
Alleviation of Cognitive and Physical Fatigue with Enzymatic Porcine Placenta Hydrolysate Intake through Reducing Oxidative Stress and Inflammation in Intensely Exercised Rats

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. EPPH Preparation
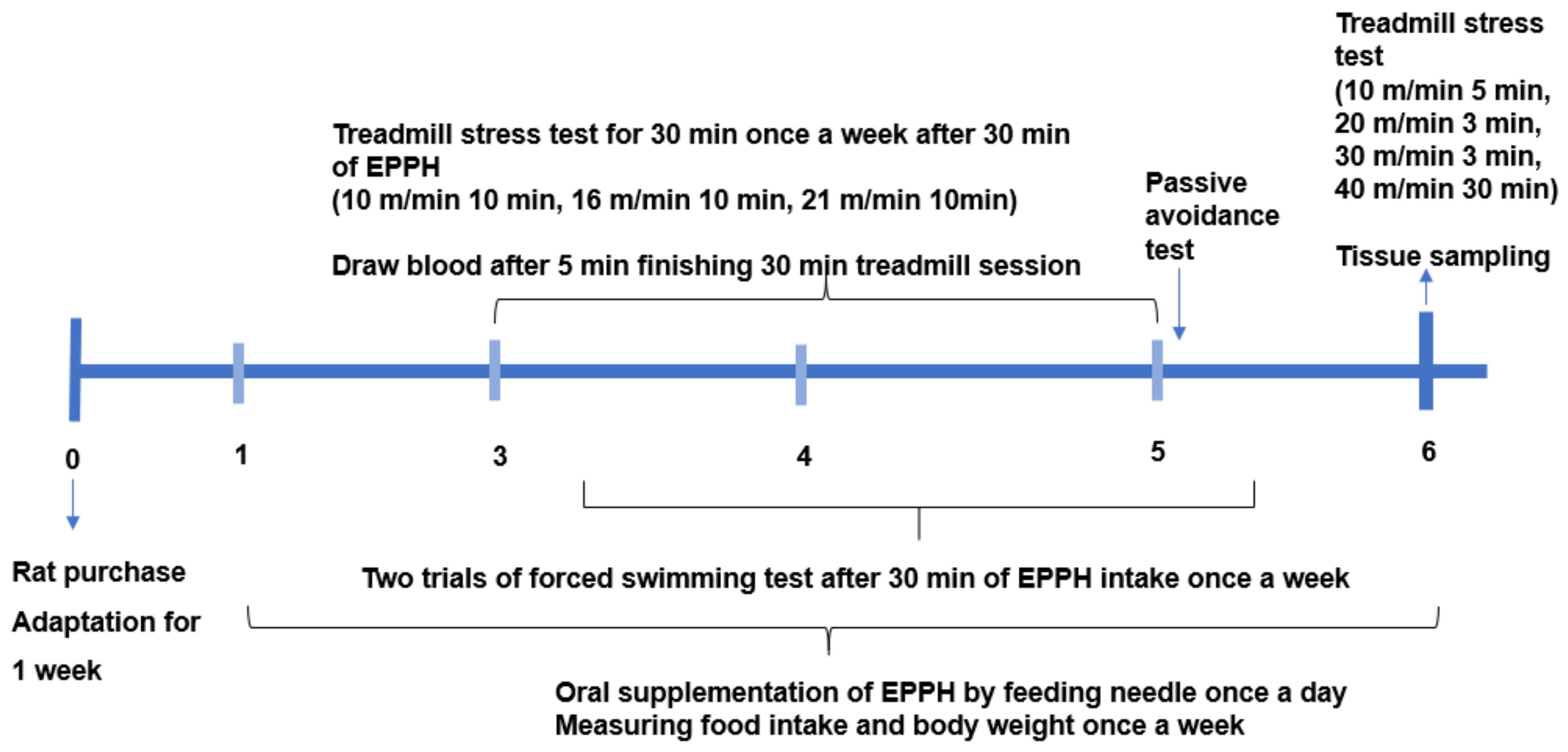
2.2. Animal Care and Experimental Design
2.3. Diet Preparation for Rats
2.4. Exercise-Induced Fatigue
2.5. Forced Swimming Test
2.6. Passive Avoidance Test
2.7. Tissue Collection and Assays
2.8. Quantitative Real-Time PCR
2.9. Statistical Analyses
3. Results
3.1. Amino Acid Composition of EPPH Product
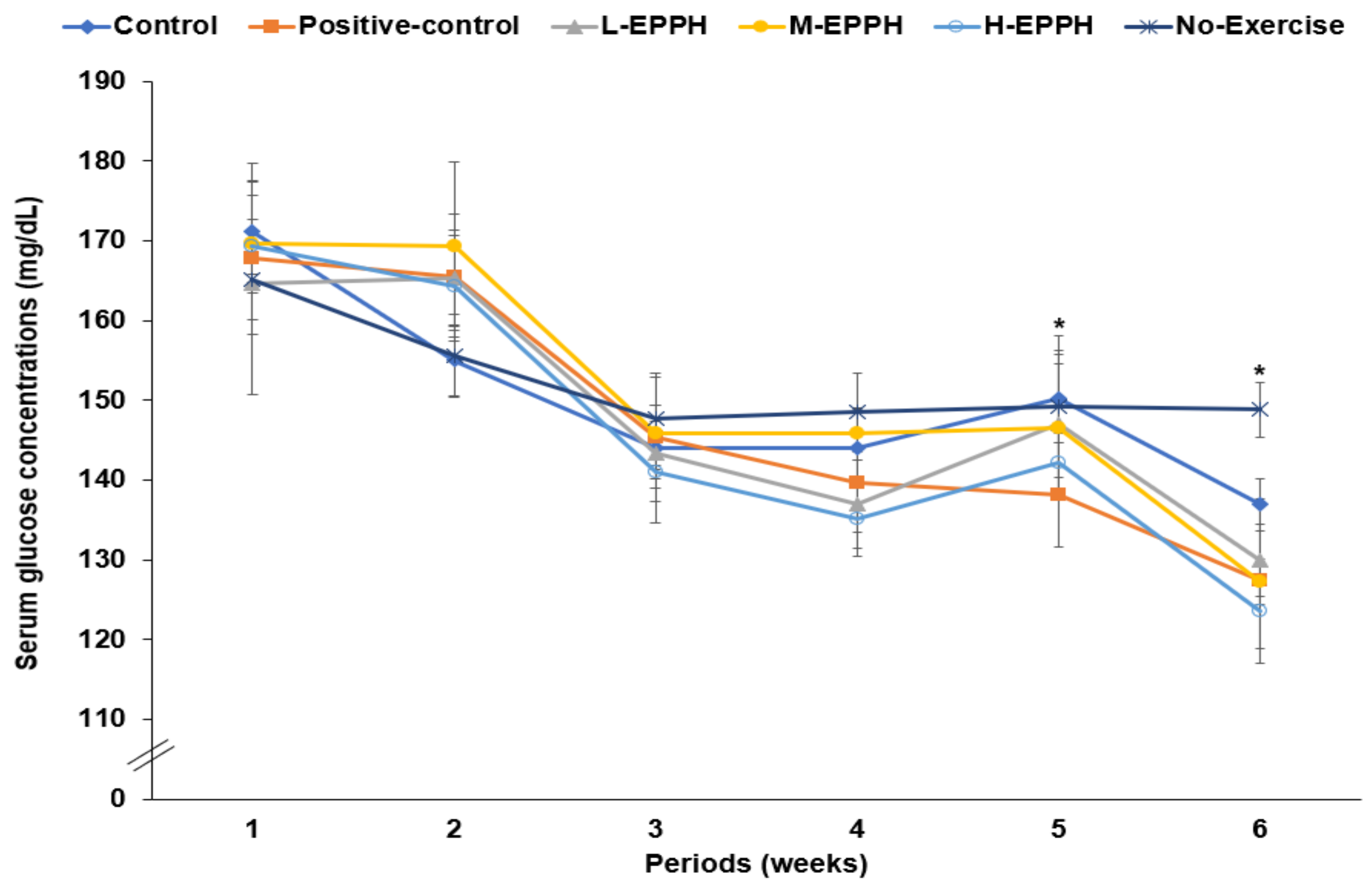
3.2. Body Weight, Organ Index, and Serum Glucose Concentrations
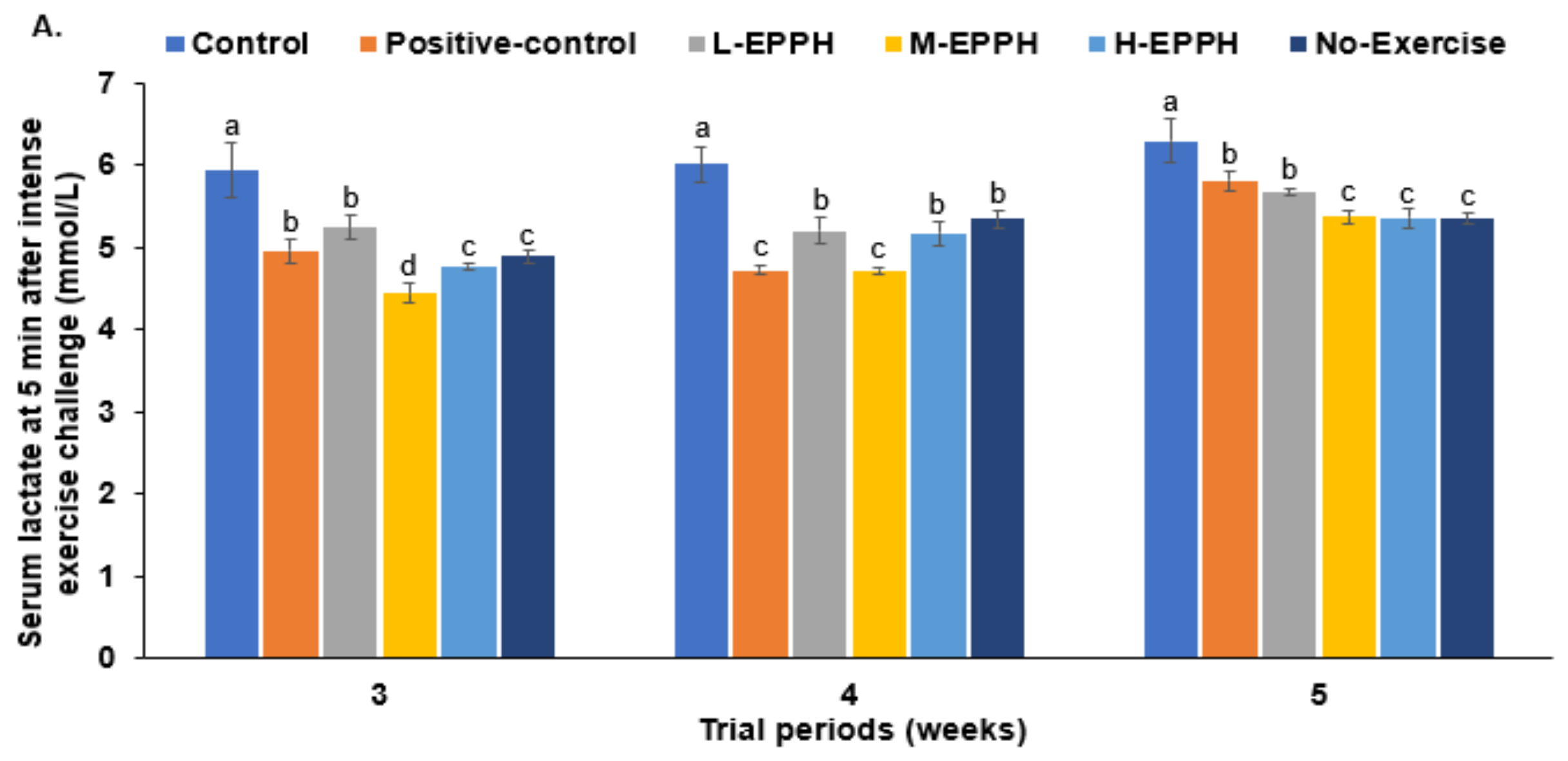
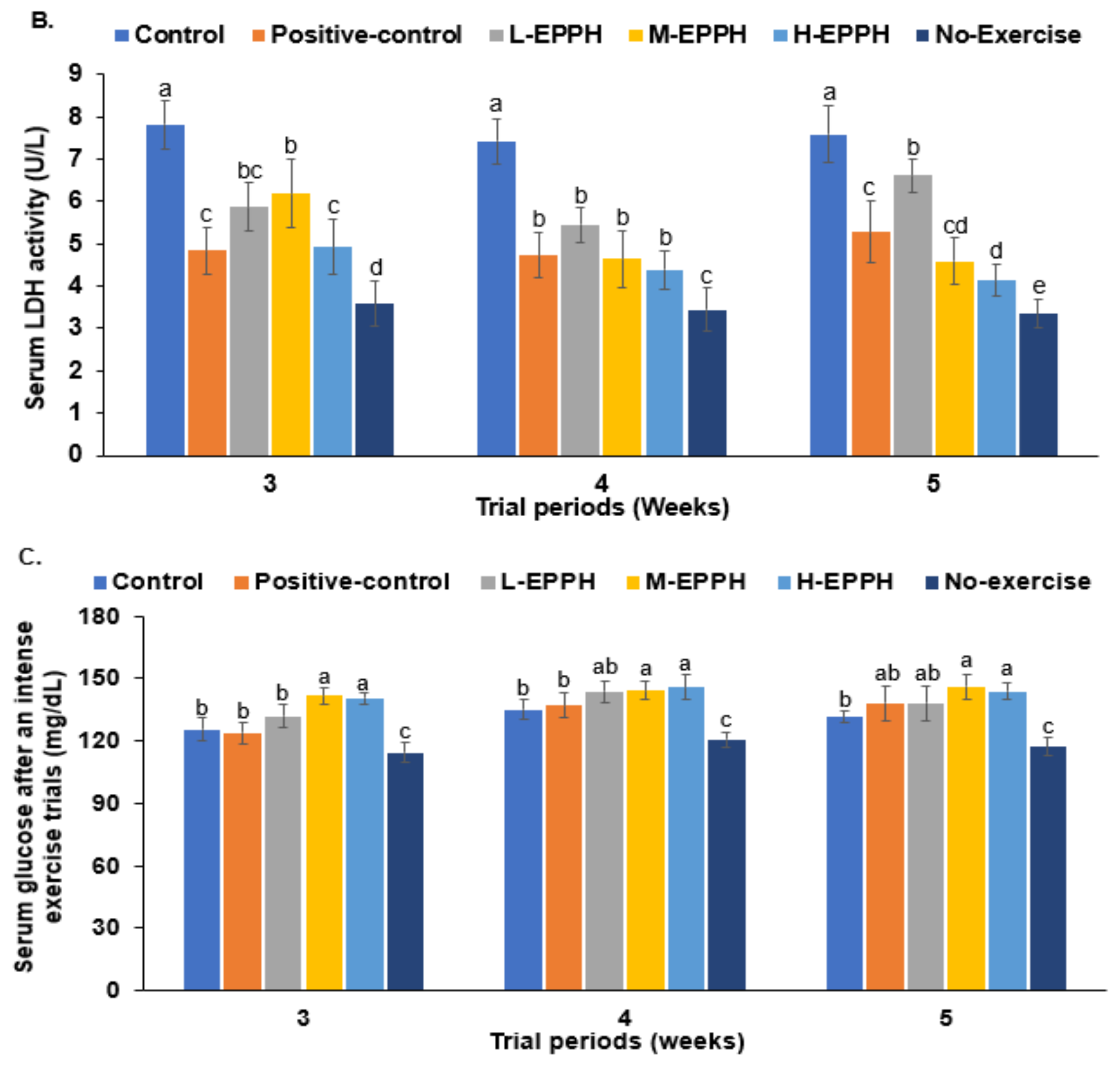
3.3. Exercise-Induced Fatigue in the Skeletal Muscles
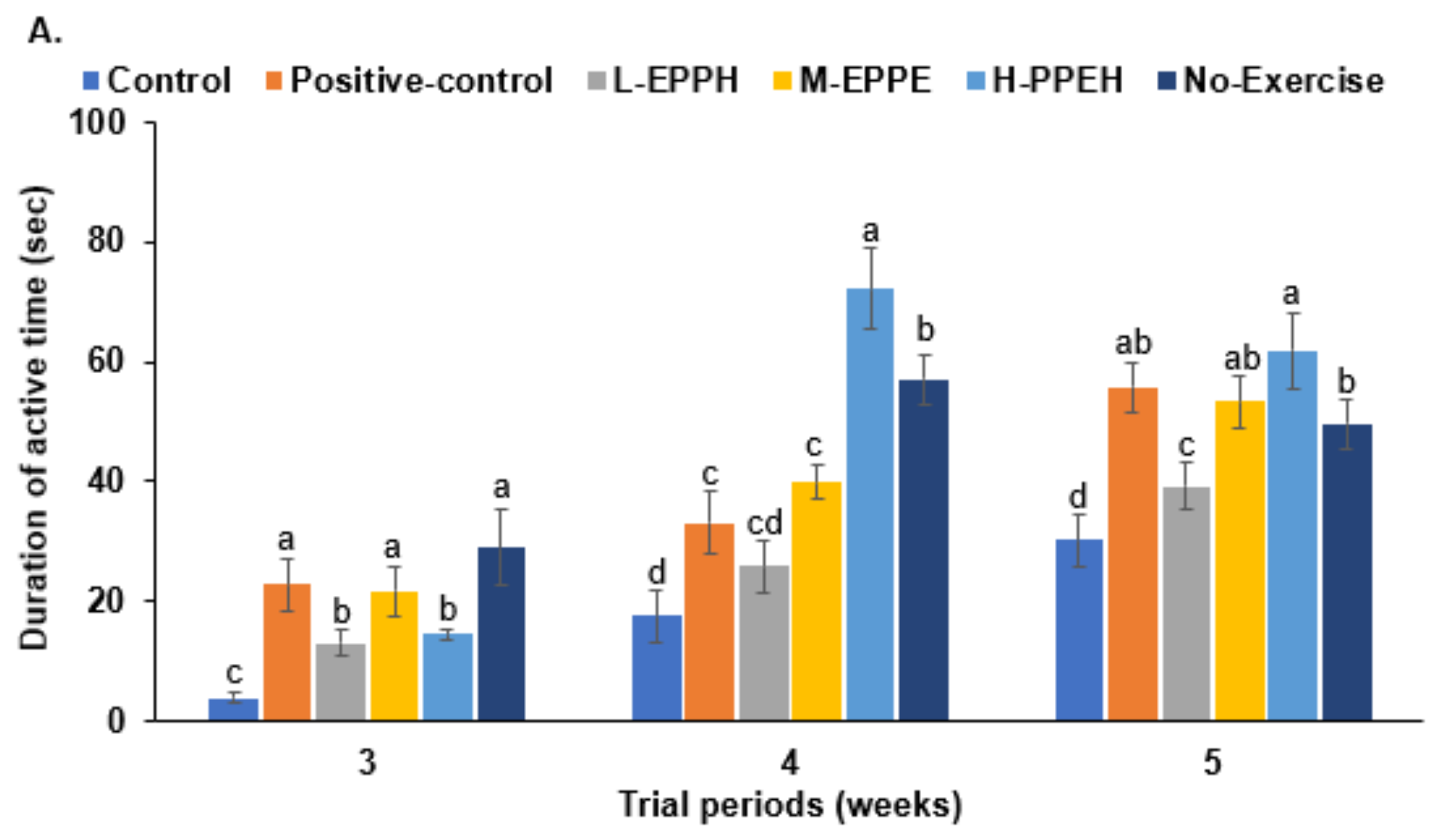
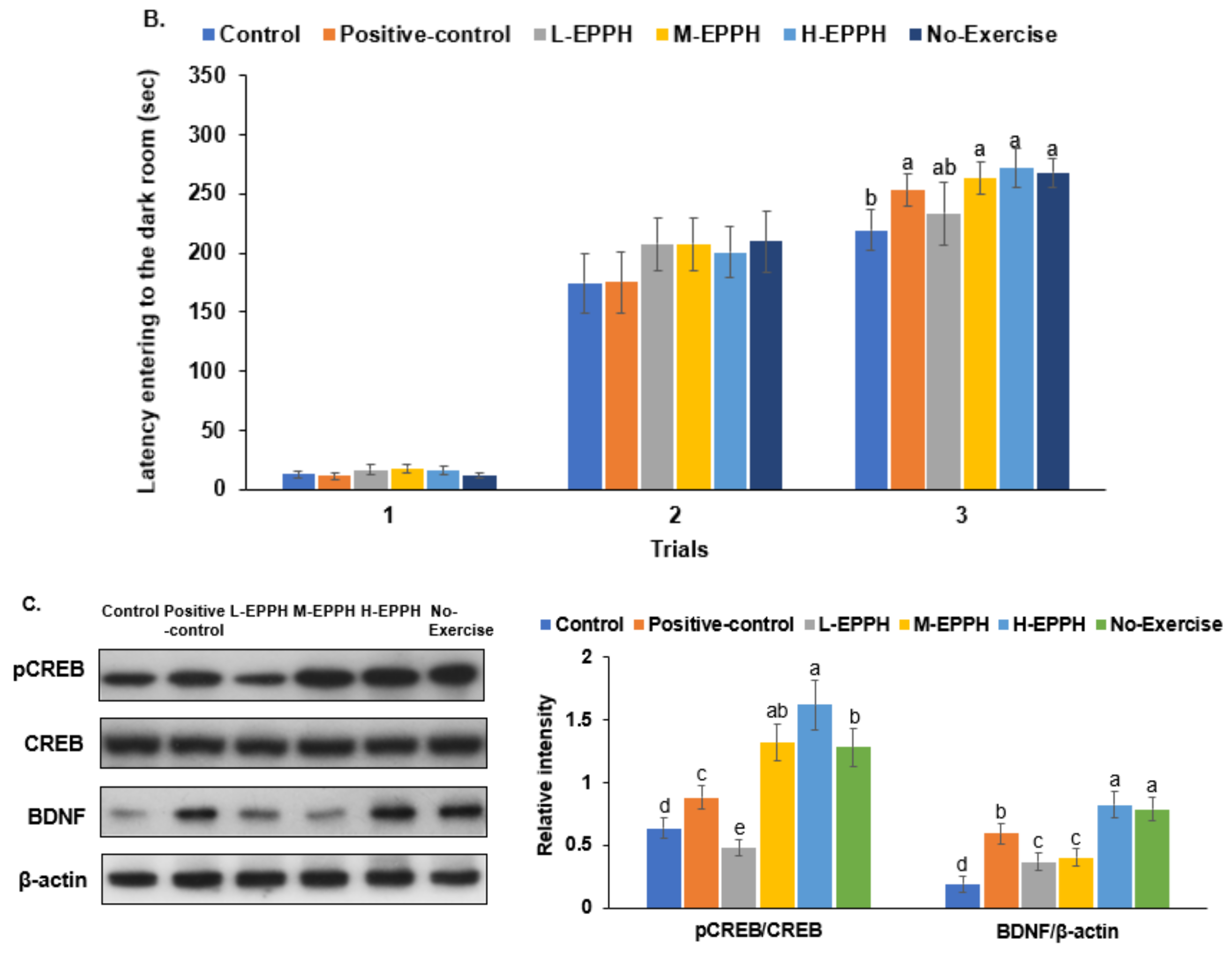
3.4. Forced Swimming and Passive Avoidance Tests
3.5. Exercise-Induced Cognitive Fatigue in the Brain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cordeiro, L.M.S.; Rabelo, P.C.R.; Moraes, M.M.; Teixeira-Coelho, F.; Coimbra, C.C.; Wanner, S.P.; Soares, D.D. Physical exercise-induced fatigue: The role of serotonergic and dopaminergic systems. Braz. J. Med. Biol. Res. 2017, 50, e6432. [Google Scholar] [CrossRef] [PubMed]
- Dotan, R.; Woods, S.; Contessa, P. On the reliability and validity of central fatigue determination. Eur. J. Appl. Physiol. 2021, 121, 2393–2411. [Google Scholar] [CrossRef] [PubMed]
- Gentile, D.; Beeler, D.; Wang, X.S.; Ben-Ayre, E.; Zick, S.M.; Bao, T.; Carlson, L.E.; Ghelman, R.; Master, V.; Tripathy, D.; et al. Cancer-Related Fatigue Outcome Measures in Integrative Oncology: Evidence for Practice and Research Recommendations. Oncology 2022, 36, 276–287. [Google Scholar] [PubMed]
- Sapra, A.; Bhandari, P. Chronic Fatigue Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Flockhart, M.; Nilsson, L.C.; Tais, S.; Ekblom, B.; Apró, W.; Larsen, F.J. Excessive exercise training causes mitochondrial functional impairment and decreases glucose tolerance in healthy volunteers. Cell Metab. 2021, 33, 957–970.e956. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.A.; Ross, L.M.; Slentz, C.A.; Huffman, K.M.; Kraus, W.E. Differential Effects of Amount, Intensity, and Mode of Exercise Training on Insulin Sensitivity and Glucose Homeostasis: A Narrative Review. Sports Med. Open 2022, 8, 90. [Google Scholar] [CrossRef]
- Park, S.W.; Son, S.M.; Lee, N.K. Exercise-induced muscle fatigue in the unaffected knee joint and its influence on postural control and lower limb kinematics in stroke patients. Neural Regen. Res. 2017, 12, 765–769. [Google Scholar]
