
A Novel ssDNA Aptamer Targeting Carcinoembryonic Antigen: Selection and Characterization

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Proteins
2.2. DNA Library, Primers
2.3. Cell Lines
2.4. In Vitro Selection of Aptamers
2.5. Next Generation Sequencing and Bioinformatics Analysis
2.6. Affinity Analysis
2.7. Confocal Microscopy
2.8. Statistical Analysis
3. Results
3.1. Selection of DNA Aptamers
3.2. Affinity Analysis of SELEX Cycle Aptamer Pools
3.3. NGS and Bioinformatics Analysis
3.4. Characterization of CEA Aptamers Using ELONA
3.5. Confocal Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sudhakar, A. History of Cancer, Ancient and Modern Treatment Methods. J. Cancer Sci. Ther. 2009, 1, i–iv. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 1 September 2022).
- World Health Organization. Cancer. Available online: https://www.who.int/health-topics/cancer#tab=tab_1 (accessed on 1 September 2022).
- National Cancer Institute. How Cancer Is Diagnosed. Available online: https://www.cancer.gov/about-cancer/diagnosis-staging/diagnosis (accessed on 17 September 2018).
- Greenwald, Z.R.; El-Zein, M.; Bouten, S.; Ensha, H.; Vazquez, F.L.; Franco, E.L. Mobile Screening Units for the Early Detection of Cancer: A Systematic Review. Cancer Epidemiol. Biomark. Prev. 2017, 26, 1679–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S. Benefits and Limitations of Regular Cancer Screening. Available online: http://www.cancer.ca/en/prevention-and-screening/reduce-cancer-risk/find-cancer-early/screening-for-cancer/benefits-and-limitations-of-regular-cancer-screening/?region=on (accessed on 4 September 2018).
- Srivastava, A.; Creek, D. Discovery and Validation of Clinical Biomarkers of Cancer: A Review Combining Metabolomics and Proteomics. Proteomics 2018, 19, 1700448. [Google Scholar] [CrossRef] [PubMed]
- Bacac, M.; Fauti, T.; Sam, J.; Colombetti, S.; Weinzierl, T.; Ouaret, D.; Bodmer, W.; Lehmann, S.; Hofer, T.; Hosse, R.J.; et al. A Novel Carcinoembryonic Antigen T-Cell Bispecific Antibody (CEA TCB) for the Treatment of Solid Tumors. Clin. Cancer Res. 2016, 22, 3286–3297. [Google Scholar] [CrossRef] [Green Version]
- Grunnet, M.; Sorensen, J.B. Carcinoembryonic Antigen (CEA) as Tumor Marker in Lung Cancer. Lung. Cancer 2012, 76, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Mo, Y.; Wu, C. Prognostic values of CEA, CA19-9, and CA72-4 in patients with stages I-III colorectal cancer. Int. J. Clin. Exp. Pathol. 2020, 13, 1608–1614. [Google Scholar] [PubMed]
- Ordoñez, C.; Screaton, R.A.; Ilantzis, C.; Stanners, C.P. Human Carcinoembryonic Antigen Functions as a General Inhibitor of Anoikis. Cancer Res. 2000, 60, 3419–3424. [Google Scholar] [PubMed]
- Meng, Q.; Shi, S.; Liang, C.; Liang, D.; Xu, W.; Ji, S.; Zhang, B.; Ni, Q.; Xu, J.; Yu, X. Diagnostic And Prognostic Value Of Carcinoembryonic Antigen In Pancreatic Cancer: A Systematic Review And Meta-Analysis. OncoTargets Ther. 2017, 10, 4591–4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadoniou, N.; Kosmas, C.; Gennatas, K.; Polyzos, A.; Mouratidou, D.; Skopeliti, E.; Tzivras, M.; Sougioultzis, S.; Papastratis, G.; Karatzas, G.; et al. Prognostic factors in patients with locally advanced (unresectable) or metastatic pancreatic adenocarcinoma: A retrospective analysis. Anticancer. Res. 2008, 28, 543–549. [Google Scholar] [PubMed]
