
Ldha-Dependent Metabolic Programs in Sertoli Cells Regulate Spermiogenesis in Mouse Testis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Histological, Immunofluorescent, and Apoptosis Analyses
2.3. Sertoli Cell Isolation and Quantitative RT-PCR
2.4. Sperm Quality and Ultrastructural Analyses(TEM)
2.5. Unlabeled Targeted Metabolomics
2.6. Lactate Concertation Measurement
2.7. Choline Supplementation
2.8. Sperm Membrane Staining and Acrosomal Reaction (AR) Analysis
2.9. In Vitro Fertilization and Embryo Culture
2.10. Western Blot
2.11. Statistical Analysis
3. Results
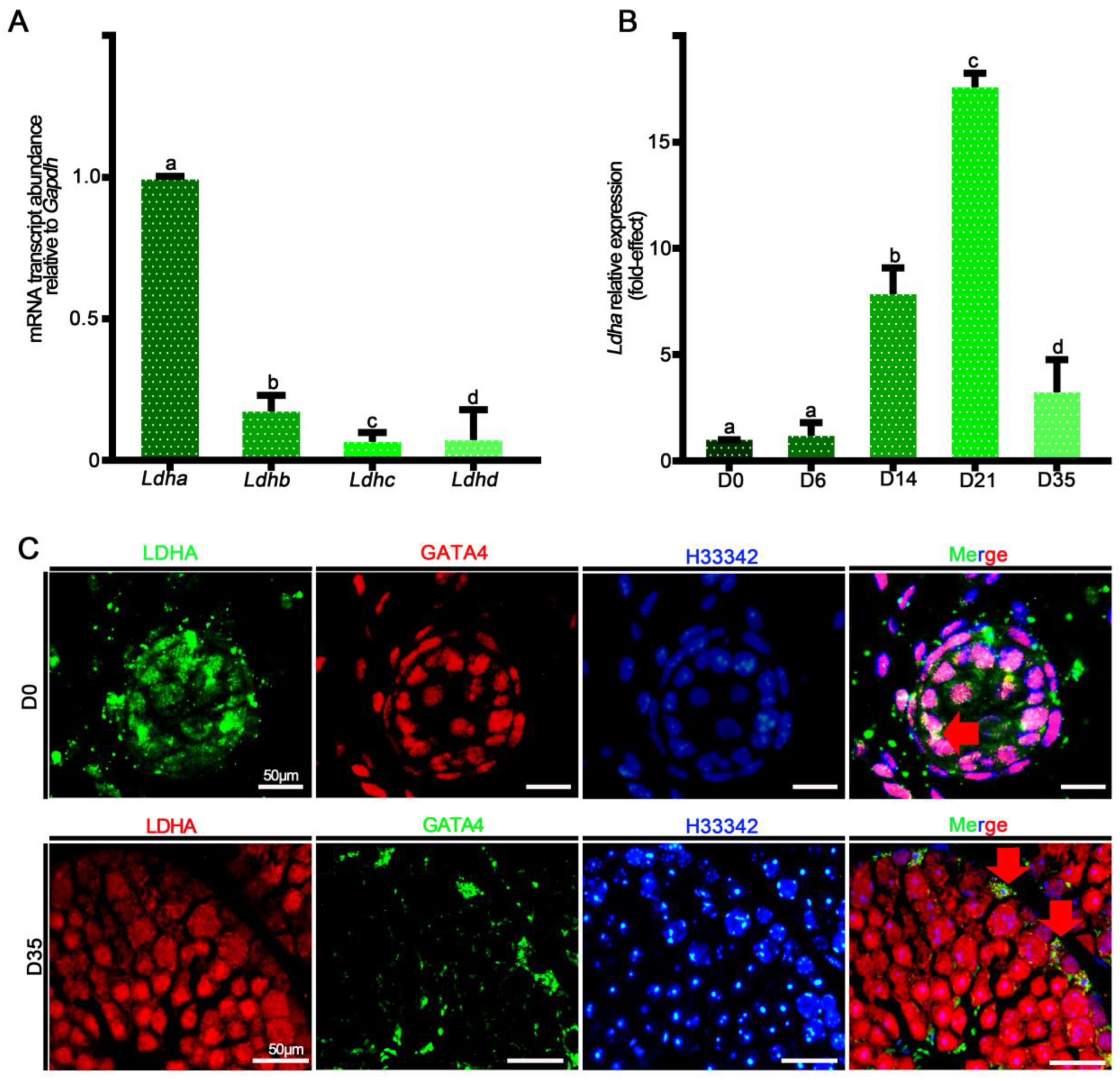
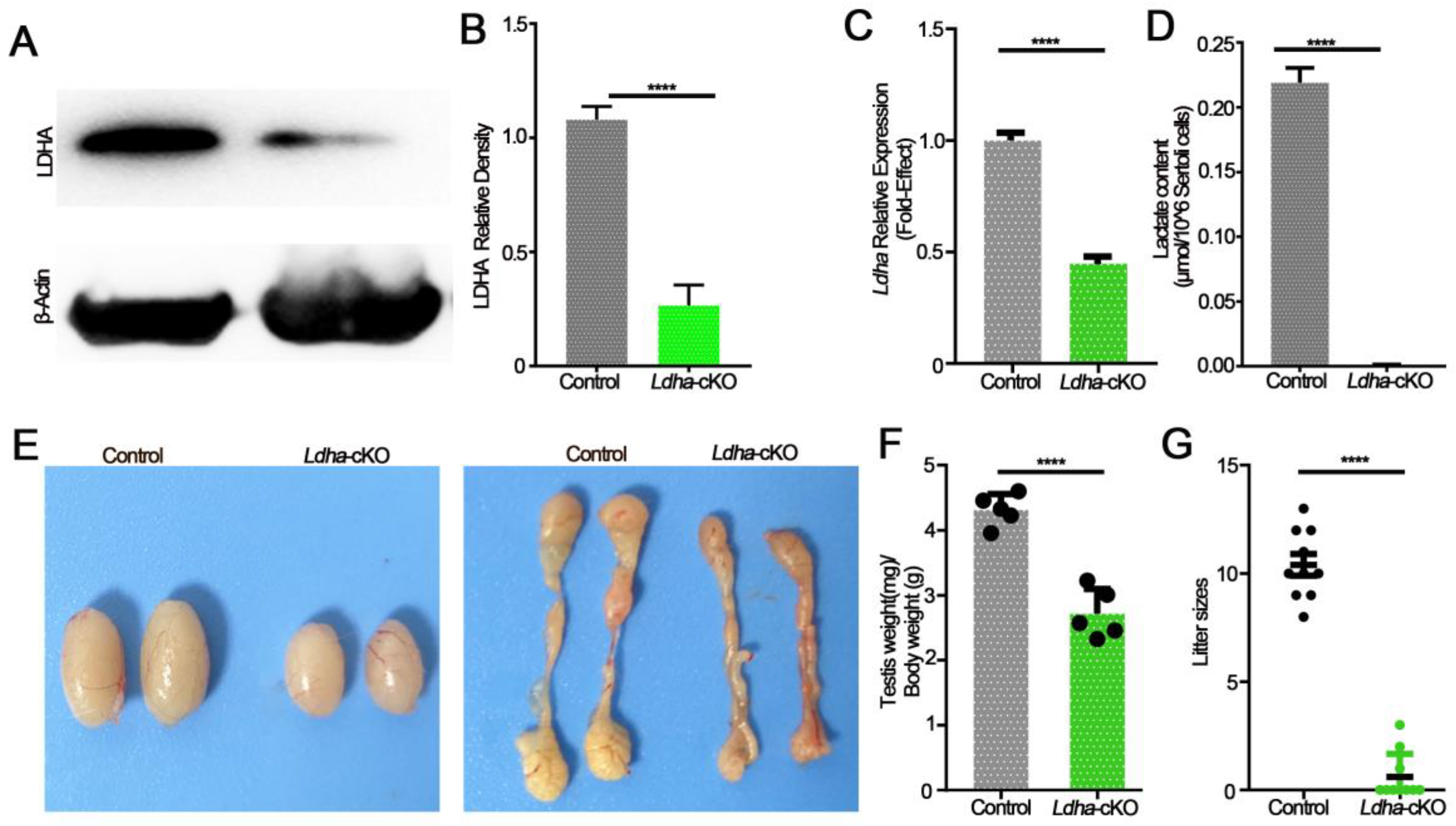
3.1. Ldha Expression Was Enriched in Sertoli Cells, and Conditional Deletion of Lhda Significantly Impacted Fertility
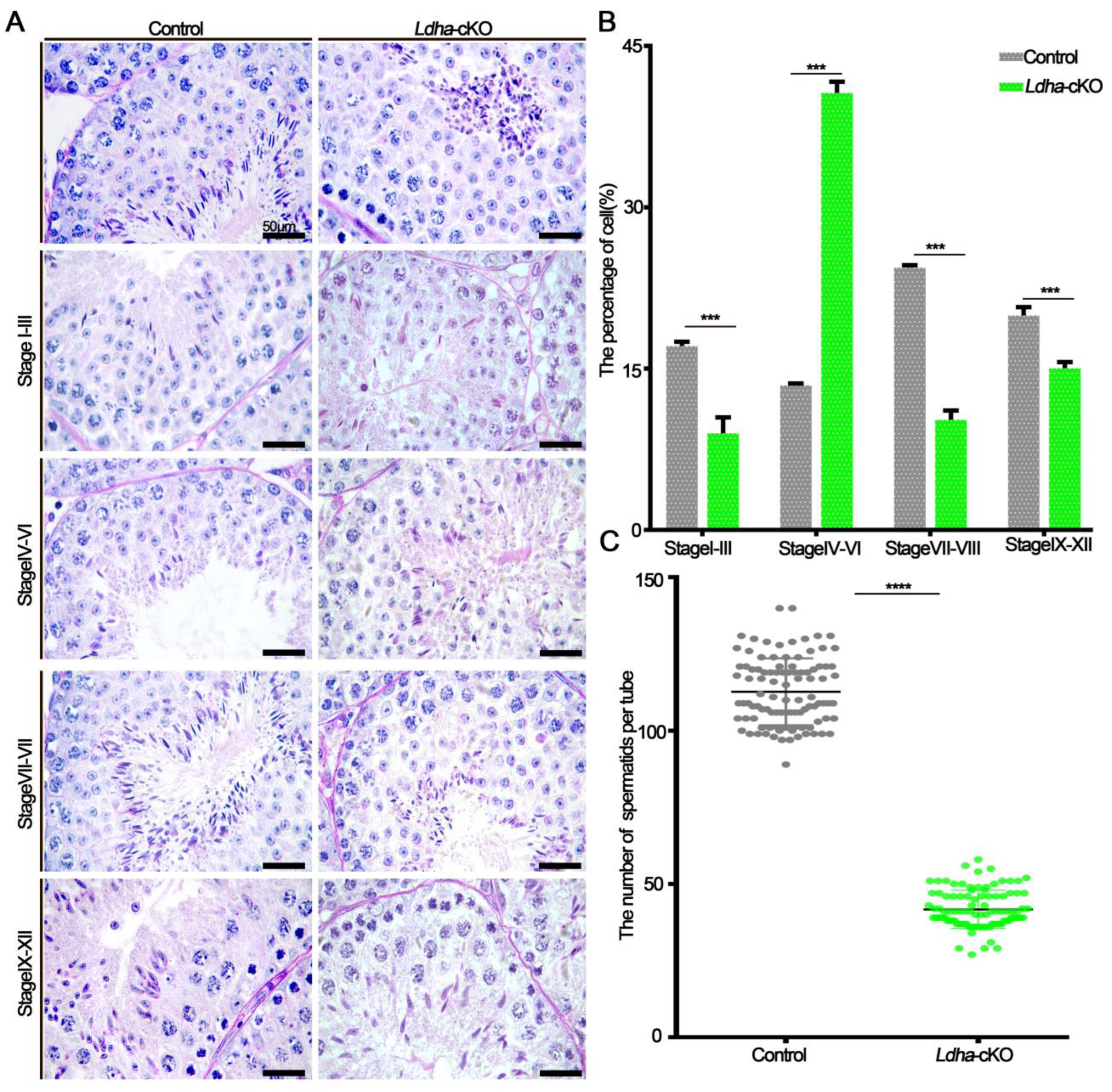
3.2. Ldha Loss of Function in Sertoli Cells Resulted in Defects in Spermiogenesis
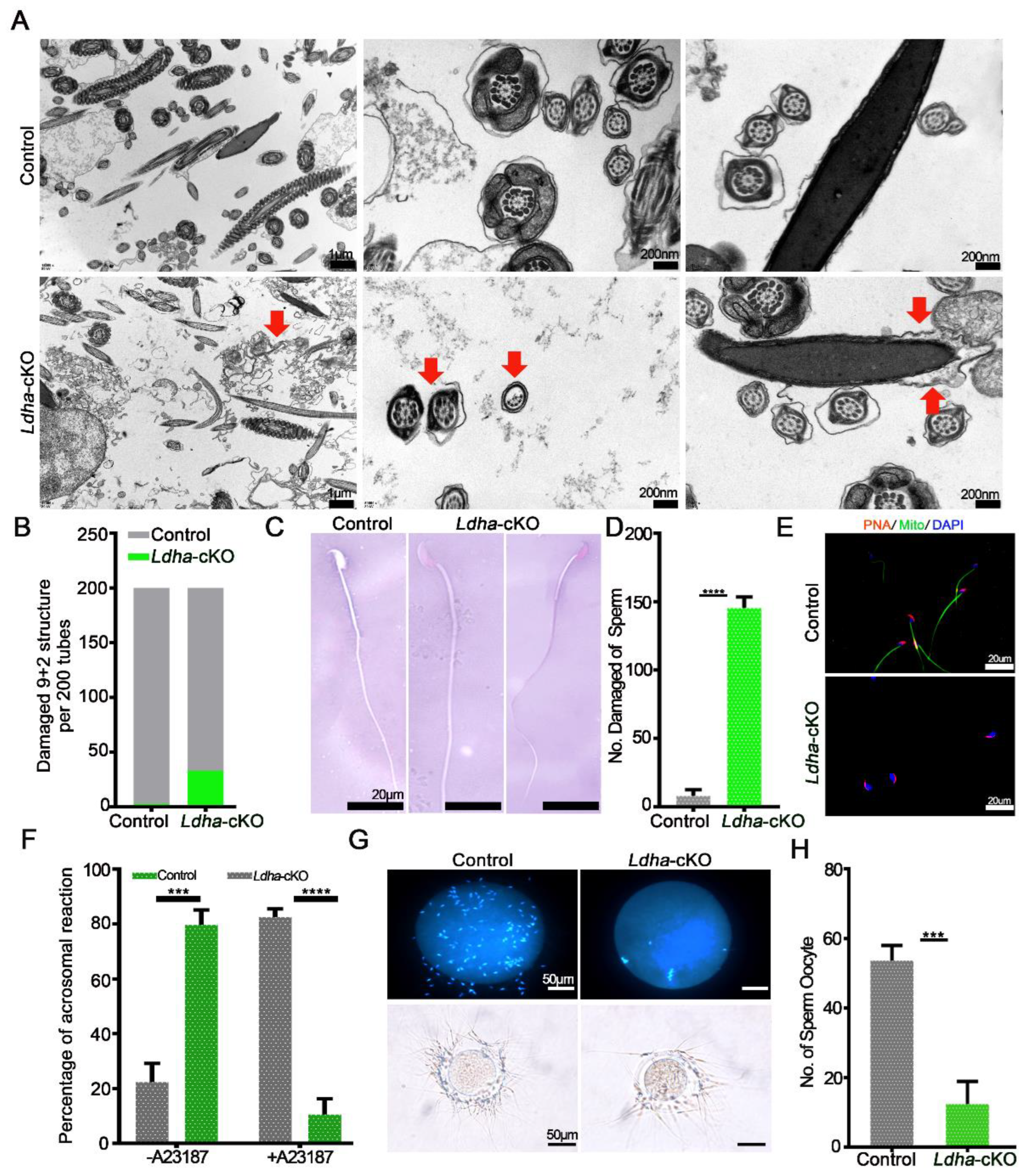
3.3. Ldha Deletion in Sperm Affected Plasma Membrane Integrity and Function
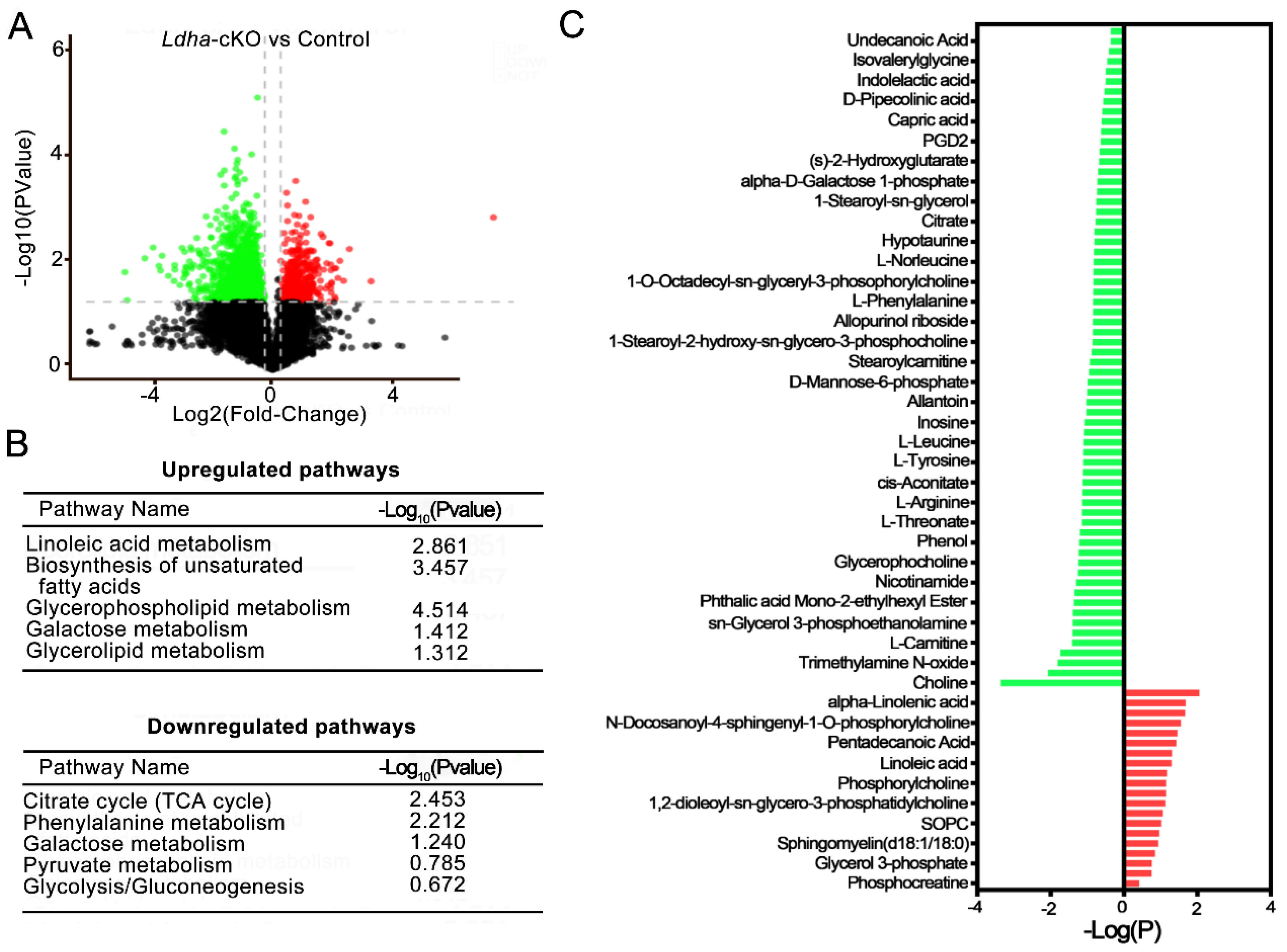
3.4. Ldha Deletion in Sertoli Cells Changed the Metabolic Profile of Sperm
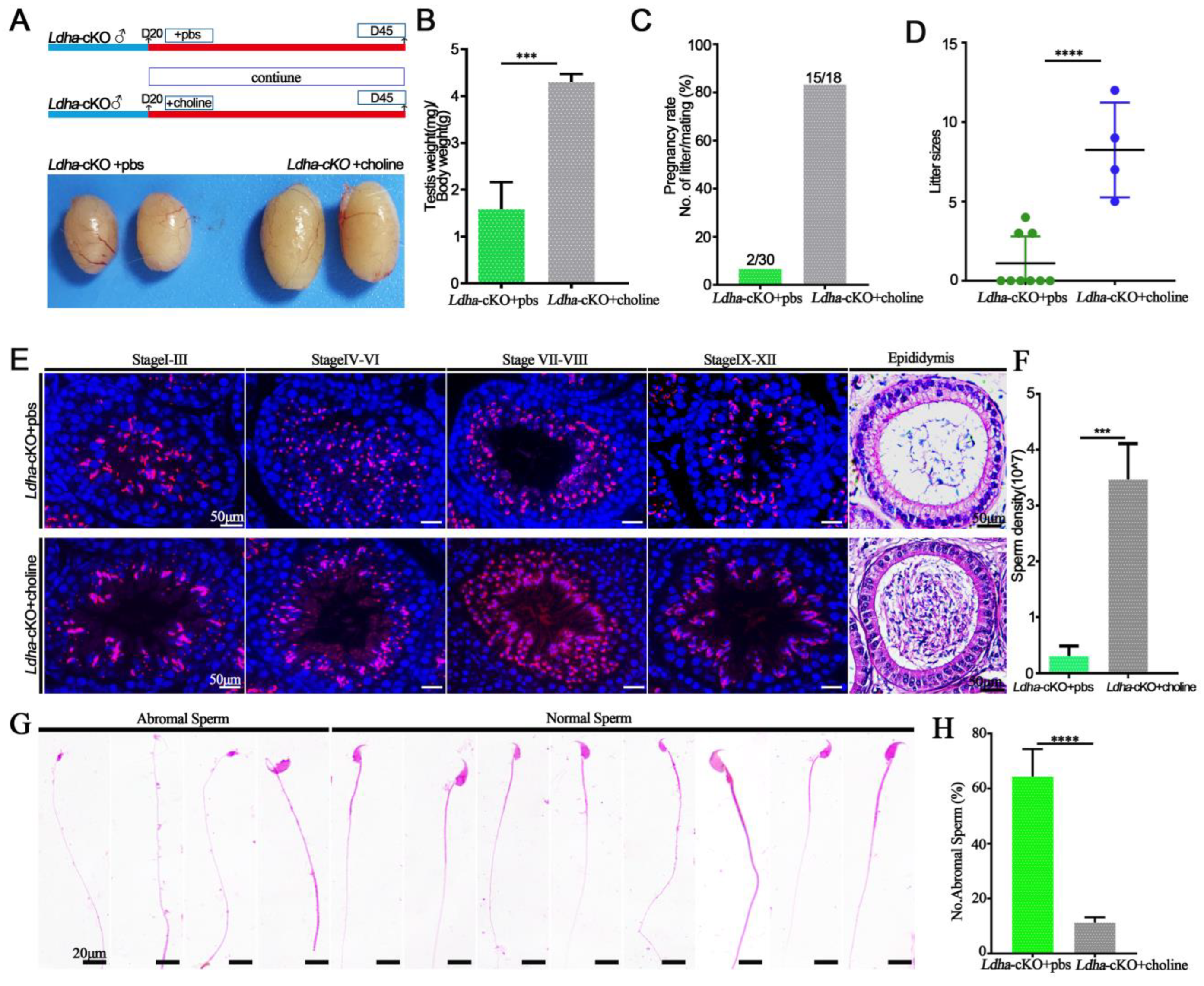
3.5. Choline Supplementation Rescued Spermatogenic Defects in Ldha-cKO Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- França, L.R.; Hess, R.A.; Dufour, J.M.; Hofmann, M.-C.; Griswold, M.D. The Sertoli cell: One hundred fifty years of beauty and plasticity. Andrology 2016, 4, 189–212. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, T.; Orwig, K.; Avarbock, M.R.; Brinster, R.L. Restoration of Spermatogenesis in Infertile Mice by Sertoli Cell Transplantation1. Biol. Reprod. 2003, 68, 1064–1071. [Google Scholar] [CrossRef] [Green Version]
- Endo, T.; Freinkman, E.; de Rooij, D.G.; Page, D.C. Periodic production of retinoic acid by meiotic and somatic cells coordinates four transitions in mouse spermatogenesis. Proc. Natl. Acad. Sci. USA 2017, 114, E10132–E10141. [Google Scholar] [CrossRef] [Green Version]
