
Biological Response of Planktic Foraminifera to Decline in Seawater pH

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Seawater Preparation
2.3. Incubator Parameter Setting
2.4. Individual Experiment Setting
2.5. Community Experiment Setting
2.6. Shell Analysis
2.7. Statistical Analyses
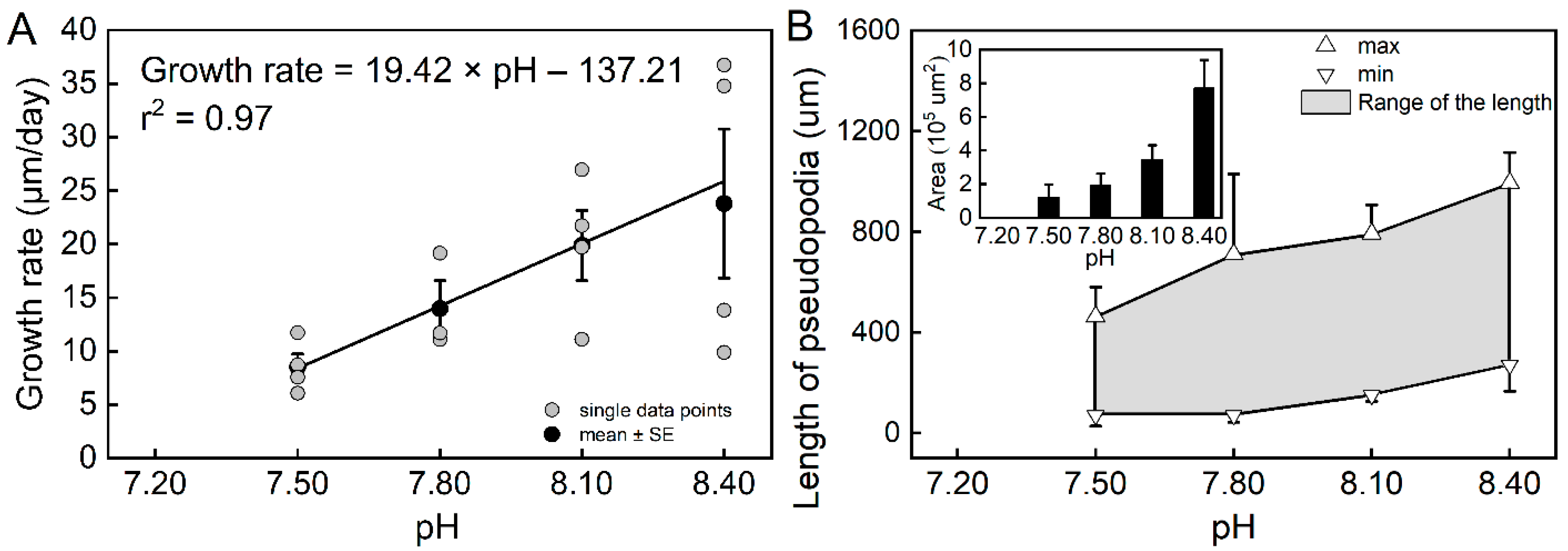
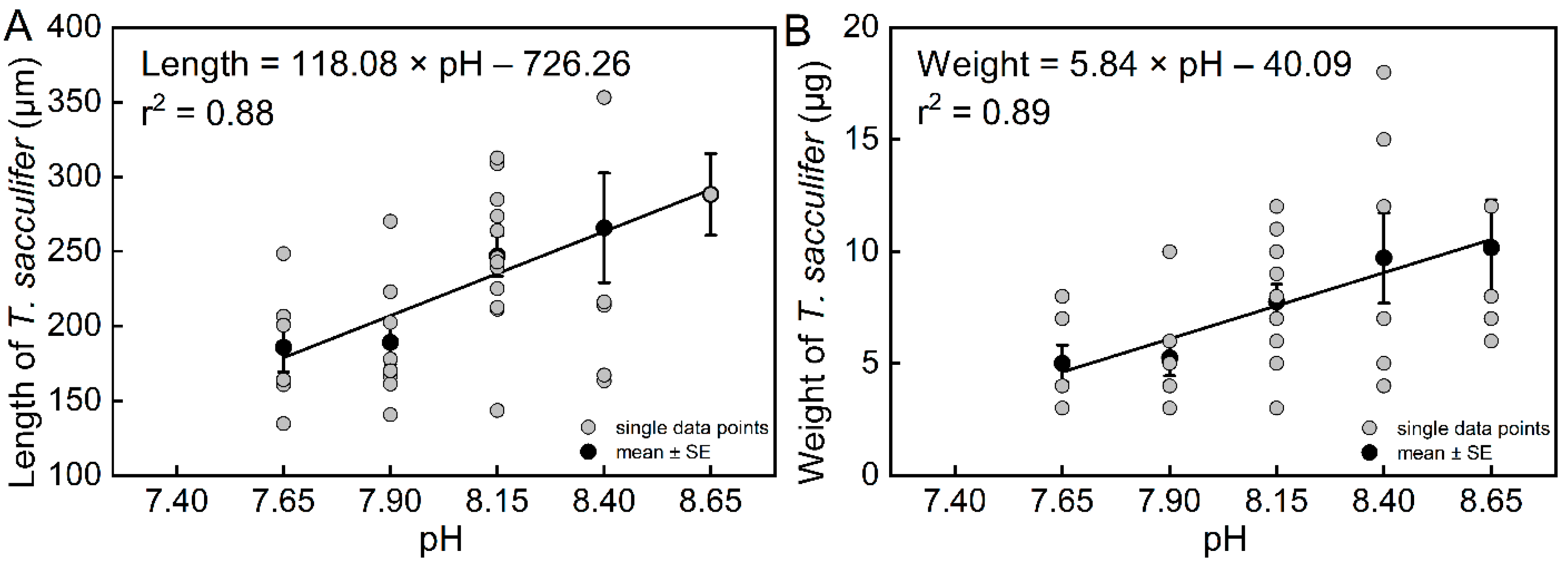
3. Results
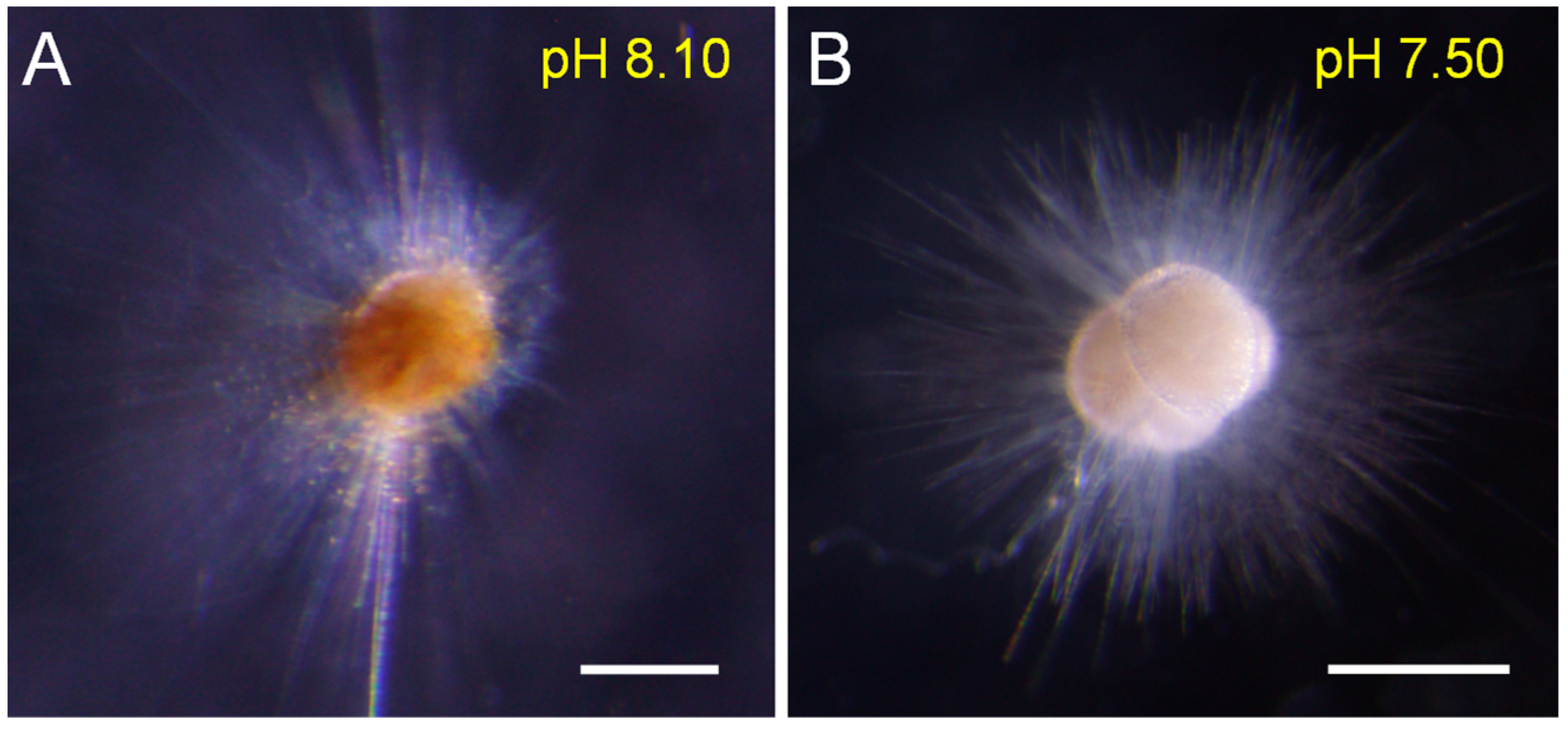
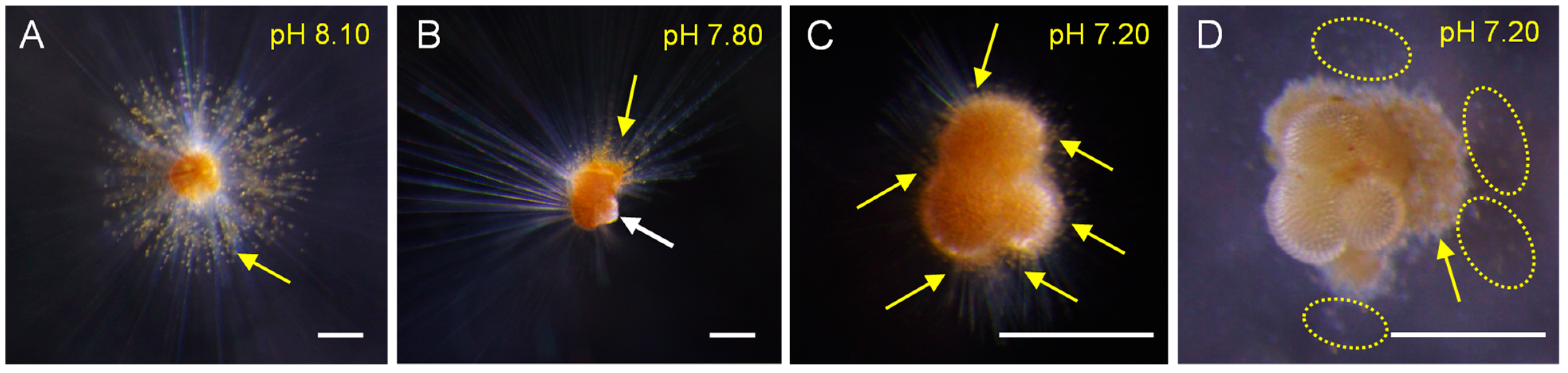
3.1. Biological Parameters from Individual Experiments
3.2. Biological Parameters from the Community Experiment
4. Discussion
4.1. Biological Response of T. sacculifer to pH Decline
4.2. Paleoceanographic Significance of the Biological Responses of Symbiont-Bearing Calcifiers to Seawater pH
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, C.V.; Rivest, E.B.; Hill, T.M.; Gaylord, B.; Russell, A.D.; Sanford, E. Ocean acidification compromises a planktic calcifier with implications for global carbon cycling. Sci. Rep. 2017, 7, 2225. [Google Scholar] [CrossRef] [PubMed]
- Rivest, E.B.; Kelly, M.W.; DeBiasse, M.B.; Hofmann, G.E. Host and symbionts in Pocillopora damicornis larvae display different transcriptomic responses to ocean acidification and warming. Front. Mar. Sci. 2018, 5, 186. [Google Scholar] [CrossRef]
- Hönisch, B.; Ridgwell, A.; Schmidt, D.N.; Thomas, E.; Gibbs, S.J.; Sluijs, A.; Zeebe, R.; Kump, L.; Martindale, R.C.; Greene, S.E.; et al. The geological record of ocean acidification. Science 2012, 335, 1058–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelejero, C.; Calvo, E.; Hoegh-Guldberg, O. Paleo-perspectives