
Neurobiological Signatures of Auditory False Perception and Phantom Perception as a Consequence of Sensory Prediction Errors
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Participants
2.2. Equipment Setup
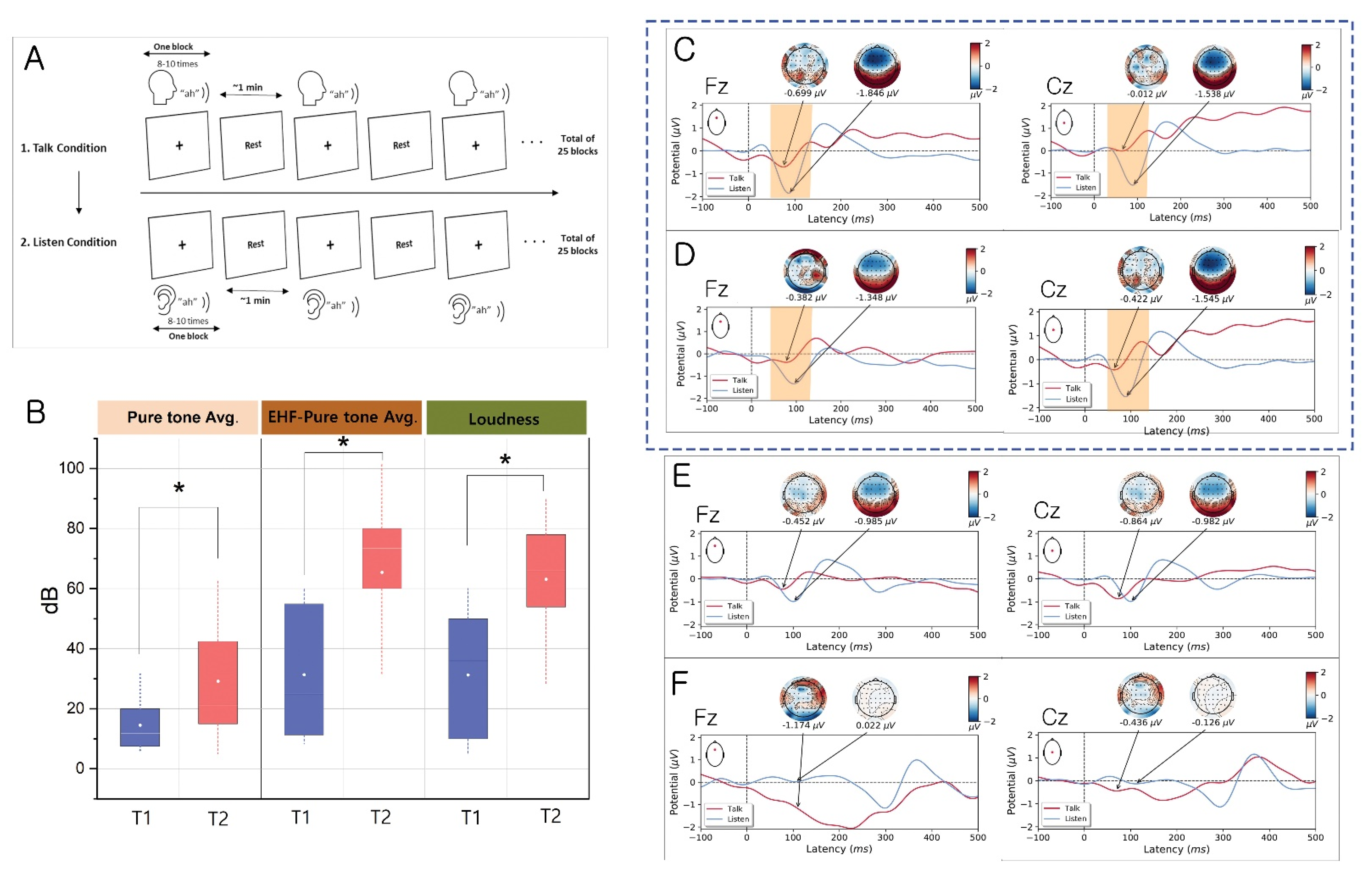
2.3. Talk/Listen Paradigm
2.4. EEG Data Acquisition and Processing
2.5. Granger Causality Analysis
2.6. Statistical Analysis
3. Results
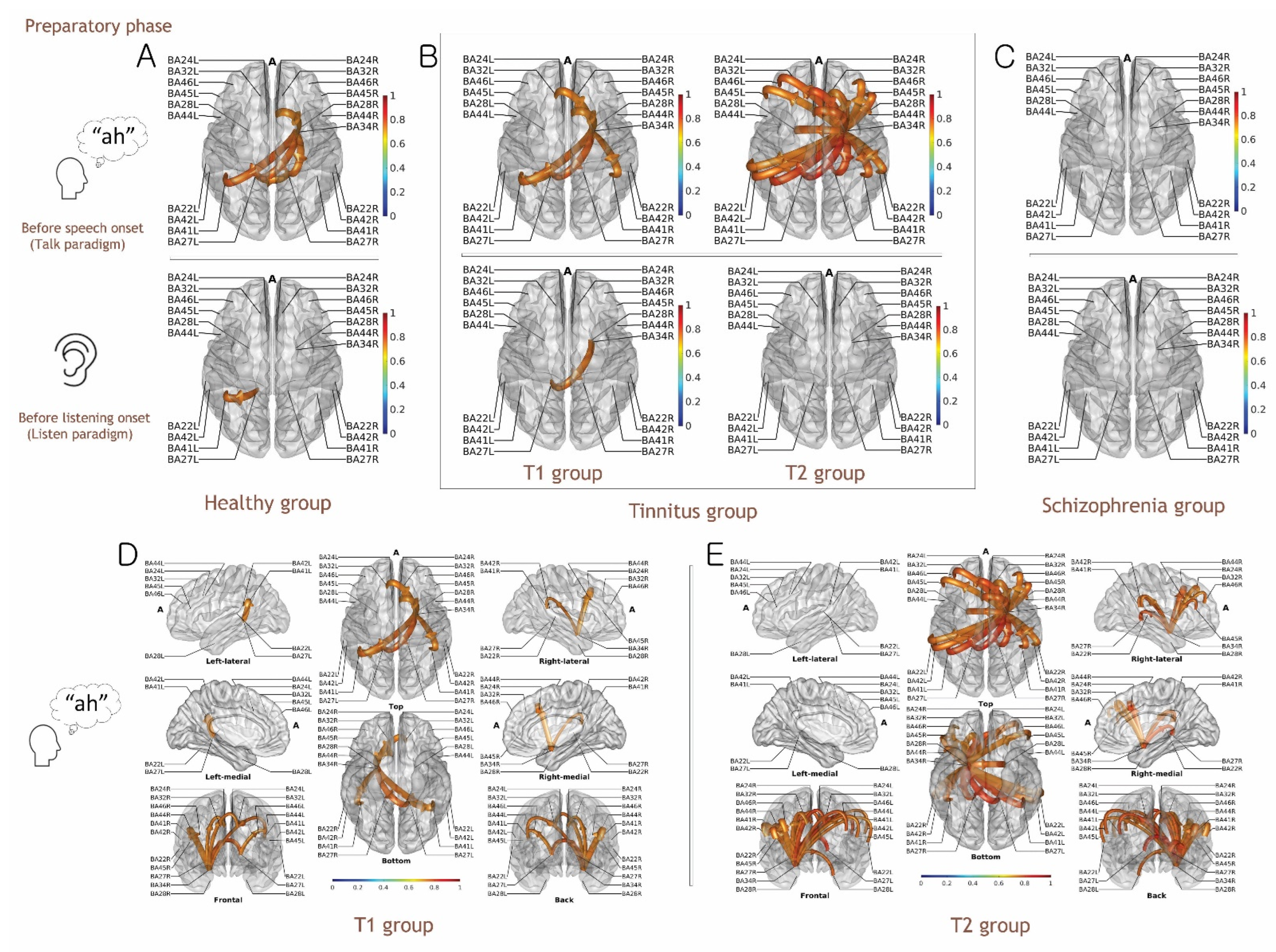
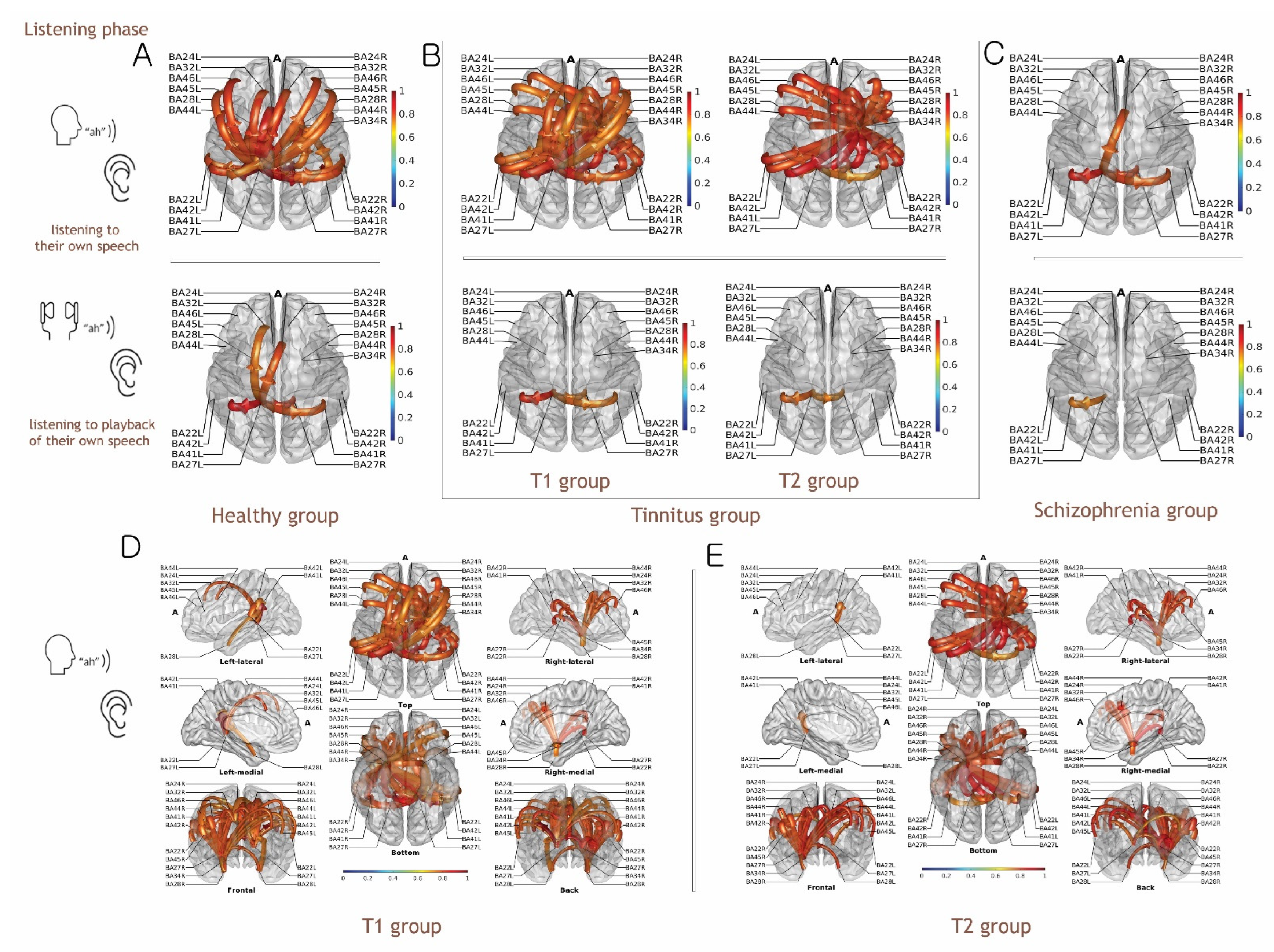
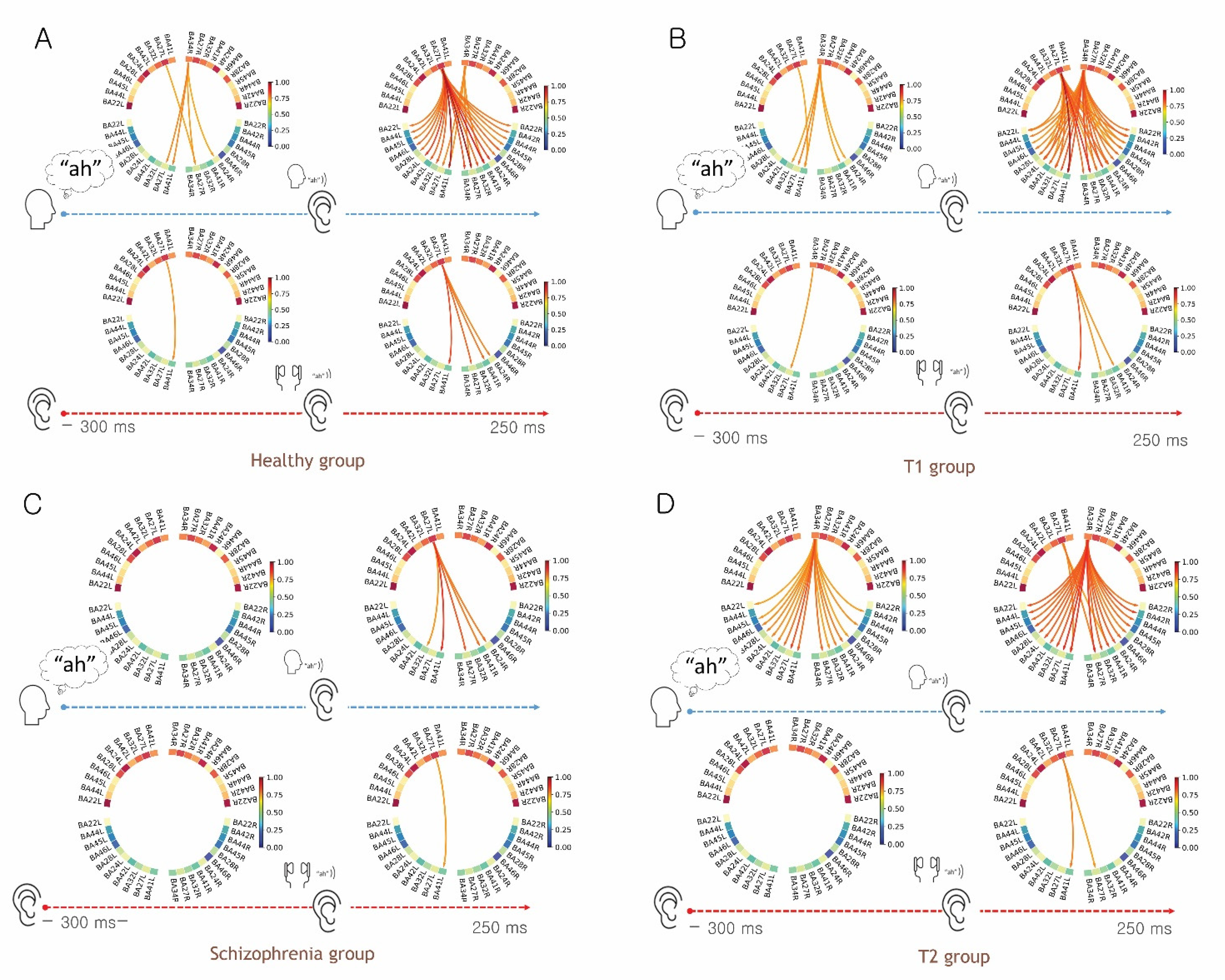
3.1. Evidence for the EC Mechanism in Normal Volunteers
3.2. Evidence for Altered EC Mechanism in the T1 Group vs. T2 Group
3.3. Evidence for Altered EC Mechanism in Schizophrenia
3.4. Functional Connectivity Analysis for the Preparatory Phase of Self-Generated Voice vs. Listening to Playback of One’s Own Voice
3.5. Functional Connectivity Analysis of Listening to Self-Generated Vocalization vs. Listening to Playback of One’s Own Voice
4. Discussion
4.1. Functional Connectivity for the Preparatory Phase of the Talk and Listen Paradigm
4.2. Functional Connectivity for Auditory Perception in the Talk and Listen Paradigm
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Penny, W. Bayesian models of brain and behaviour. Int. Sch. Res. Not. 2012, 2012, 785791. [Google Scholar] [CrossRef] [Green Version]
- Grunwald, T.; Boutros, N.N.; Pezer, N.; von Oertzen, J.; Fernández, G.; Schaller, C.; Elger, C.E. Neuronal substrates of sensory gating within the human brain. Biol. Psychiatry 2003, 53, 511–519. [Google Scholar] [CrossRef]
- Lee, S.; Kruglikov, I.; Huang, Z.J.; Fishell, G.; Rudy, B. A disinhibitory circuit mediates motor integration in the somatosensory cortex. Nat. Neurosci. 2013, 16, 1662–1670. [Google Scholar] [CrossRef] [Green Version]