- Larose, H.; Kent, T.; Ma, Q.; Shami, A.N.; Harerimana, N.; Li, J.Z.; Hammoud, S.S.; Handel, M.A. Regulation of meiotic progression by Sertoli-cell androgen signaling. Mol. Biol. Cell 2020, 31, 2841–2862. [Google Scholar] [CrossRef] [PubMed]
- Holdcraft, R.W.; Braun, R.E. Androgen receptor function is required in Sertoli cells for the terminal differentiation of haploid spermatids. Development 2004, 131, 459–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebourcet, D.; Darbey, A.; Monteiro, A.; Soffientini, U.; Tsai, Y.T.; Handel, I.; Pitetti, J.-L.; Nef, S.; Smith, L.B.; O’Shaughnessy, P.J. Sertoli Cell Number Defines and Predicts Germ and Leydig Cell Population Sizes in the Adult Mouse Testis. Endocrinology 2017, 158, 2955–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, J.; Heinrich, J.; Brehm, R. Blood–testis barrier and Sertoli cell function: Lessons from SCCx43KO mice. Reproduction 2016, 151, R15–R27. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, R.; Trindade, F.; Vitorino, R. Proteome Profiling of Sertoli Cells Using a GeLC-MS/MS Strategy. Methods Mol. Biol. 2018, 1748, 173–190. [Google Scholar] [CrossRef]
- Boussouar, F.; Benahmed, M. Lactate and energy metabolism in male germ cells. Trends Endocrinol. Metab. 2004, 15, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Tatapudy, S.; Aloisio, F.; Barber, D.; Nystul, T. Cell fate decisions: Emerging roles for metabolic signals and cell morphology. EMBO Rep. 2017, 18, 2105–2118. [Google Scholar] [CrossRef]
- Rogatzki, M.J.; Ferguson, B.S.; Goodwin, M.L.; Gladden, L.B. Lactate is always the end product of glycolysis. Front. Neurosci. 2015, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valvona, C.J.; Fillmore, H.L.; Nunn, P.B.; Pilkington, G.J. The Regulation and Function of Lactate Dehydrogenase A: Therapeutic Potential in Brain Tumor. Brain Pathol. 2016, 26, 3–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monroe, G.R.; van Eerde, A.M.; Tessadori, F.; Duran, K.J.; Savelberg, S.M.C.; van Alfen, J.C.; Terhal, P.A.; van der Crabben, S.N.; Lichtenbelt, K.D.; Fuchs, S.A.; et al. Identification of human D lactate dehydrogenase deficiency. Nat. Commun. 