on ocean acidification. Trends Ecol. Evol. 2010, 25, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Zachos, J.C.; Röhl, U.; Schellenberg, S.A.; Sluijs, A.; Hodell, D.A.; Kelly, D.C.; Thomas, E.; Nicolo, M.; Raffi, I.; Lourens, L.J.; et al. Rapid acidification of the ocean during the Paleocene-Eocene thermal maximum. Science 2005, 308, 1611–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moy, A.D.; Howard, W.R.; Bray, S.G.; Trull, T.W. Reduced calcification in modern Southern Ocean planktonic foraminifera. Nat. Geosci. 2009, 2, 276–280. [Google Scholar] [CrossRef]
- Henehan, M.J.; Evans, D.; Shankle, M.; Burke, J.E.; Foster, G.L.; Anagnostou, E.; Chalk, T.B.; Stewart, J.A.; Alt, C.H.S.; Durrant, J.; et al. Size-dependent response of foraminiferal calcification to seawater carbonate chemistry. Biogeosciences 2017, 14, 3287–3308. [Google Scholar] [CrossRef] [Green Version]
- Lombard, F.; da Rocha, R.E.; Bijma, J.; Gattuso, J.P. Effect of carbonate ion concentration and irradiance on calcification in planktonic foraminifera. Biogeosciences 2010, 7, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Haynes, L.L.; Hönisch, B.; Dyez, K.A.; Holland, K.; Rosenthal, Y.; Fish, C.R.; Subhas, A.V.; Rae, J.W.B. Calibration of the B/Ca proxy in the planktic foraminifer Orbulina universa to Paleocene seawater conditions. Paleoceanography 2017, 32, 580–599. [Google Scholar] [CrossRef] [Green Version]