- Reitz, D.; Gerger, A.; Seidel, J.; Kornprat, P.; Samonigg, H.; Stotz, M.; Szkandera, J.; Pichler, M. Combination Of Tumour Markers CEA And CA19-9 Improves The Prognostic Prediction In Patients With Pancreatic Cancer. J. Clin. Pathol. 2015, 68, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, J.; Lin, J. 3,3′–Diaminobenzidine (DAB)–H2O2–HRP Voltammetric Enzyme-Linked Immunoassay for the Detection of Carcionembryonic Antigen. Bioelectrochemistry 2008, 72, 47–52. [Google Scholar] [CrossRef]
- Yan, F.; Zhou, J.; Lin, J.; Ju, H.; Hu, X. Flow Injection Immunoassay for Carcinoembryonic Antigen Combined with Time-Resolved Fluorometric Detection. J. Immunol. Methods 2005, 305, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, B.; Yin, H.; Wei, J.; Qian, X.; Yu, L. Selection of DNA Aptamer That Specific Binding Human Carcinoembryonic Antigen in Vitro. J. Nanjing Med. Univ. 2007, 21, 277–281. [Google Scholar] [CrossRef]
- Liu, M.; Yu, X.; Chen, Z.; Yang, T.; Yang, D.; Liu, Q.; Du, K.; Li, B.; Wang, Z.; Li, S.; et al. Aptamer Selection and Applications for Breast Cancer Diagnostics and Therapy. J. Nanobiotechnology 2017, 15, 81. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In Vitro Selection of RNA Molecules That Bind Specific Ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Tuerk, C.; Gold, L. Systematic Evolution of Ligands by Exponential Enrichment: RNA Ligands to Bacteriophage T4 DNA Polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Yang, S. Replacing Antibodies with Aptamers in Lateral Flow Immunoassay. Biosens. Bioelectron. 2015, 71, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.; Strom, M.; Hammond, D.; Shigdar, S. Anything You Can Do, I Can Do Better: Can Aptamers Replace Antibodies in Clinical Diagnostic Applications? Molecules 2019, 24, 4377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Tan, W.; Zu, Y. Aptamers: Versatile Molecular Recognition Probes for Cancer Detection. The Analyst 2016, 141, 403–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayasena, S.D. Aptamers: An Emerging Class of Molecules That Rival Antibodies in Diagnostics. Clin. Chem. 1999, 45, 1628–1650. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T. Aptamers and SELEX: The Technology. World Pat. Inf. 2003, 25, 123–129. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, Y.; Di, Y.; He, L.; Liao, S.; Li, D.; Liu, X. In Vitroselection of DNA Aptamers for the Development of Chemiluminescence Aptasensor for Neuron-Specific Enolase (NSE) Detection. RSC Adv. 2019, 9, 15513–15520. [Google Scholar] [CrossRef] [Green Version]
- Witt, M.; Walter, J.-G.; Stahl, F. Aptamer Microarrays—Current Status and Future Prospects. Microarrays 2015, 4, 115–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKeague, M.; Calzada, V.; Cerchia, L.; DeRosa, M.; M Heemstra, J.; Janjic, N.; E Johnson, P.; Kraus, L.; Limson, J.; Mayer, G.; et al. The Minimum Aptamer Publication Standards (MAPS Guidelines) For De Novo Aptamer Selection. Aptamers 2022, 6, 10–18. [Google Scholar]
- Van der Straaten, T. Next-Generation Sequencing: Current Technologies and Applications. ChemMedChem 2015, 10, 419–420. [Google Scholar] [CrossRef]