2019, 10, 1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauchi, S.; Rauch, M.C.; Alfaro, I.E.; Cea, C.; Concha, I.I.; Benos, D.J.; Reyes, J.G. Kinetics, molecular basis, and differentiation ofl-lactate transport in spermatogenic cells. Am. J. Physiol. Physiol. 2005, 288, C523–C534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajpai, M.; Gupta, G.; Setty, B.S. Changes in carbohydrate metabolism of testicular germ cells during meiosis in the rat. Eur. J. Endocrinol. 1998, 138, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Jutte, N.H.P.M.; Jansen, R.; Grootegoed, J.A.; Rommerts, F.F.G.; Clausen, O.P.F.; Van Der Molen, H.J. Regulation of survival of rat pachytene spermatocytes by lactate supply from Sertoli cells. Reproduction 1982, 65, 431–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jutte, N.H.P.M.; Grootegoed, J.A.; Rommerts, F.F.G.; Van Der Molen, H.J. Exogenous lactate is essential for metabolic activities in isolated rat spermatocytes and spermatids. Reproduction 1981, 62, 399–405. [Google Scholar] [CrossRef]
- Thomas, K.; Del Mazo, J.; Eversole, P.; Bellve, A.; Hiraoka, Y.; Li, S.; Simon, M. Developmental regulation of expression of the lactate dehydrogenase (LDH) multigene family during mouse spermatogenesis. Development 1990, 109, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Odet, F.; Duan, C.; Willis, W.D.; Goulding, E.H.; Kung, A.; Eddy, E.M.; Goldberg, E. Expression of the Gene for Mouse Lactate Dehydrogenase C (Ldhc) Is Required for Male Fertility1. Biol. Reprod. 2008, 79, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Odet, F.; Gabel, S.A.; Williams, J.; London, R.E.; Goldberg, E.; Eddy, E.M. Lactate Dehydrogenase C and Energy Metabolism in Mouse Sperm. Biol. Reprod. 2011, 85, 556–564. [Google Scholar] [CrossRef]
- Tang, H.; Duan, C.; Bleher, R.; Goldberg, E. Human Lactate Dehydrogenase A (LDHA) Rescues Mouse Ldhc-Null Sperm Function1. Biol. Reprod. 2013, 88, 96. [Google Scholar] [CrossRef] [PubMed]