- Hou, Y.; Tang, Y.; Wang, X.; Ai, X.; Wang, H.; Li, X.; Chen, X.; Zhang, Y.; Hu, Y.; Meng, X.; et al. Rhodiola Crenulata ameliorates exhaustive exercise-induced fatigue in mice by suppressing mitophagy in skeletal muscle. Exp. Ther. Med. 2020, 20, 3161–3173. [Google Scholar] [CrossRef]
- Meeusen, R.; Van Cutsem, J.; Roelands, B. Endurance exercise-induced and mental fatigue and the brain. Exp. Physiol. 2021, 106, 2294–2298. [Google Scholar] [CrossRef]
- Razgado-Hernandez, L.F.; Espadas-Alvarez, A.J.; Reyna-Velazquez, P.; Sierra-Sanchez, A.; Anaya-Martinez, V.; Jimenez-Estrada, I.; Bannon, M.J.; Martinez-Fong, D.; Aceves-Ruiz, J. The transfection of BDNF to dopamine neurons potentiates the effect of dopamine D3 receptor agonist recovering the striatal innervation, dendritic spines and motor behavior in an aged rat model of Parkinson’s disease. PLoS ONE 2015, 10, e0117391. [Google Scholar] [CrossRef] [Green Version]
- Proschinger, S.; Freese, J. Neuroimmunological and neuroenergetic aspects in exercise-induced fatigue. Exerc. Immunol. Rev. 2019, 25, 8–19. [Google Scholar]
- Moslemi, E.; Dehghan, P.; Khani, M.; Sarbakhsh, P.; Sarmadi, B. The effects of date seed (Phoenix dactylifera) supplementation on exercise-induced oxidative stress and aerobic and anaerobic performance following high-intensity interval training sessions: A randomized, double-blind, placebo-controlled trial. Br. J. Nutr. 2022, 1–12. [Google Scholar] [CrossRef]
- Miranda-Castro, S.; Aidar, F.J.; de Moura, S.S.; Marcucci-Barbosa, L.; Lobo, L.F.; de Assis Dias Martins-Júnior, F.; da Silva Filha, R.; Vaz de Castro, P.A.S.; Simões, E.S.A.C.; da Glória de Souza, D.; et al. The Curcumin Supplementation with Piperine Can Influence the Acute Elevation of Exercise-Induced Cytokines: Double-Blind Crossover Study. Biology 2022, 11, 573. [Google Scholar] [CrossRef]
- Cui, J.; Shi, C.; Xia, P.; Ning, K.; Xiang, H.; Xie, Q. Fermented Deer Blood Ameliorates Intense Exercise-Induced Fatigue via Modulating Small Intestine Microbiota and Metabolites in Mice. Nutrients 2021, 13, 1543. [Google Scholar] [CrossRef]
- Hureau, T.J.; Weavil, J.C.; Sidhu, S.K.; Thurston, T.S.; Reese, V.R.; Zhao, J.; Nelson, A.D.; Birgenheier, N.M.; Richardson, R.S.; Amann, M. Ascorbate attenuates cycling exercise-induced neuromuscular fatigue but fails to improve exertional dyspnea and exercise tolerance in COPD. J. Appl. Physiol. 2021, 130, 69–79. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Gaspar, D.; Flores-Félix, J.D.; Falcão, A.; Alves, G.; Silva, L.R. Effects of Functional Phenolics Dietary Supplementation on Athletes’ Performance and Recovery: A Review. Int. J. Mol. Sci. 2022, 23, 4652. [Google Scholar] [CrossRef]