- Metzker, M.L. Sequencing Technologies—The next Generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izabella Abreu de Melo, M.; Rodrigues Correa, C.; da Silva Cunha, P.; Miranda de Góes, A.; Assis Gomes, D.; Silva Ribeiro de Andrade, A. DNA Aptamers Selection for Carcinoembryonic Antigen (CEA). Bioorg. Med. Chem. Lett. 2020, 30, 127278. [Google Scholar] [CrossRef] [PubMed]
- Torrini, F.; Palladino, P.; Brittoli, A.; Baldoneschi, V.; Minunni, M.; Scarano, S. Characterization of Troponin T Binding Aptamers for an Innovative Enzyme-Linked Oligonucleotide Assay (ELONA). Anal. Bioanal. Chem. 2019, 411, 7709–7716. [Google Scholar] [CrossRef] [PubMed]
- Sypabekova, M.; Bekmurzayeva, A.; Wang, R.; Li, Y.; Nogues, C.; Kanayeva, D. Selection, Characterization, and Application of DNA Aptamers for Detection of Mycobacterium Tuberculosis Secreted Protein MPT64. Tuberculosis 2017, 104, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Drolet, D.W.; Moon-McDermott, L.; Romig, T.S. An Enzyme-Linked Oligonucleotide Assay. Nat. Biotechnol. 1996, 14, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.G.; Stecker, J.R.; Carrillo, M.P.; Phillips, T.; Savage, A.; Garcia, D.M.; Koke, J.R. Chapter 4: Novel aptamer-based therapeutic strategies. In Biomedical Applications of Aptamers; Bruno, J.G., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2013; pp. 55–72. [Google Scholar]
- Laing, C.; Schlick, T. Computational Approaches to RNA Structure Prediction, Analysis, and Design. Curr. Opin. Struct. Biol. 2011, 21, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Zulkeflee Sabri, M.; Azzar Abdul Hamid, A.; Mariam Sayed Hitam, S.; Zulkhairi Abdul Rahim, M. In-Silico Selection of Aptamer: A Review on the Revolutionary Approach to Understand the Aptamer Design and Interaction Through Computational Chemistry. Mater. Today Proc. 2019, 19, 1572–1581. [Google Scholar] [CrossRef]
- Sabri, M.Z.; Abdul Hamid, A.A.; Sayed Hitam, S.M.; Abdul Rahim, M.Z. In Silico Screening of Aptamers Configuration against Hepatitis B Surface Antigen. Adv. Bioinforma. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, S.; Yan, J.; Xiong, H.; Liu, Y.; Peng, D.; Liu, Z. Investigations on the Interface of Nucleic Acid Aptamers and Binding Targets. The Analyst 2018, 143, 5317–5338. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Lin, Y.; Wang, C.; Li, F.; Wang, Z.; Zhang, H.; Li, X.-F.; Le, X.C. Aptamer Binding Assays for Proteins: The Thrombin Example—A Review. Anal. Chim. Acta 2014, 837, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Riccitelli, N.J.; Lupták, A. Computational Discovery of Folded RNA Domains in Genomes and in Vitro Selected Libraries. Methods 2010, 52, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.; Pina, A.; Sousa, D.; Ferreira, D.; Santos-Pereira, C.; Rodrigues, J.; Melo, L.; Sales, G.; Sousa, S.; Rodrigues, L. Identification Of Novel Aptamers Targeting Cathepsin B-Overexpressing Prostate Cancer Cells. Mol. Syst. Des. Eng. 2022, 7, 637–650. [Google Scholar] [CrossRef]
- Tan, S.Y.; Acquah, C.; Sidhu, A.; Ongkudon, C.M.; Yon, L.S.; Danquah, M.K. SELEX Modifications and Bioanalytical Techniques for Aptamer–Target Binding Characterization. Crit. Rev. Anal. Chem. 2016, 46, 521–537. [Google Scholar] [CrossRef] [PubMed]