- Koiri, R.K.; Rawat, D.; Chonker, S.K.; Naik, R.A.; Mehrotra, A.; Trigun, S.K. Lactate as a signaling molecule Journey from dead end product of glycolysis to tumor survival. Front. Biosci. 2019, 24, 366–381. [Google Scholar] [CrossRef] [PubMed]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, P.; Shanmugam, A.; Swafford, D.; Suryawanshi, A.; Bhattacharjee, P.; Hussein, M.S.; Koni, P.A.; Prasad, P.D.; Kurago, Z.B.; Thangaraju, M.; et al. GPR81, a Cell-Surface Receptor for Lactate, Regulates Intestinal Homeostasis and Protects Mice from Experimental Colitis. J. Immunol. 2018, 200, 1781–1789. [Google Scholar] [CrossRef] [Green Version]
- Chang, A.J.; Ortega, F.E.; Riegler, J.; Madison, D.V.; Krasnow, M.A. Oxygen regulation of breathing through an olfactory receptor activated by lactate. Nature 2015, 527, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griswold, M.D. 50 years of spermatogenesis: Sertoli cells and their interactions with germ cells. Biol. Reprod. 2018, 99, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Urbańska, K.; Orzechowski, A. Unappreciated Role of LDHA and LDHB to Control Apoptosis and Autophagy in Tumor Cells. Int. J. Mol. Sci. 2019, 20, 2085. [Google Scholar] [CrossRef] [Green Version]
- Read, J.A.; Winter, V.J.; Eszes, C.M.; Sessions, R.B.; Brady, R.L. Structural basis for altered activity of M- and H-isozyme forms of human lactate dehydrogenase. Proteins: Struct. Funct. Bioinform. 2001, 43, 175–185. [Google Scholar] [CrossRef]
- Liu, Z.; Yuan, M.; Meng, X.; Bie, H.; Yao, S. Identification of testicular Foxq1 as a critical modulator of lactate metabolism in mouse Sertoli cells. Histochem. Cell Biol. 2021, 156, 227–237. [Google Scholar] [CrossRef]
- Riera, M.; Meroni, S.; Gómez, G.; Schteingart, H.; Pellizzari, E.; Cigorraga, S. Regulation of Lactate Production by FSH, IL1β, and TNFα in Rat Sertoli Cells. Gen. Comp. Endocrinol. 2001, 122, 88–97. [Google Scholar] [CrossRef]
- Jia, G.-X.; Lin, Z.; Yan, R.-G.; Wang, G.-W.; Zhang, X.-N.; Li, C.; Tong, M.-H.; Yang, Q.-E. WTAP Function in Sertoli Cells Is Essential for Sustaining the Spermatogonial Stem Cell Niche. Stem Cell Rep. 2020, 15, 968–982. [Google Scholar] [CrossRef] [PubMed]