- Pynn, L.K.; DeSouza, J.F. The function of efference copy signals: Implications for symptoms of schizophrenia. Vis. Res. 2013, 76, 124–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, K.E. Sensory signals during active versus passive movement. Curr. Opin. Neurobiol. 2004, 14, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.M.; Roach, B.J.; Mathalon, D.H. Assessing corollary discharge in humans using noninvasive neurophysiological methods. Nat. Protoc. 2010, 5, 1160. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.M.; Mathalon, D.H. Corollary discharge dysfunction in schizophrenia: Can it explain auditory hallucinations? Int. J. Psychophysiol. 2005, 58, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D.M.; Nelson, A.; Mooney, R. A synaptic and circuit basis for corollary discharge in the auditory cortex. Nature 2014, 513, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ffytche, D.H.; Wible, C.G. From tones in tinnitus to sensed social interaction in schizophrenia: How understanding cortical organization can inform the study of hallucinations and psychosis. Schizophr. Bull. 2014, 40, S305–S316. [Google Scholar] [CrossRef] [Green Version]
- De Ridder, D.; Vanneste, S.; Freeman, W. The Bayesian brain: Phantom percepts resolve sensory uncertainty. Neurosci. Biobehav. Rev. 2014, 44, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Hazell, J.W.; Jastreboff, P.J. Tinnitus. I: Auditory mechanisms: A model for tinnitus and hearing impairment. J. Otolaryngol. 1990, 19, 1–5. [Google Scholar] [PubMed]
- De Ridder, D.; Elgoyhen, A.B.; Romo, R.; Langguth, B. Phantom percepts: Tinnitus and pain as persisting aversive memory networks. Proc. Natl. Acad. Sci. USA 2011, 108, 8075–8080. [Google Scholar] [CrossRef] [Green Version]