- de Castro, M.A.G.; Rammner, B.; Opazo, F. Aptamer Stainings for Super-Resolution Microscopy. In Nucleic Acid Aptamers; Mayer, G., Ed.; Methods in Molecular Biology; Springer New York: New York, NY, USA, 2016; Volume 1380, pp. 197–210. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An Interactive Tool for Rooted Phylogenetic Trees and Networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Antczak, M.; Popenda, M.; Zok, T.; Sarzynska, J.; Ratajczak, T.; Tomczyk, K.; Adamiak, R.W.; Szachniuk, M. New Functionality of RNAComposer: Application to Shape the Axis of MiR160 Precursor Structure. Acta Biochim. Pol. 2017, 63, 737–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korotkova, N.; Yang, Y.; Le Trong, I.; Cota, E.; Demeler, B.; Marchant, J.; Thomas, W.E.; Stenkamp, R.E.; Moseley, S.L.; Matthews, S. Binding of Dr Adhesins of Escherichia Coli to Carcinoembryonic Antigen Triggers Receptor Dissociation: Dr Adhesins Trigger CEA Dimer Dissociation. Mol. Microbiol. 2007, 67, 420–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotherham, L.S.; Maserumule, C.; Dheda, K.; Theron, J.; Khati, M. Selection and Application of SsDNA Aptamers to Detect Active TB from Sputum Samples. PLoS ONE 2012, 7, e46862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laguinge, L.; Bajenova, O.; Bowden, E.; Sayyah, J.; Thomas, P.; Juhl, H. Surface Expression and CEA Binding of HnRNP M4 Protein in HT29 Colon Cancer Cells. Anticancer. Res. 2005, 25, 23–31. [Google Scholar]
- Yarizadeh, K.; Behbahani, M.; Mohabatkar, H.; Noorbakhsh, A. Computational Analysis and Optimization of Carcinoembryonic Antigen Aptamers and Experimental Evaluation. J. Biotechnol. 2019, 306, 1–8. [Google Scholar] [CrossRef]
- Pierce, B.G.; Wiehe, K.; Hwang, H.; Kim, B.-H.; Vreven, T.; Weng, Z. ZDOCK Server: Interactive Docking Prediction of Protein-Protein Complexes and Symmetric Multimers. Bioinformatics 2014, 30, 1771–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, M.; Fernández-Algar, M.; Fernández-Chamorro, J.; Ramajo, J.; Martínez-Salas, E.; Briones, C. A Combined ELONA-(RT)QPCR Approach for Characterizing DNA and RNA Aptamers Selected against PCBP-2. Molecules 2019, 24, 1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivekananda, J.; Kiel, J.L. Anti-Francisella Tularensis DNA Aptamers Detect Tularemia Antigen from Different Subspecies by Aptamer-Linked Immobilized Sorbent Assay. Lab. Investig. 2006, 86, 610–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Jee, M.H.; Kwon, O.S.; Keum, S.J.; Jang, S.K. Infectivity of Hepatitis C Virus Correlates with the Amount of Envelope Protein E2: Development of a New Aptamer-Based Assay System Suitable for Measuring the Infectious Titer of HCV. Virology 2013, 439, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, R.E. RNA Methodologies: Laboratory Guide for Isolation and Characterization, 4th ed.; Elsevier/Academic Press: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Pauff, S.; Withers, J.M.; McKean, I.J.; Mackay, S.P.; Burley, G.A. Synthetic Biological Approaches for RNA Labelling and Imaging: Design Principles and Future Opportunities. Curr. Opin. Biotechnol. 2017, 48, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Ezra, A.; Rabinovich-Nikitin, I.; Rabinovich-Toidman, P.; Solomon, B. Multifunctional Effects of Human Serum Albumin Toward Neuroprotection in Alzheimer Disease. In Neuroprotection in Alzheimer’s Disease; Elsevier: Amsterdam, The Netherlands, 2017; pp. 217–238. [Google Scholar] [CrossRef]