- Leblond, C.P. Spermatogenesis of rat, mouse, hamster and guinea pig as revealed by the periodic acid-fuchsin sulfurous acid technic. Am. J. Anat. 1952, 90, 167–215. [Google Scholar] [CrossRef]
- Yan, R.-G.; Li, B.-Y.; Yang, Q.-E. Function and transcriptomic dynamics of Sertoli cells during prospermatogonia development in mouse testis. Reprod. Biol. 2020, 20, 525–535. [Google Scholar] [CrossRef]
- Li, S.; Yang, Q.-E. Hypobaric hypoxia exposure alters transcriptome in mouse testis and impairs spermatogenesis in offspring. Gene 2022, 823, 146390. [Google Scholar] [CrossRef]
- Tao, H.-P.; Jia, G.-X.; Zhang, X.-N.; Wang, Y.-J.; Li, B.-Y.; Yang, Q.-E. Paternal hypoxia exposure impairs fertilization process and preimplantation embryo development. Zygote 2021, 30, 48–56. [Google Scholar] [CrossRef]
- Jaramillo, R.; Garrido, O.; Jorquera, B. Ultrastructural analysis of spermiogenesis and sperm morphology in Chorus giganteus (Lesson, 1829)(Prosobranchia: Muricidae). Veliger 1986, 29, 217–225. [Google Scholar]
- Jasmine, C.; Othman, S.; Carin, L.; Iurie, C.; Shuzhao, L.; Guillaume, B.; Wishart, D.S.; Jianguo, X. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, W1, W486–W494. [Google Scholar]
- Cermak, J.M. The Prenatal Availability of Choline Alters the Neurochemical Development of the Hippocampal Cholinergic System; Boston University: Boston, MA, USA, 1998. [Google Scholar]
- Yamaguchi, R.; Yamagata, K.; Ikawa, M.; Moss, S.B.; Okabe, M. Aberrant Distribution of ADAM3 in Sperm from Both Angiotensin-Converting Enzyme (Ace)- and Calmegin (Clgn)-Deficient Mice1. Biol. Reprod. 2006, 75, 760–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galardo, M.N.; Gorga, A.; Merlo, J.P.; Regueira, M.; Pellizzari, E.H.; Cigorraga, S.B.; Riera, M.F.; Meroni, S.B. Participation of HIFs in the regulation of Sertoli cell lactate production. Biochimie 2017, 132, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Maloy, S.; Hughes, K. (Eds.) Brenner’s Encyclopedia of Genetics; Academic Press: Cambridge, MA, USA, 2013; pp. 55–56. [Google Scholar]