- Leung, P.C. Placenta and umbilical cord in traditional chinese medicine. In Regenerative Medicine Using Pregnancy-Specific Biological Substances; Bhattacharya, N., Stubblefield, P., Eds.; Springer: London, UK, 2011; pp. 25–27. [Google Scholar]
- Ko, B.S.; Kim, D.S.; Kang, S.; Lee, N.R.; Ryuk, J.A.; Park, S. Wnt-signaling-mediated antiosteoporotic activity of porcine placenta hydrolysates in ovariectomized rats. Evid. Based Complement. Alternat. Med. 2012, 2012, 367698. [Google Scholar] [CrossRef] [Green Version]
- Nam, S.Y.; Kim, H.M.; Jeong, H.J. Anti-fatigue effect by active dipeptides of fermented porcine placenta through inhibiting the inflammatory and oxidative reactions. Biomed. Pharmacother. 2016, 84, 51–59. [Google Scholar] [CrossRef]
- Hou, L.; Li, K.E.; Hu, Y.; Bian, Y.; Ji, W.; Shi, K.; Li, Y.; Chen, M.; Li, J.; Liu, X.; et al. Evaluation of a rat model of exercise-induced fatigue using treadmill running with progressively increasing load. An. Acad. Bras. Cienc. 2019, 91, e20180957. [Google Scholar] [CrossRef]
- Aoi, W.; Ogaya, Y.; Takami, M.; Konishi, T.; Sauchi, Y.; Park, E.Y.; Wada, S.; Sato, K.; Higashi, A. Glutathione supplementation suppresses muscle fatigue induced by prolonged exercise via improved aerobic metabolism. J. Int. Soc. Sports Nutr. 2015, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wittert, G.; Taylor, A.W.; Adams, R.; Shi, Z. Associations between Macronutrient Intake and Obstructive Sleep Apnoea as Well as Self-Reported Sleep Symptoms: Results from a Cohort of Community Dwelling Australian Men. Nutrients 2016, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Moon, N.R.; Kang, S.; Kim, D.S. Ferulic acid and vinpocetine intake improves memory function by enhancing insulin sensitivity and reducing neuroinflammation and oxidative stress in type 2 diabetic animals with induced Alzheimer’s disease. J. Funct. Foods 2022, 95, 105180. [Google Scholar] [CrossRef]
- Yang, H.J.; Zhang, T.; Wu, X.G.; Kim, M.J.; Kim, Y.H.; Yang, E.S.; Yoon, Y.S.; Park, S. Aqueous Blackcurrant Extract Improves Insulin Sensitivity and Secretion and Modulates the Gut Microbiome in Non-Obese Type 2 Diabetic Rats. Antioxidants 2021, 10, 756. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Yang, W.; Lin, Y.; Du, C.; Huang, D.; Chen, S.; Yu, T.; Cong, X. Antioxidant and anti-fatigue activities of selenium-enriched peptides isolated from Cardamine violifolia protein hydrolysate. J. Funct. Foods 2021, 79, 104412. [Google Scholar] [CrossRef]