- Roberts, L.E.; Husain, F.T.; Eggermont, J.J. Role of attention in the generation and modulation of tinnitus. Neurosci. Biobehav. Rev. 2013, 37, 1754–1773. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.K.; Park, S.; Ahn, M.-H.; Min, B.-K. Top-down and bottom-up neurodynamic evidence in patients with tinnitus. Hear. Res. 2016, 342, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Singla, S.; Dempsey, C.; Warren, R.; Enikolopov, A.G.; Sawtell, N.B. A cerebellum-like circuit in the auditory system cancels responses to self-generated sounds. Nat. Neurosci. 2017, 20, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malfatti, T.; Ciralli, B.; Hilscher, M.M.; Leao, R.N.; Leao, K.E. Decreasing dorsal cochlear nucleus activity ameliorates noise-induced tinnitus perception in mice. BMC Biol. 2022, 20, 102. [Google Scholar] [CrossRef]
- Han, K.H.; Mun, S.K.; Sohn, S.; Piao, X.Y.; Park, I.; Chang, M. Axonal sprouting in the dorsal cochlear nucleus affects gap-prepulse inhibition following noise exposure. Int. J. Mol. Med. 2019, 44, 1473–1483. [Google Scholar] [CrossRef] [Green Version]
- Welniarz, Q.; Worbe, Y.; Gallea, C. The forward model: A unifying theory for the role of the cerebellum in motor control and sense of agency. Front. Syst. Neurosci. 2021, 15, 644059. [Google Scholar] [CrossRef] [PubMed]
- Edition, F. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Kay, S.R.; Opler, L.A.; Lindenmayer, J.-P. The positive and negative syndrome scale (PANSS): Rationale and standardisation. Br. J. Psychiatry 1989, 155, 59–65. [Google Scholar] [CrossRef]
- Newman, C.W.; Jacobson, G.P.; Spitzer, J.B. Development of the tinnitus handicap inventory. Arch. Otolaryngol. Head Neck Surg. 1996, 122, 143–148. [Google Scholar] [CrossRef]
- Schaette, R.; McAlpine, D. Tinnitus with a normal audiogram: Physiological evidence for hidden hearing loss and computational model. J. Neurosci. 2011, 31, 13452–13457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, B. The distortion of auditory perception by sensorineural hearing impairment. Audiol. Online 2003. [Google Scholar]