- Ito, C.; Yamatoya, K.; Yoshida, K.; Fujimura, L.; Sugiyama, H.; Suganami, A.; Tamura, Y.; Hatano, M.; Miyado, K.; Toshimori, K. Deletion of Eqtn in mice reduces male fertility and sperm–egg adhesion. Reproduction 2018, 156, 579–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinkovska-Galcheva, V.; Srivastava, P.N. Phospholipids of rabbit and bull sperm membranes: Structural order parameter and steady-state fluorescence anistropy of membranes and membrane leaflets. Mol. Reprod. Dev. 1993, 35, 209–217. [Google Scholar] [CrossRef]
- Johnson, A.R.; Craciunescu, C.N.; Guo, Z.; Teng, Y.; Thresher, R.J.; Blusztajn, J.K.; Zeisel, S.H. Deletion of murine choline dehydrogenase results in diminished sperm motility. FASEB J. 2010, 24, 2752–2761. [Google Scholar] [CrossRef]
- Johnson, A.R.; Lao, S.; Wang, T.W.; Galanko, J.; Zeisel, S. Choline dehydrogenase polymorphism rs12676 is a functional variation associated with changes in human sperm cell function. PLoS ONE 2012, 7, e36047. [Google Scholar]
- Lewis, B.C.; Shim, H.; Li, Q.; Wu, C.S.; Lee, L.A.; Maity, A.; Dang, C.V. Identification of putative c-Myc-responsive genes: Characterization of rcl, a novel growth-related gene. Mol. Cell. Biol. 1997, 17, 4967–4978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Shi, M.; Xie, D.; Wei, D.; Jia, Z.; Zheng, S.; Gao, Y.; Huang, S.; Xie, K. FOXM1 Promotes the Warburg Effect and Pancreatic Cancer Progression via Transactivation of LDHA Expression. Clin. Cancer Res. 2014, 20, 2595–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Cui, J.; Du, J.; Wei, D.; Jia, Z.; Zhang, J.; Zhu, Z.; Gao, Y.; Xie, K. A Novel KLF4/LDHA Signaling Pathway Regulates Aerobic Glycolysis in and Progression of Pancreatic Cancer. Clin. Cancer Res. 2014, 20, 4370–4380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcivar, A.A.; Trasler, J.M.; Hake, L.E.; Salehi-Ashtiani, K.; Goldberg, E.; Hecht, N.B. DNA Methylation and Expression of the Genes Coding for Lactate Dehydrogenases A and C during Rodent Spermatogenesis1. Biol. Reprod. 