- Yoon, G.A.; Park, S. Antioxidant action of soy isoflavones on oxidative stress and antioxidant enzyme activities in exercised rats. Nutr. Res. Pract. 2014, 8, 618–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Kim, D.S.; Kang, S. Exercise training attenuates cerebral ischemic hyperglycemia by improving hepatic insulin signaling and β-cell survival. Life Sci. 2013, 93, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ma, X.; Cao, L.; Zhao, S.; Zhao, C.; Yin, S.; Hu, H. A Multi-Ingredient Formula Ameliorates Exercise-Induced Fatigue by Changing Metabolic Pathways and Increasing Antioxidant Capacity in Mice. Foods 2021, 10, 3120. [Google Scholar] [CrossRef]
- Lee, T.H.; Park, D.S.; Jang, J.Y.; Lee, I.; Kim, J.M.; Choi, G.S.; Oh, C.T.; Kim, J.Y.; Han, H.J.; Han, B.S.; et al. Human Placenta Hydrolysate Promotes Liver Regeneration via Activation of the Cytokine/Growth Factor-Mediated Pathway and Anti-oxidative Effect. Biol. Pharm. Bull. 2019, 42, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Han, N.R.; Kim, H.Y.; Kim, N.R.; Lee, W.K.; Jeong, H.; Kim, H.M.; Jeong, H.J. Leucine and glycine dipeptides of porcine placenta ameliorate physical fatigue through enhancing dopaminergic systems. Mol. Med. Rep. 2018, 17, 4120–4130. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, J.A.; VanDusseldorp, T.A.; Doyle, J.A.; Otis, J.S. Taurine in sports and exercise. J. Int. Soc. Sports Nutr. 2021, 18, 39. [Google Scholar] [CrossRef]
- Wang, D.; Wang, X. GLT-1 mediates exercise-induced fatigue through modulation of glutamate and lactate in rats. Neuropathology 2018, 38, 237–246. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Jing, Z.; Ordovas, J.M.; Wang, J.; Shen, L. Anti-fatigue and antioxidant effects of curcumin supplementation in exhaustive swimming mice via Nrf2/Keap1 signal pathway. Curr. Res. Food Sci. 2022, 5, 1148–1157. [Google Scholar] [CrossRef]
- Broskey, N.T.; Zou, K.; Dohm, G.L.; Houmard, J.A. Plasma Lactate as a Marker for Metabolic Health. Exerc. Sport Sci. Rev. 2020, 48, 119–124. [Google Scholar] [CrossRef]
- Williams, J.H.; Batts, T.W.; Lees, S. Reduced Muscle Glycogen Differentially Affects Exercise Performance and Muscle Fatigue. ISRN Physiol. 2013, 2013, 371235. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, H.G.; Lee, D.S.; Son, C.G. Oxidative Stress is a Convincing Contributor to Idiopathic Chronic Fatigue. Sci. Rep. 2018, 8, 12890. [Google Scholar] [CrossRef]