- Gurgel, R.K.; Jackler, R.K.; Dobie, R.A.; Popelka, G.R. A new standardized format for reporting hearing outcome in clinical trials. Otolaryngol. Head Neck Surg. 2012, 147, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.; Shulman, A. Tinnitus evaluation. In Tinnitus Diagnosis and Treatment; Lea and Febiger: Philadelphia, PA, USA, 1991; pp. 293–318. [Google Scholar]
- Delorme, A.; Sejnowski, T.; Makeig, S. Enhanced detection of artifacts in EEG data using higher-order statistics and independent component analysis. Neuroimage 2007, 34, 1443–1449. [Google Scholar] [CrossRef] [Green Version]
- He, B.; Dai, Y.; Astolfi, L.; Babiloni, F.; Yuan, H.; Yang, L. eConnectome: A MATLAB toolbox for mapping and imaging of brain functional connectivity. J. Neurosci. Methods 2011, 195, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Zhang, W.; Dickens, D.L.; He, B. Source connectivity analysis from MEG and its application to epilepsy source localization. Brain Topogr. 2012, 25, 157–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, M.-H.; Hong, S.K.; Min, B.-K. The absence of resting-state high-gamma cross-frequency coupling in patients with tinnitus. Hear. Res. 2017, 356, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Choi, I.; Shinn-Cunningham, B. Topographic specificity of alpha power during auditory spatial attention. Neuroimage 2020, 207, 116360. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Worrell, G.A.; Lagerlund, T.D.; He, B. Ictal source analysis: Localization and imaging of causal interactions in humans. Neuroimage 2007, 34, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palus, M.; Hoyer, D. Detecting nonlinearity and phase synchronization with surrogate data. IEEE Eng. Med. Biol. Mag. 1998, 17, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Riès, S.K.; Dronkers, N.F.; Knight, R.T. Choosing words: Left hemisphere, right hemisphere, or both? Perspective on the lateralization of word retrieval. Ann. New York Acad. Sci. 2016, 1369, 111. [Google Scholar] [CrossRef]