1991, 44, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Rebourcet, D.; O’Shaughnessy, P.J.; Pitetti, J.-L.; Monteiro, A.; O’Hara, L.; Milne, L.; Tsai, Y.T.; Cruickshanks, L.; Riethmacher, D.; Guillou, F.; et al. Sertoli cells control peritubular myoid cell fate and support adult Leydig cell development in the prepubertal testis. Development 2014, 141, 2139–2149. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhang, Z.; Chang, C.; Yang, Z.; Wang, P.; Fu, H.; Wei, X.; Chen, E.; Tan, S.; Huang, W.; et al. A bioenergetic shift is required for spermatogonial differentiation. Cell Discov. 2020, 6, 56. [Google Scholar] [CrossRef]
- Voigt, A.L.; Kondro, D.A.; Powell, D.; Valli-Pulaski, H.; Ungrin, M.; Stukenborg, J.; Klein, C.; Lewis, I.A.; Orwig, K.E.; Dobrinski, I. Unique metabolic phenotype and its transition during maturation of juvenile male germ cells. FASEB J. 2021, 35, e21513. [Google Scholar] [CrossRef]
- Odet, F.; Gabel, S.; London, R.E.; Goldberg, E.; Eddy, E.M. Glycolysis and Mitochondrial Respiration in Mouse LDHC-Null Sperm1. Biol. Reprod. 2013, 88, 95. [Google Scholar] [CrossRef] [PubMed]
- Abdelatty, A.M.; Badr, O.A.M.; Mohamed, S.A.; Khattab, M.S.; Dessouki, S.H.M.; Farid, O.A.A.; Elolimy, A.A.; Sakr, O.G.; Elhady, M.A.; Mehaisen, G.M.K.; et al. Correction: Long term conjugated linoleic acid supplementation modestly improved growth performance but induced testicular tissue apoptosis and reduced sperm quality in male rabbit. PLoS ONE 2020, 15, e0231280. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.S.; Steeds, C.M.; Wiseman, R.W.; Pavlath, G.K. Phosphocreatine as an energy source for actin cytoskeletal rearrangements during myoblast fusion. J. Physiol. 2008, 586, 2841–2853. [Google Scholar] [CrossRef]
- Geer, B.W. Dietary Choline Requirements for Sperm Motility and Normal Mating Activity in Drosophila Melanogaster. Biol. Bull. 1967, 133, 548–566. [Google Scholar] [CrossRef]
- Johnson, A.R.; Zeisel, S. Choline dehydrogenase is necessary for normal energy metabolism in mouse sperm. FASEB J. 2011, 25, 354. [Google Scholar]
