, X.; Marrink, S.J. Computational “microscopy” of Cellular Membranes. J. Cell Sci. 2016, 129, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Zloh, M.; Kirton, S.B. The Benefits of in Silico Modeling to Identify Possible Small-Molecule Drugs and Their off-Target Interactions. Future Med. Chem. 2018, 10, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Farooq, F.T.; Holcik, M.; MacKenzie, A. Spinal Muscular Atrophy: Classification, Diagnosis, Background, Molecular Mechanism and Development of Therapeutics; IntechOpen: London, UK, 2013; pp. 561–579. [Google Scholar] [CrossRef] [Green Version]
- Carlomagno, N.; Incollingo, P.; Tammaro, V.; Peluso, G.; Rupealta, N.; Chiacchio, G.; Sandoval Sotelo, M.L.; Minieri, G.; Pisani, A.; Riccio, E.; et al. Diagnostic, Predictive, Prognostic, and Therapeutic Molecular Biomarkers in Third Millennium: A Breakthrough in Gastric Cancer. BioMed Res. Int. 2017, 2017, 7869802. [Google Scholar] [CrossRef]
- Johnsen, J.I.; Dyberg, C.; Fransson, S.; Wickström, M. Molecular Mechanisms and Therapeutic Targets in Neuroblastoma. Pharmacol. Res. 2018, 131, 164–176. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Baghdadi, A.; Tayebi-Meigooni, A. Alpha-Mangostin-Rich Extracts from Mangosteen Pericarp: Optimization of Green Extraction Protocol and Evaluation of Biological Activity. Molecules 2018, 23, 1852. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.M.M.; Proença, C.; Freitas, M.; Araújo, A.N.; Silva, A.M.S.; Fernandes, E. Inhibition of the Carbohydrate-Hydrolyzing Enzymes α-Amylase and α-Glucosidase by Hydroxylated Xanthones. Food Funct. 2022, 13, 7930–7941. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D.; Earnshaw, W.C.; Lippincott-Schwartz, J.; Johnson, G.T. Biophysical Principles; Elsevier: Philadelphia, PA, USA, 2017; pp. 53–62. [Google Scholar]
- Zhou, H.X.; Pang, X. Electrostatic Interactions in Protein Structure, Folding, Binding, and Condensation. Chem. Rev. 2018, 118, 1691–1741. [Google Scholar] [CrossRef] [PubMed]
- Proença, C.; Ribeiro, D.; Freitas, M.; Carvalho, F.; Fernandes, E. A Comprehensive Review on the Antidiabetic Activity of Flavonoids Targeting PTP1B and DPP-4: A Structure-Activity Relationship Analysis. Crit. Rev. Food Sci. Nutr. 2022, 62, 4095–4151. [Google Scholar] [CrossRef] [PubMed]
- Ngawhirunpat, T.; Opanasopi, P.; Sukma, M.; Sittisombut, C.; Kat, A.; Adachi, I. Antioxidant, Free Radical-Scavenging Activity and Cytotoxicity of Different Solvent Extracts and Their Phenolic Constituents from the Fruit Hull of Mangosteen (Garcinia mangostana). Pharm. Biol. 2010, 48, 55–62. [Google Scholar] [CrossRef]
- Destiarani, W.; Mulyani, R.; Yusuf, M.; Maksum, I.P. Molecular Dynamics Simulation of T10609C and C10676G Mutations of Mitochondrial ND4L Gene Associated with Proton Translocation in Type 2 Diabetes Mellitus and Cataract Patients. Bioinform. Biol. Insights 2020, 14, 1177932220978672. [Google Scholar] [CrossRef]
- Maksum, I.P.; Maulana, A.F.; Yusuf, M.; Mulyani, R.; Destiarani, W.; Rustaman, R. Molecular Dynamics Simulation of a TRNA-Leucine Dimer with an A3243G Heteroplasmy Mutation in Human Mitochondria Using a Secondary Structure Prediction Approach. Indones. J. Chem. 2022, 22, 1043–1051. [Google Scholar] [CrossRef]
- Maksum, I.P.; Saputra, S.R.; Indrayati, N.; Yusuf, M.; Subroto, T. Bioinformatics Study of m.9053G>A Mutation at the ATP6 Gene in Relation to Type 2 Diabetes Mellitus and Cataract Diseases. Bioinform. Biol. Insights 2017, 11, 1–5. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No | Compound Name | Total Energy (kcal/mol) |