- Mach, J.; Midgley, A.W.; Dank, S.; Grant, R.S.; Bentley, D.J. The effect of antioxidant supplementation on fatigue during exercise: Potential role for NAD+(H). Nutrients 2010, 2, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Laosam, P.; Panpipat, W.; Yusakul, G.; Cheong, L.-Z.; Chaijan, M. Porcine placenta hydrolysate as an alternate functional food ingredient: In vitro antioxidant and antibacterial assessments. PLoS ONE 2021, 16, e0258445. [Google Scholar] [CrossRef]
- Komaroff, A.L. Inflammation correlates with symptoms in chronic fatigue syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, 8914–8916. [Google Scholar] [CrossRef] [Green Version]
- Hennigar, S.R.; McClung, J.P.; Pasiakos, S.M. Nutritional interventions and the IL-6 response to exercise. FASEB J. 2017, 31, 3719–3728. [Google Scholar] [CrossRef] [Green Version]
- Lacourt, T.E.; Vichaya, E.G.; Chiu, G.S.; Dantzer, R.; Heijnen, C.J. The High Costs of Low-Grade Inflammation: Persistent Fatigue as a Consequence of Reduced Cellular-Energy Availability and Non-adaptive Energy Expenditure. Front. Behav. Neurosci. 2018, 12, 78. [Google Scholar] [CrossRef] [Green Version]
- Karshikoff, B.; Sundelin, T.; Lasselin, J. Role of Inflammation in Human Fatigue: Relevance of Multidimensional Assessments and Potential Neuronal Mechanisms. Front. Immunol. 2017, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Haß, U.; Herpich, C.; Norman, K. Anti-Inflammatory Diets and Fatigue. Nutrients 2019, 11, 2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhang, J.; Li, H.; Liu, W.; Xi, Y.; Liu, X. A Comprehensive Comparison of Different Selenium Supplements: Mitigation of Heat Stress and Exercise Fatigue-Induced Liver Injury. Front. Nutr. 2022, 9, 917349. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.D.; Romine, M.W.; O’Connor, P.J.; Tomporowski, P.D. The influence of exercise-induced fatigue on cognitive function. J. Sports Sci. 2012, 30, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Northey, J.M.; Cherbuin, N.; Pumpa, K.L.; Smee, D.J.; Rattray, B. Exercise interventions for cognitive function in adults older than 50: A systematic review with meta-analysis. Br. J. Sports Med. 2018, 52, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Ochi, G.; Kuwamizu, R.; Suwabe, K.; Fukuie, T.; Hyodo, K.; Soya, H. Cognitive fatigue due to exercise under normobaric hypoxia is related to hypoxemia during exercise. Sci. Rep. 2022, 12, 9835. [Google Scholar] [CrossRef]
- Xue, X.; Liu, B.; Hu, J.; Bian, X.; Lou, S. The potential mechanisms of lactate in mediating exercise-enhanced cognitive function: A dual role as an energy supply substrate and a signaling molecule. Nutr. Metab. 2022, 19, 52. [Google Scholar] [CrossRef]