- Ibáñez, C.F.; Pelto-Huikko, M.; Söder, O.; Ritzèn, E.M.; Hersh, L.B.; Hökfelt, T.; Persson, H. Expression of choline acetyltransferase mRNA in spermatogenic cells results in an accumulation of the enzyme in the postacrosomal region of mature spermatozoa. Proc. Natl. Acad. Sci. USA 1991, 88, 3676–3680. [Google Scholar] [CrossRef] [Green Version]
- Parks, J.E.; Lynch, D.V. Lipid composition and thermotropic phase behavior of boar, bull, stallion, and rooster sperm membranes. Cryobiology 1992, 29, 255–266. [Google Scholar] [CrossRef]
- Zanetti, S.R.; Monclus, M.D.L.; Rensetti, D.E.; Fornés, M.W.; Aveldaño, M.I. Differential involvement of rat sperm choline glycerophospholipids and sphingomyelin in capacitation and the acrosomal reaction. Biochimie 2010, 92, 1886–1894. [Google Scholar] [CrossRef]
- Jiang, X.; Li, X.; Feng, W.; Qin, Y.; Li, Z.; Nie, H.; Qin, W.; Han, L.; Bai, W. Baking of methionine-choline deficient diet aggravates testis injury in mice. Food Chem. Toxicol. 2021, 154, 112245. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Liang, C.; Wang, N.; Zheng, H.; Wang, J. Choline supplementation alleviates fluoride-induced testicular toxicity by restoring the NGF and MEK expression in mice. Toxicol. Appl. Pharmacol. 2016, 310, 205–214. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.-N.; Tao, H.-P.; Li, S.; Wang, Y.-J.; Wu, S.-X.; Pan, B.; Yang, Q.-E. Ldha-Dependent Metabolic Programs in Sertoli Cells Regulate Spermiogenesis in Mouse Testis. Biology 2022, 11, 1791. https://doi.org/10.3390/biology11121791
Zhang X-N, Tao H-P, Li S, Wang Y-J, Wu S-X, Pan B, Yang Q-E. Ldha-Dependent Metabolic Programs in Sertoli Cells Regulate Spermiogenesis in Mouse Testis. Biology. 2022; 11(12):1791. https://doi.org/10.3390/biology11121791
Chicago/Turabian StyleZhang, Xiao-Na, Hai-Ping Tao, Shuang Li, Yu-Jun Wang, Shi-Xin Wu, Bo Pan, and Qi-En Yang. 2022. "Ldha-Dependent Metabolic Programs in Sertoli Cells Regulate Spermiogenesis in Mouse Testis" Biology 11, no. 12: 1791. https://doi.org/10.3390/biology11121791