- Henehan, M.J.; Foster, G.L.; Bostock, H.C.; Greenop, R.; Marshall, B.J.; Wilson, P.A. A new boron isotope-pH calibration for Orbulina universa, with implications for understanding and accounting for ‘vital effects’. Earth Planet. Sci. Lett. 2016, 454, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, A.; Bijma, J.; Spero, H.; Lea, D.W. Empirical relationship between pH and the boron isotopic composition of Globigerinoides sacculifer: Implications for the boron isotope paleo-pH proxy. Paleoceanography 2001, 16, 515–519. [Google Scholar] [CrossRef] [Green Version]
- Russell, A.D.; Hönisch, B.; Spero, H.J.; Lea, D.W. Effects of seawater carbonate ion concentration and temperature on shell U, Mg, and Sr in cultured planktonic foraminifera. Geochim. Cosmochim. Acta 2004, 68, 4347–4361. [Google Scholar] [CrossRef]
- Foster, G.L. Seawater pH, pCO2 and [CO32−] variations in the Caribbean Sea over the last 130 kyr: A boron isotope and B/Ca study of planktic foraminifera. Earth Planet. Sci. Lett. 2008, 271, 254–266. [Google Scholar] [CrossRef]
- Avnaim-Katav, S.; Herut, B.; Rahav, E.; Katz, T.; Weinstein, Y.; Alkalay, R.; Berman-Frank, I.; Zlatkin, O.; Almogi-Labin, A. Sediment trap and deep sea coretop sediments as tracers of recent changes in planktonic foraminifera assemblages in the southeastern ultra-oligotrophic Levantine Basin. Deep-Sea Res. II 2020, 171, 104669. [Google Scholar] [CrossRef]
- Michaels, A.F.; Caron, D.A.; Swanberg, N.R.; Howse, F.A.; Michaels, C.M. Planktonic sarcodines (Acantharia, Radiolaria, Foraminifera) in surface waters near Bermuda: Abundance, biomass and vertical flux. J. Plankton Res. 1995, 17, 131–163. [Google Scholar] [CrossRef]
- Boudagher-Fadel, M.K. Biostratigraphic and Geological Significance of Planktonic Foraminifera; UCL Press: London, UK, 2015; p. 298. [Google Scholar]