|---|---|---|
| 1 | α-Mangostin | 25.2737 |
| 2 | 1,3,6-trimethyl-ester-α-mangostin | 57.8246 |
| 3 | β-Mangostin | 32.7636 |
| 4 | 1,6-dimethyl-ester-β-mangostin | 55.602 |
| 5 | 3-Isomangostin | 34.796 |
| 6 | 1,6-dimethyl-ester-3-isomangostin | 65.6732 |
| 7 | γ-Mangostin | 17.1138 |
| 8 | 1,3,6-tetramethyl-ester-γ-mangostin | 54.6131 |
| 9 | Acarbose | 50.9353 |
| Compound | Affinity (kcal/mol) |
|---|---|
| Acarbose | −7.1 |
| α-Mangostin | −7.3 |
| 1,3,6-trimethyl-ester-α-mangostin | −7.2 |
| β-Mangostin | −7.4 |
| 1,6-dimethyl-ester-β-mangostin | −6.7 |
| γ-Mangostin | −7.7 |
| 3,6,7-trimethyl-ester-γ-mangostin | −7.8 |
| 3-Isomangostin | −8.1 |
| 1,6-dimethyl-ester-3-isomangostin | −8.2 |
| No | Molecule | Receptor (Å) | Ligand (Å) |
|---|---|---|---|
| 1 | Acarbose | 1.98 | 2.24 |
| 2 | α-Mangostin | 1.7 | 1.61 |
| 3 | 1,3,6-trimethyl-ester-α-mangostin | 2.1 | 2.86 |
| 4 | β-Mangostin | 1.53 | 1.59 |
| 5 | 1,6-dimethyl-ester-β-mangostin | 1.7 | 1.42 |
| 6 | γ-Mangostin | 1.7 | 1.34 |
| 7 | 3,6,7-trimethyl-ester-γ-mangostin | 1.99 | 1.53 |
| 8 | 3-Isomangostin | 1.75 | 1.38 |
| 9 | 1,6-dimethyl-ester-3-isomangostin | 2.01 | 1.18 |
| No | Molecule | Energy Component(Kcal/mol) | ||||||
|---|---|---|---|---|---|---|---|---|
| VDWaals | EEL | EGB | Esurf | ▲G (Gas) | ▲G (Solv) | ▲G (Total) | ||
| 1 | Acarbose | −22.8293 | −61.6643 | 74.415 | −4.3124 | −84.4936 | 70.0981 | −14.3955 |
| 2 | α-Mangostin | −23.6659 | −4.21 | 15.6258 | −2.2922 | −27.8759 | 12.6636 | −15.2123 |
| 3 | 1,3,6-trimethyl-ester-α-mangostin | −26.7758 | −4.7378 | 16.4819 | −3.0714 | −31.5136 | 13.4645 | −18.0491 |
| 4 | β-Mangostin | −22.6442 | −2.0799 | 11.9138 | −2.5386 | −24.7241 | 9.3752 | −15.3489 |
| 5 | 1,6-dimethyl-ester-β-mangostin | −30.4346 | −8.7252 | 23.3196 | −3.3416 | −39.1597 | 19.978 | −19.1817 |
| 6 | γ-Mangostin | −25.2716 | −38.8053 | 46.8243 | −3.743 | −64.0769 | 43.0813 | −20.9956 |
| 7 | 3,6,7-trimethyl-ester-γ-mangostin | −25.2606 | −17.0004 | 30.7017 | −3.1499 | −42.2611 | 27.5518 | −14.7093 |
| 8 | 3-Isomangostin | −20.3336 | −2.3135 | 12.1838 | −2.4471 | −22.6471 | 9.7367 | −12.9104 |
| 9 | 1,6-dimethyl-ester-3-isomangostin | −25.679 | −7.2242 | 17.9591 | −2.7725 | −32.9032 | 15.1866 | −17.7166 |
| No | Molecule | Hydrogen Bond |
|---|---|---|
| 1 | Acarbose | ASP518, ASP616, ASP282, ARG600 |
| 2 | α-Mangostin | NA |
| 3 | 1,3,6-trimethyl-ester-α-mangostin | ASN652 |
| 4 | β-Mangostin | NA |
| 5 | 1,6-dimethyl-ester-β-mangostin | NA |
| 6 | γ-Mangostin | ASP404, ASP518 |
| 7 | 3,6,7-trimethyl-ester-γ-mangostin | NA |
| 8 | 3-Isomangostin | NA |
| 9 | 1,6-dimethyl-ester-3-isomangostin | TRP618 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maulana, A.F.; Sriwidodo, S.; Rukayadi, Y.; Maksum, I.P. In Silico Study of Mangostin Compounds and Its Derivatives as Inhibitors of α-Glucosidase Enzymes for Anti-Diabetic Studies. Biology 2022, 11, 1837. https://doi.org/10.3390/biology11121837
Maulana AF, Sriwidodo S, Rukayadi Y, Maksum IP. In Silico Study of Mangostin Compounds and Its Derivatives as Inhibitors of α-Glucosidase Enzymes for Anti-Diabetic Studies. Biology. 2022; 11(12):1837. https://doi.org/10.3390/biology11121837
Chicago/Turabian StyleMaulana, Ahmad Fariz, Sriwidodo Sriwidodo, Yaya Rukayadi, and Iman Permana Maksum. 2022. "In Silico Study of Mangostin Compounds and Its Derivatives as Inhibitors of α-Glucosidase Enzymes for Anti-Diabetic Studies" Biology 11, no. 12: 1837. https://doi.org/10.3390/biology11121837