- Li, G.; Huang, S.; Xu, W.; Jiao, W.; Jiang, Y.; Gao, Z.; Zhang, J. The impact of mental fatigue on brain activity: A comparative study both in resting state and task state using EEG. BMC Neurosci. 2020, 21, 20. [Google Scholar] [CrossRef]
- Park, S.H.; Jang, S.; Lee, S.W.; Park, S.D.; Sung, Y.Y.; Kim, H.K. Akebia quinata Decaisne aqueous extract acts as a novel anti-fatigue agent in mice exposed to chronic restraint stress. J. Ethnopharmacol. 2018, 222, 270–279. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content (mg/mL) | Content (mg/mL) | Content (mg/mL) | |||
|---|---|---|---|---|---|
| ASP | 0.60 ± 0.25 | ARG | 1.43 ± 0.31 | VAL | 1.00 ± 0.40 |
| SER | 0.98 ± 0.20 | TRE | 0.59 ± 0.15 | MET | 0.62 ± 0.15 |
| GLU | 0.97 ± 0.19 | ALA | 1.45 ± 0.29 | LYS | 1.11 ± 0.23 |
| GLY | 1.30 ± 0.26 | PRO | 0.20 ± 0.10 | ILU | 0.84 ± 0.17 |
| HIS | 0.92 ± 0.18 | TYR | 0.36 ± 0.13 | LUE | 1.70 ± 0.34 |
| PHE | 0.61 ± 0.12 |
| Control (n = 10) | Positive-Control (n = 10) | L-EPPH (n = 10) | M-EPPH (n = 10) | H-EPPH (n = 10) | No-Exercise (n = 10) | |
|---|---|---|---|---|---|---|
| Body weight (g) | 429 ± 8.18 a | 425 ± 6.84 a | 429 ± 8.89 a | 416 ± 5.03 b | 418 ± 9.45 b | 420 ± 10.3 ab |
| Weight gain during the 6-week intervention (g) | 228 ± 11.5 ab | 220 ± 10.4 b | 233 ± 12.9 a | 208 ± 9.29 b | 213 ± 12.5 b | 219 ± 14.3 ab |
| Food intake (g/day) | 19.1 ± 0.40 | 18.4 ± 0.92 | 18.5 ± 0.68 | 19.5 ± 1.23 | 19.1 ± 1.56 | 18.9 ± 0.91 |
| Protein intake (g/day) | 5.17 ± 0.11 | 4.98 ± +0.25 | 5.01 ± 0.18 | 5.28 ± 0.33 | 5.17 ± 0.42 | 5.11 ± 0.25 |
| Food efficiency (%) | 12.2 ± 0.43 a | 12.3 ± 0.52 a | 12.7 ± 0.79 a | 11.2 ± 0.90 b | 11.8 ± 1.01 ab | 11.7 ± 0.83 ab |
| Epididymal fat pads (g) | 8.6 ± 0.48 b | 9.3 ± 0.86 a | 8.70 ± 0.86 ab | 7.05 ± 0.73 c | 7.94 ± 0.61 b | 8.01 ± 0.63 b |
| Retroperitoneal fat (g) | 10.1 ± 0.82 b | 11.5 ± 0.84 a | 9.58 ± 0.72 b | 8.12 ± 0.47 c | 8.71 ± 0.54 c | 8.48 ± 0.78 c |
| Visceral fat mass (% of body weight) | 4.36 ± 0.19 b | 4.89 ± 0.34 a | 4.26 ± 0.32 b | 3.65 ± 0.24 c | 3.97 ± 0.27 c | 3.93 ± 0.26 c |
| Muscle mass (% of body weight) | 2.43 ± 0.45 | 2.19 ± 0.29 | 2.35 ± 0.45 | 2.32 ± 0.33 | 2.39 ± 0.43 | 2.42 ± 0.44 |
| Liver index (% of body weight) | 3.91 ± 0.12 a | 3.68 ± 0.11 b | 3.75 ± 0.10 ab | 3.69 ± 0.07 b | 3.58 ± 0.10 b | 3.69 ± 0.09 b |
| Spleen index (% of body weight) | 4.33 ± 0.19 a | 4.41 ± 0.24 a | 4.24 ± 0.32 a | 3.63 ± 0.24 c | 3.91 ± 0.22 b | 3.91 ± 0.26 b |
| Control (n = 10) | Positive-Control (n = 10) | L-EPPH (n = 10) | M-EPPH (n = 10) | H-EPPH (n = 10) | No-Exercise (n = 10) | |
|---|---|---|---|---|---|---|
| At fasting state in the 3rd week before the exercise challenge. | ||||||