- Bé, A.W.H.; Spero, H.J.; Anderson, O.R. Effects of symbiont elimination and reinfection on the life processes of the planktonic foraminifer Globigerinoides sacculifer. Mar. Biol. 1982, 70, 73–86. [Google Scholar] [CrossRef]
- Barker, J.B.; Jonathan, E.; Peter, R.; Yehuda, C. Symbiotic photosynthesis in a planktonic foraminiferan, Globeriginoides sacculifer (Brady), studied with microelectrodes. Limnol. Oceanogr. 1985, 30, 1253–1267. [Google Scholar] [CrossRef]
- Schiebel, R.; Hemleben, C. Planktic Foraminifers in the Modern Ocean; Springer: Berlin/Heidelberg, Germany, 2017; p. 358. [Google Scholar]
- Palmer, M.R.; Pearson, P.N.; Cobb, S.J. Reconstructing past ocean pH-depth profiles. Science 1998, 282, 1468–1471. [Google Scholar] [CrossRef]
- Beer, C.J.; Schiebel, R.; Wilson, P.A. Technical note: On methodologies for determining the size-normalised weight of planktic foraminifera. Biogeosciences 2010, 7, 2193–2198. [Google Scholar] [CrossRef] [Green Version]
- Gerasimova, A.V.; Maximovich, N.V.; Filippova, N.A. Cohort life tables for a population of the soft-shell clam, Mya arenaria L., in the White Sea. Helgol. Mar. Res. 2015, 69, 147–158. [Google Scholar] [CrossRef]
- Bijma, J.; Faber, W.W.; Hemleben, C. Temperature and salinity limits for growth and survival of some planktonic foraminifers in laboratory cultures. J. Foraminifer. Res. 1990, 20, 95–116. [Google Scholar] [CrossRef] [Green Version]
- Lombard, F.; Erez, J.; Michel, E.; Labeyrie, L. Temperature effect on respiration and photosynthesis of the symbiont-bearing planktonic foraminifera Globigerinoides ruber, Orbulina universa, and Globigerinella siphonifera. Limnol. Oceanogr. 2009, 54, 210–218. [Google Scholar] [CrossRef]