- Frost, J.A.; Binder, J.R.; Springer, J.A.; Hammeke, T.A.; Bellgowan, P.S.; Rao, S.M.; Cox, R.W. Language processing is strongly left lateralized in both sexes: Evidence from functional MRI. Brain 1999, 122, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, A.J.; Fitzgerald, J.K.; Maimon, G. Cellular evidence for efference copy in Drosophila visuomotor processing. Nat. Neurosci. 2015, 18, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Sperry, R.W. Neural basis of the spontaneous optokinetic response produced by visual inversion. J. Comp. Physiol. Psychol. 1950, 43, 482. [Google Scholar] [CrossRef] [PubMed]
- Straka, H.; Simmers, J.; Chagnaud, B.P. A new perspective on predictive motor signaling. Curr. Biol. 2018, 28, R232–R243. [Google Scholar] [CrossRef] [Green Version]
- Ford, J.M.; Mathalon, D.H. Electrophysiological evidence of corollary discharge dysfunction in schizophrenia during talking and thinking. J. Psychiatr. Res. 2004, 38, 37–46. [Google Scholar] [CrossRef]
- Heinks-Maldonado, T.H.; Mathalon, D.H.; Houde, J.F.; Gray, M.; Faustman, W.O.; Ford, J.M. Relationship of imprecise corollary discharge in schizophrenia to auditory hallucinations. Arch. Gen. Psychiatry 2007, 64, 286–296. [Google Scholar] [CrossRef] [Green Version]
- Boutros, N.N.; Korzyukov, O.; Jansen, B.; Feingold, A.; Bell, M. Sensory gating deficits during the mid-latency phase of information processing in medicated schizophrenia patients. Psychiatry Res. 2004, 126, 203–215. [Google Scholar] [CrossRef]
- Jastreboff, P.J. Phantom auditory perception (tinnitus): Mechanisms of generation and perception. Neurosci. Res. 1990, 8, 221–254. [Google Scholar] [CrossRef]
- van de Ven, V.; Waldorp, L.; Christoffels, I. Hippocampus plays a role in speech feedback processing. NeuroImage 2020, 223, 117319. [Google Scholar] [CrossRef]