| Serum glucose (mg/dL) | 137 ± 9.09 a | 127 ± 8.75 b | 130 ± 9.70 ab | 127 ± 9.64 b | 123 ± 8.41 b | 148 ± 6.38 a |
| Serum insulin at fasting | 1.96 ± 0.24 a | 1.50 ± 0.31 b | 1.75 ± 0.31 ab | 1.47 ± 0.27 b | 1.50 ± 0.19 b | 1.90 ± 0.20 a |
| HOMA-IR | 11.9 ± 1.11 a | 8.49 ± 0.87 c | 10.1 ± 1.27 b | 8.32 ± 0.91 c | 8.23 ± 0.76 c | 12.6 ± 1.30 a |
| In the 5th week, after exercise-induced fatigue. | ||||||
| Serum lactate at fasting state (mmol/L) | 5.63 ± 0.25 a | 5.17 ± 0.12 b | 5.53 ± 0.16 a | 5.32 ± 0.10 b | 5.01 ± 0.09 c | 5.07 ± 0.14 c |
| Serum cortisol (ng/mL) | 129 ± 4.11 a | 120 ± 4.28 b | 126 ± 4.32 ab | 115 ± 4.53 bc | 113 ± 4.08 c | 104 ± 4.34 d |
| Serum total antioxidant capacity | 0.92 ± 0.05 c | 1.05 ± 0.05 a | 0.98 ± 0.05 b | 1.08 ± 0.04 a | 1.11 ± 0.06 a | 1.03 ± 0.06 b |
| Serum SOD (U/mL) | 1.35 ± 0.15 c | 1.72 ± 0.14 a | 1.44 ± 0.14 c | 1.63 ± 0.16 b | 1.76 ± 0.14 a | 1.48 ± 0.15 bc |
| Serum GSH-Px (U/mL) | 11.3 ± 0.85 d | 18.4 ± 1.78 a | 11.8 ± 0.91 d | 14.8 ± 1.07 c | 19.6 ± 1.19 a | 13.4 ± 1.05 c |
| Serum TNF-α (pg/mL) | 47.7 ± 1.13 a | 42.0 ± 1.28 c | 47.9 ± 1.44 a | 45.6 ± 1.06 b | 42.8 ± 0.83 c | 45.3 ± 1.35 b |
| Serum IL-1β (pg/mL) | 82.3 ± 1.05 a | 81.1 ± 1.23 a | 81.3 ± 1.17 a | 79.1 ± 0.60 b | 77.9 ± 0.70 c | 79.1 ± 0.94 b |
| Serum BUN (mg/dL) | 42.5 ± 1.67 a | 36.9 ± 1.60 c | 39.4 ± 1.37 b | 37.2 ± 0.89 bc | 36.5 ± 1.24 c | 36.2 ± 1.48 c |
| Serum creatinine (mg/dL) | 1.34 ± 0.06 a | 0.98 ± 0.10 c | 1.18 ± 0.13 b | 1.05 ± 0.15 bc | 0.98 ± 012 c | 1.02 ± 0.09 c |
| Serum creatine kinase (U/L) | 5.88 ± 0.57 a | 4.68 ± 0.24 b | 5.65 ± 0.41 a | 4.39 ± 0.41 b | 3.81 ± 0.36 c | 3.42 ± 0.40 c |
| Serum AST (U/L) | 72.5 ± 5.13 a | 62.9 ± 2.68 b | 61.5 ± 2.44 b | 52.0 ± 3.29 c | 46.3 ± 1.31 d | 52.8 ± 2.27 c |
| Serum ALT (U/L) | 63.3 ± 5.38 a | 53.4 ± 1.58 b | 51.2 ± 2.09 b | 39.0 ± 2.85 c | 33.3 ± 2.24 d | 35.8 ± 4.09 d |
| Control (n = 10) | Positive-Control (n = 10) | L-EPPH (n = 10) | M-EPPH (n = 10) | H-EPPH (n = 10) | No-Exercise (n = 10) | |
|---|---|---|---|---|---|---|
| In gastrocnemius muscle | ||||||
| Lactate (mg/mg protein) | 4.34 ± 0.19 a | 4.03 ± 0.12 b | 4.33 ± 0.18 a | 3.73 ± 0.20 c | 3.57 ± 0.31 c | 3.88 ± 0.19 bc |
| LDH activity (U/mg protein) | 69.4 ± 3.40 a | 62.0 ± 3.45 b | 65.2 ± 4.21 b | 64.3 ± 1.87 b | 57.2 ± 3.94 c | 64.2 ± 4.65 b |
| Creatine kinase (U/mg protein) | 36.6 ± 0.79 c | 34.2 ± 0.74 a | 33.8 ± 0.81 b | 32.8 ± 1.01 b | 32.4 ± 3.2 a | 33.9 ± 0.66 |
| Lipid peroxides (MDA nmol/mg protein) | 5.42 ± 0.48 a | 4.57 ± 0.37 b | 5.37 ± 0.51 a | 4.62 ± 0.42 b | 4.17 ± 0.36 c | 3.96 ± 0.34 c |
| SOD (U/mg protein) | 3.66 ± 0.05 d | 4.25 ± 0.21 b | 3.92 ± 0.07 c | 4.51 ± 0.16 b | 4.91 ± 0.11 a | 4.08 ± 0.16 c |