- Hecht, A.D. Size variations in planktonic foraminifera: Implications for quantitative paleoclimatic analysis. Science 1976, 192, 1330–1332. [Google Scholar] [CrossRef] [PubMed]
- Todd, C.L.; Schmidt, D.N.; Robinson, M.M.; De Schepper, S. Planktic foraminiferal test size and weight response to the late Pliocene environment. Paleoceanogr. Paleoclimatol. 2020, 35, e2019PA003738. [Google Scholar] [CrossRef] [Green Version]
- Spalding, C.; Finnegan, S.; Fischer, W.W. Energetic costs of calcification under ocean acidification. Glob. Biogeochem. Cycles 2017, 31, 866–877. [Google Scholar] [CrossRef] [Green Version]
- Beer, C.J.; Schiebel, R.; Wilson, P.A. Testing planktic foraminiferal shell weight as a surface water [CO32−] proxy using plankton net samples. Geology 2010, 38, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Fox, L.; Stukins, S.; Hill, T.; Miller, C.G. Quantifying the effect of anthropogenic climate change on calcifying plankton. Sci. Rep. 2020, 10, 1620. [Google Scholar] [CrossRef] [Green Version]
- Hemleben, C.; Spindler, M.; Anderson, O. Modern Planktonic Foraminifera; Springer: New York, NY, USA, 1989; p. 363. [Google Scholar]
- Fehrenbacher, J.S.; Russell, A.D.; Davis, C.V.; Gagnon, A.C.; Spero, H.J.; Cliff, J.B.; Zhu, Z.; Martin, P. Link between light-triggered Mg-banding and chamber formation in the planktic foraminifera Neogloboquadrina dutertrei. Nat. Commun. 2017, 8, 15441. [Google Scholar] [CrossRef]