- Peters, T. All about Albumin: Biochemistry, Genetics, and Medical Applications; Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Quinlan, G.J.; Martin, G.S.; Evans, T.W. Albumin: Biochemical Properties and Therapeutic Potential. Hepatology 2005, 41, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.; Adams, M.C.; Naik, R.R.; Milam, V.T. Analyzing Secondary Structure Patterns in DNA Aptamers Identified via CompELS. Molecules 2019, 24, 1572. [Google Scholar] [CrossRef] [Green Version]
- Jeddi, I.; Saiz, L. Three-Dimensional Modeling of Single Stranded DNA Hairpins for Aptamer-Based Biosensors. Sci. Rep. 2017, 7, 1178. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Hu, L.; Zhang, B.-T.; Lu, A.; Wang, Y.; Yu, Y.; Zhang, G. Artificial Intelligence in Aptamer–Target Binding Prediction. Int. J. Mol. Sci. 2021, 22, 3605. [Google Scholar] [CrossRef] [PubMed]
- Schrewe, H.; Thompson, J.; Bona, M.; Hefta, L.J.; Maruya, A.; Hassauer, M.; Shively, J.E.; von Kleist, S.; Zimmermann, W. Cloning of the Complete Gene for Carcinoembryonic Antigen: Analysis of Its Promoter Indicates a Region Conveying Cell Type-Specific Expression. Mol. Cell Biol. 1990, 10, 2738–2748. [Google Scholar] [CrossRef] [PubMed]
- Orava, E.W.; Abdul-Wahid, A.; Huang, E.H.-B.; Mallick, A.I.; Gariépy, J. Blocking the Attachment of Cancer Cells in Vivo with DNA Aptamers Displaying Anti-Adhesive Properties against the Carcinoembryonic Antigen. Mol. Oncol. 2013, 7, 799–811. [Google Scholar] [CrossRef]
- Hasegawa, H.; Savory, N.; Abe, K.; Ikebukuro, K. Methods for Improving Aptamer Binding Affinity. Molecules 2016, 21, 421. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
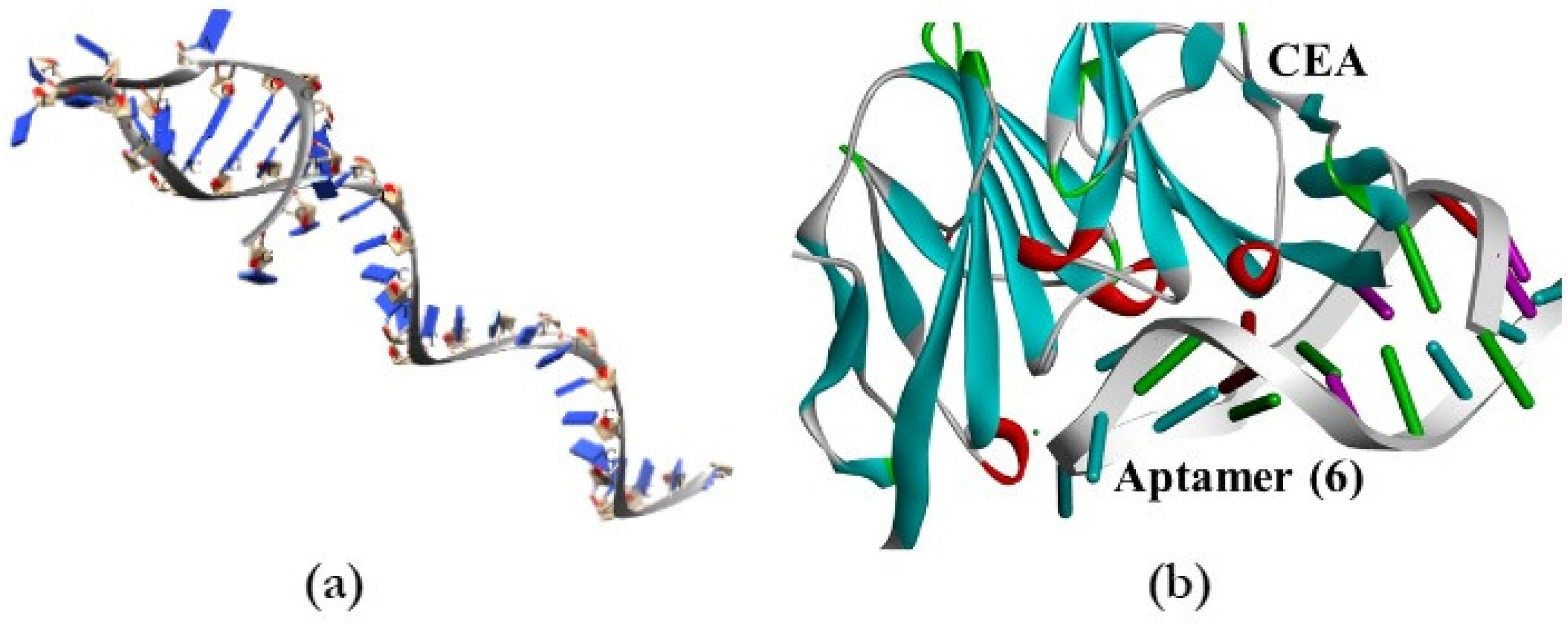{kind=link}
{kind=link}
| SELEX Cycle | Amount of Target Protein (CEA) | Amount of ssDNA | nM Ratio CEA/ssDNA | Incubation Time, min | ||
|---|---|---|---|---|---|---|
| μg/mL | nM | ng/µL | nM | |||
| 1 | 45 | 250 | 1000 | 40466 | 1/0.2 | 105 |
| 2 | 45 | 250 | 22.1 | 894.3 | 1/4 | 90 |
| 3 | 7 | 38.9 | 32.8 | 1327 | 1/34 | 60 |
| 4 | 4.9 | 27.2 | 21.9 | 886.2 | 1/32 | 60 |
| 5 | 2 | 11.1 | 14 | 566.5 | 1/50 | 45 |
| 6 | 1.5 | 8.33 | 10 | 404.7 | 1/48 | 30 |
| 7 | 1.3 | 7.22 | 8 | 323.7 | 1/45 | 25 |
| 8 | 1.5 | 8.33 | 5 | 202.3 | 1/25 | 20 |
| 9 | 1.09 | 6.06 | 4.5 | 182.1 | 1/30 | 20 |
| 10 | 0.85 | 4.72 | 4 | 161.9 | 1/35 | 15 |
| 11 | 0.65 | 3.61 | 3.5 | 141.6 | 1/40 | 10 |
| 12 | 0.47 | 2.61 | 3 | 121.4 | 1/45 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yunussova, N.; Sypabekova, M.; Zhumabekova, Z.; Matkarimov, B.; Kanayeva, D. A Novel ssDNA Aptamer Targeting Carcinoembryonic Antigen: Selection and Characterization. Biology 2022, 11, 1540. https://doi.org/10.3390/biology11101540
Yunussova N, Sypabekova M, Zhumabekova Z, Matkarimov B, Kanayeva D. A Novel ssDNA Aptamer Targeting Carcinoembryonic Antigen: Selection and Characterization. Biology. 2022; 11(10):1540. https://doi.org/10.3390/biology11101540
Chicago/Turabian StyleYunussova, Nigara, Marzhan Sypabekova, Zhazira Zhumabekova, Bakhyt Matkarimov, and Damira Kanayeva. 2022. "A Novel ssDNA Aptamer Targeting Carcinoembryonic Antigen: Selection and Characterization" Biology 11, no. 10: 1540. https://doi.org/10.3390/biology11101540