| GSH-Px (U/mg protein) | 43.7 ± 1.38 d | 65.6 ± 1.75 b | 48.4 ± 1.71 c | 63.6 ± 1.64 b | 69.5 ± 2.01 a | 47.5 ± 1.76 c |
| Glycogen (mg/g tissue) | 7.14 ± 0.52 d | 8.85 ± 0.89 b | 7.87 ± 1.05 c | 9.80 ± 0.97 a | 9.34 ± 1.06 b | 10.3 ± 1.34 a |
| In liver | ||||||
| Glycogen (mg/g tissue) | 0.76 ± 0.08 d | 0.95 ± 0.08 c | 0.89 ± 0.07 c | 1.08 ± 0.09 b | 1.15 ± 0.08 b | 1.34 ± 0.09 a |
| Lipid peroxides (MDA nmol/ mg protein) | 8.86 ± 0.78 a | 6.47 ± 0.71 b | 8.14 ± 0.77 a | 7.08 ± 0.65 b | 6.76 ± 0.69 bc | 6.17 ± 0.59 c |
| SOD (U/mg protein) | 2.45 ± 0.21 c | 3.34 ± 0.24 b | 2.58 ± 0.17 c | 3.42 ± 0.23 b | 4.05 ± 0.28 a | 3.27 ± 0.27 b |
| GSH-Px (U/mg protein) | 31.5 ± 1.08 d | 45.8 ± 1.34 b | 33.9 ± 1.31 c | 44.9 ± 1.54 b | 49.8 ± 1.38 a | 35.1 ± 1.34 c |
| Control | Positive-C | L-EPPH | M-EPPH | H-EPPH | No-Exercise | |
|---|---|---|---|---|---|---|
| Hypothalamic corticosterone expression (AU) | 1 a | 0.88 ± 0.04 b | 1.05 ± 0.08 a | 0.66 ± 0.04 c | 1.71 ± 0.04 c | 0.71 ± 0.03 c |
| Hypothalamic lipid peroxides (MDA nmol/mg protein) | 54.5 ± 4.6 a | 47.2 ± 4.2 b | 47.4 ± 7.2 b | 33.2 ± 3.9 d | 39.9 ± 4.1 c | 34.8 ± 3.7 d |
| Hippocampal TNF-α (pg/mg protein) | 10.7 ± 1.03 a | 8.55 ± 0.97 b | 8.65 ± 0.97 b | 7.75 ± 0.65 c | 7.41 ± 0.51 c | 7.85 ± 0.73 c |
| Hippocampal IL-1β (pg/mg protein) | 6.84 ± 0.47 a | 5.81 ± 0.46 b | 5.98 ± 0.61 ab | 5.04 ± 0.52 c | 4.97 ± 0.41 c | 6.12 ± 0.34 b |
| Hippocampal BDNF expression (AU) | 1 a | 1.18 ± 0.11 b | 0.92 ± 0.11 ab | 1.45 ± 0.10 bc | 1.37 ± 0.09 bc | 1.47 ± 0.14 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.J.; Zhang, T.; Kim, K.N.; Bae, G.W.; Yoon, S.M.; Yue, Y.; Wu, X.; Park, S. Alleviation of Cognitive and Physical Fatigue with Enzymatic Porcine Placenta Hydrolysate Intake through Reducing Oxidative Stress and Inflammation in Intensely Exercised Rats. Biology 2022, 11, 1739. https://doi.org/10.3390/biology11121739
Kim MJ, Zhang T, Kim KN, Bae GW, Yoon SM, Yue Y, Wu X, Park S. Alleviation of Cognitive and Physical Fatigue with Enzymatic Porcine Placenta Hydrolysate Intake through Reducing Oxidative Stress and Inflammation in Intensely Exercised Rats. Biology. 2022; 11(12):1739. https://doi.org/10.3390/biology11121739
Chicago/Turabian StyleKim, Min Ju, Ting Zhang, Keun Nam Kim, Gun Won Bae, Sun Myung Yoon, Yu Yue, Xuangao Wu, and Sunmin Park. 2022. "Alleviation of Cognitive and Physical Fatigue with Enzymatic Porcine Placenta Hydrolysate Intake through Reducing Oxidative Stress and Inflammation in Intensely Exercised Rats" Biology 11, no. 12: 1739. https://doi.org/10.3390/biology11121739