- von Langen, P.J.; Pak, D.K.; Spero, H.J.; Lea, D.W. Effects of temperature on Mg/Ca in neogloboquadrinid shells determined by live culturing. Geochem. Geophys. Geosyst. 2005, 6, Q10P03. [Google Scholar] [CrossRef]
- Schmidt, C.; Heinz, P.; Kucera, M.; Uthicke, S. Temperature-induced stress leads to bleaching in larger benthic foraminifera hosting endosymbiotic diatoms. Limnol. Oceanogr. 2011, 56, 1587–1602. [Google Scholar] [CrossRef]
- Sully, S.; van Woesik, R. Turbid reefs moderate coral bleaching under climate-related temperature stress. Glob. Chang. Biol. 2020, 26, 1367–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sully, S.; Burkepile, D.E.; Donovan, M.K.; Hodgson, G.; van Woesik, R. A global analysis of coral bleaching over the past two decades. Nat. Commun. 2019, 10, 1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.E.; Hallock, P. Bleaching in Amphistegina gibbosa d’Orbigny (Class Foraminifera): Observations from laboratory experiments using visible and ultraviolet light. Mar. Biol. 2004, 145, 641–649. [Google Scholar] [CrossRef]
- Henehan, M.J.; Ridgwell, A.; Thomas, E.; Zhang, S.; Alegret, L.; Schmidt, D.N.; Rae, J.W.B.; Witts, J.D.; Landman, N.H.; Greene, S.E.; et al. Rapid ocean acidification and protracted Earth system recovery followed the end-Cretaceous Chicxulub impact. Proc. Natl. Acad. Sci. USA 2019, 116, 22500–22504. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, B.P.; Middelburg, J.J.; Sluijs, A.; van der Ploeg, R. Secular variations in the carbonate chemistry of the oceans over the Cenozoic. Earth Planet. Sci. Lett. 2019, 512, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Bé, A.W.H.; Anderson, O.R.; Faber, W.W.; Caron, D.A. Sequence of morphological and cytoplasmic changes during gametogenesis in the planktonic foraminifer Globigerinoides sacculifer (Brady). Micropaleontology 1983, 29, 310–325. [Google Scholar] [CrossRef]