- Stachenfeld, K.L.; Botvinick, M.M.; Gershman, S.J. The hippocampus as a predictive map. Nat. Neurosci. 2017, 20, 1643. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.; Buzsáki, G. The functional anatomy of time: What and when in the brain. Trends Cogn. Sci. 2016, 20, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Mu, P.; Ploog, D. Inhibition of auditory cortical neurons during phonation. Brain Res. 1981, 215, 61–76. [Google Scholar]
- Creutzfeldt, O.; Ojemann, G.; Lettich, E. Neuronal activity in the human lateral temporal lobe. Exp. Brain Res. 1989, 77, 451–475. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.Z.; Munhall, K.G.; Johnsrude, I.S. Functional overlap between regions involved in speech perception and in monitoring one's own voice during speech production. J. Cogn. Neurosci. 2010, 22, 1770–1781. [Google Scholar] [CrossRef] [Green Version]
- Tourville, J.A.; Reilly, K.J.; Guenther, F.H. Neural mechanisms underlying auditory feedback control of speech. Neuroimage 2008, 39, 1429–1443. [Google Scholar] [CrossRef] [Green Version]
- McCormick, D.A.; Bal, T. Sensory gating mechanisms of the thalamus. Curr. Opin. Neurobiol. 1994, 4, 550–556. [Google Scholar] [CrossRef]
- Cromwell, H.C.; Mears, R.P.; Wan, L.; Boutros, N.N. Sensory gating: A translational effort from basic to clinical science. Clin. EEG Neurosci. 2008, 39, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Darvas, F.; Ermer, J.J.; Mosher, J.C.; Leahy, R.M. Generic head models for atlas-based EEG source analysis. Hum. Brain Mapp. 2006, 27, 129–143. [Google Scholar] [CrossRef]
- Michel, C.M.; Murray, M.M.; Lantz, G.; Gonzalez, S.; Spinelli, L.; de Peralta, R.G. EEG source imaging. Clin. Neurophysiol. 2004, 115, 2195–2222. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Tinnitus | Schizophrenia | Controls | |
|---|---|---|---|---|
| T1 | T2 | |||
| Total Number (n) | 8 | 15 | 10 | 23 |