- de Nooijer, L.J.; Toyofuku, T.; Kitazato, H. Foraminifera promote calcification by elevating their intracellular pH. Proc. Natl. Acad. Sci. USA 2009, 106, 15374–15378. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
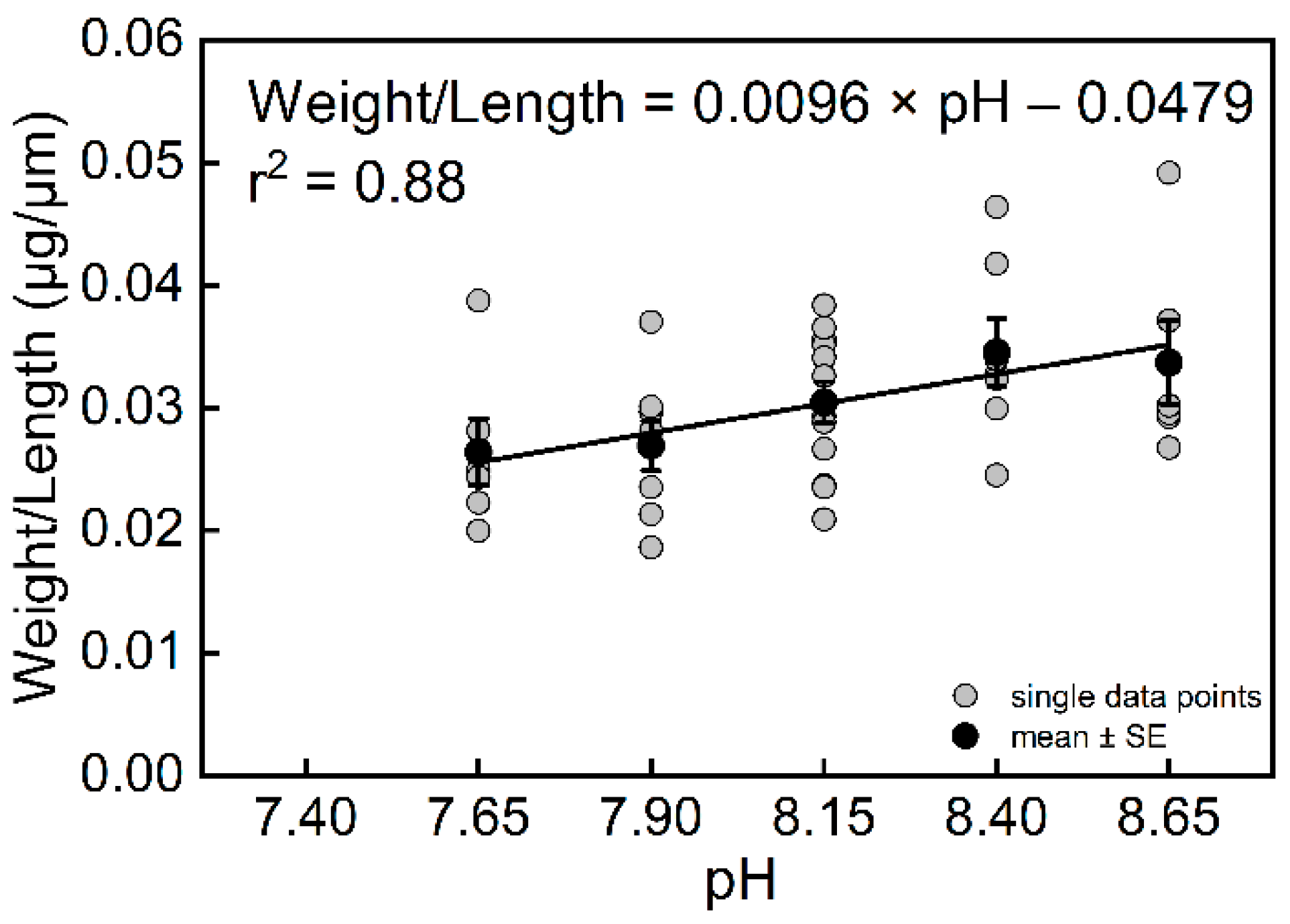{kind=link}
| Trilobatus sacculifer | r2 | p-Value | Number of Specimens | Equations | |
|---|---|---|---|---|---|
| Growth rate (G) | pH | 0.97 | 0.0147 | 23 | pH = (G + 137.21 (±18.20))/(19.42 ± 2.38) |
| Length (L) | 0.88 | 0.0181 | 39 | pH = (L + 726.26 (±201.19))/(118.08 ± 25.08) | |
| Weight (W) | 0.89 | 0.0159 | 39 | pH = (W + 40.09 (±9.40))/(5.84 ± 1.18) | |
| Weight/Length (W/L) | 0.88 | 0.0186 | 39 | pH = (W/L + 0.0096 (±0.0021))/(0.0479 ± 0.0167) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, S.; Lei, Y.; Bi, H.; Xu, K.; Li, T.; Jian, Z. Biological Response of Planktic Foraminifera to Decline in Seawater pH. Biology 2022, 11, 98. https://doi.org/10.3390/biology11010098
Dong S, Lei Y, Bi H, Xu K, Li T, Jian Z. Biological Response of Planktic Foraminifera to Decline in Seawater pH. Biology. 2022; 11(1):98. https://doi.org/10.3390/biology11010098
Chicago/Turabian StyleDong, Shuaishuai, Yanli Lei, Hongsheng Bi, Kuidong Xu, Tiegang Li, and Zhimin Jian. 2022. "Biological Response of Planktic Foraminifera to Decline in Seawater pH" Biology 11, no. 1: 98. https://doi.org/10.3390/biology11010098