| Gender (female/male) | 2 /6 | 6/9 | 7/3 | 12/11 |
| Age ± SD | 39.12 ± 12.59 | 40.40 ± 13.50 | 30.10 ± 6.00 | 34.76 ± 7.40 |
| Pure tone average * (dB) for the right Ear | 12.03 ± 7.70 | 27.33 ± 20.11 | 7.87 ± 9.82 | 5.27 ± 3.40 |
| High frequency pure tone average ** (dB) for the right ear | 29.68 ± 19.93 | 57.94 ± 28.85 | ||
| Pure tone average (dB) for the left ear * | 11.56 ± 9.08 | 20.25 ± 15.81 | 4.12 ± 2.63 | 4.13 ± 3.90 |
| High frequency pure tone average (dB) for the left ear | 23.85 ± 21.57 | 46.94 ± 13.50 | ||
| Pure tone average (dB) for the involved side * | 14.50 ± 10.08 | 29.10 ± 19.16 | ||
| High frequency pure tone average (dB) for the involved side | 31.35 ± 22.07 | 65.38 ± 26.6 | ||
| Tinnitus side (n) | ||||
| Right/Left/Both | 1/3/4 | 10/2/3 | ||
| Pitch in kHz | 6.00 ± 2.83 | 5.25 ± 2.98 | ||
| Loudness in dB HL | 31.28 ± 20.46 | 63.08 ± 20.35 | ||
| THI score | 57.00 ± 25.16 | 42.93 ± 18.39 | ||
| PANSS score | 52.70 ± 14.23 | |||
| Hallucinatory behavior score on PANSS | 2.30 ± 1.25 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, M.-H.; Alsabbagh, N.; Lee, H.-J.; Kim, H.-J.; Jung, M.-H.; Hong, S.-K. Neurobiological Signatures of Auditory False Perception and Phantom Perception as a Consequence of Sensory Prediction Errors. Biology 2022, 11, 1501. https://doi.org/10.3390/biology11101501
Ahn M-H, Alsabbagh N, Lee H-J, Kim H-J, Jung M-H, Hong S-K. Neurobiological Signatures of Auditory False Perception and Phantom Perception as a Consequence of Sensory Prediction Errors. Biology. 2022; 11(10):1501. https://doi.org/10.3390/biology11101501
Chicago/Turabian StyleAhn, Min-Hee, Nour Alsabbagh, Hyo-Jeong Lee, Hyung-Jong Kim, Myung-Hun Jung, and Sung-Kwang Hong. 2022. "Neurobiological Signatures of Auditory False Perception and Phantom Perception as a Consequence of Sensory Prediction Errors" Biology 11, no. 10: 1501. https://doi.org/10.3390/biology11101501