
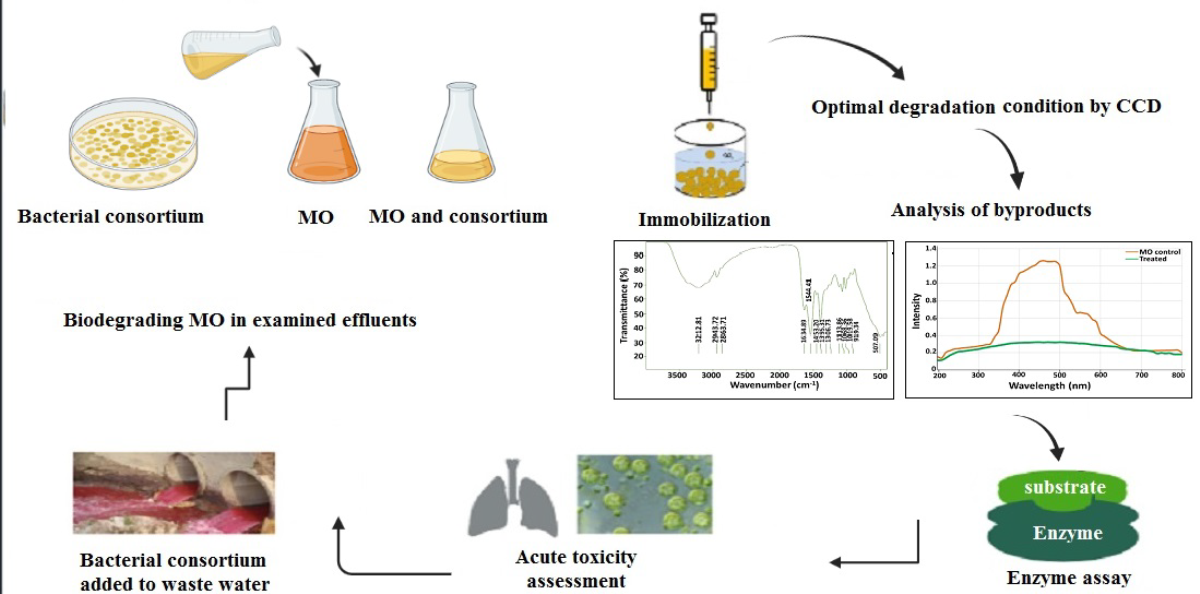
Methyl Orange Biodegradation by Immobilized Consortium Microspheres: Experimental Design Approach, Toxicity Study and Bioaugmentation Potential


 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Screening, Selection and Growth Condition Analysis of MO-Degrading Microorganisms
2.2. Antagonism Assay for Detecting the Compatibility of the Consortium Strains
2.3. Inoculum Preparation and Consortium Immobilization in Alginate Microspheres
2.4. Characterization of Immobilized Beads by Scanning Electron Microscopy (SEM)
2.5. Biodegradation Test of Immobilized Beads
2.6. MO Degradation by Consortium Immobilized in Alginate Beads Using Central Composite Design (CCD)
2.7. Statistical Analysis
2.8. Decolorization Assay of MO
2.8.1. Extraction and Analysis of Degradation By-Products
2.8.2. UV−Visible Spectrophotometric Analysis
2.8.3. FTIR
2.9. Enzymes Activity Assays
2.10. Protein Content Determination
2.11. Acute Toxicity Assessments
2.11.1. Phytotoxicity Bioassays with Chlorella vulgaris
2.11.2. Determination of Cytotoxicity against Normal Lung and Kidney Cell Lines
2.12. Bioaugmentation of Free and Immobilized Cells in Real Waste Effluents
3. Results
3.1. Selection of MO-Degrading Microorganisms and Their Compatibility
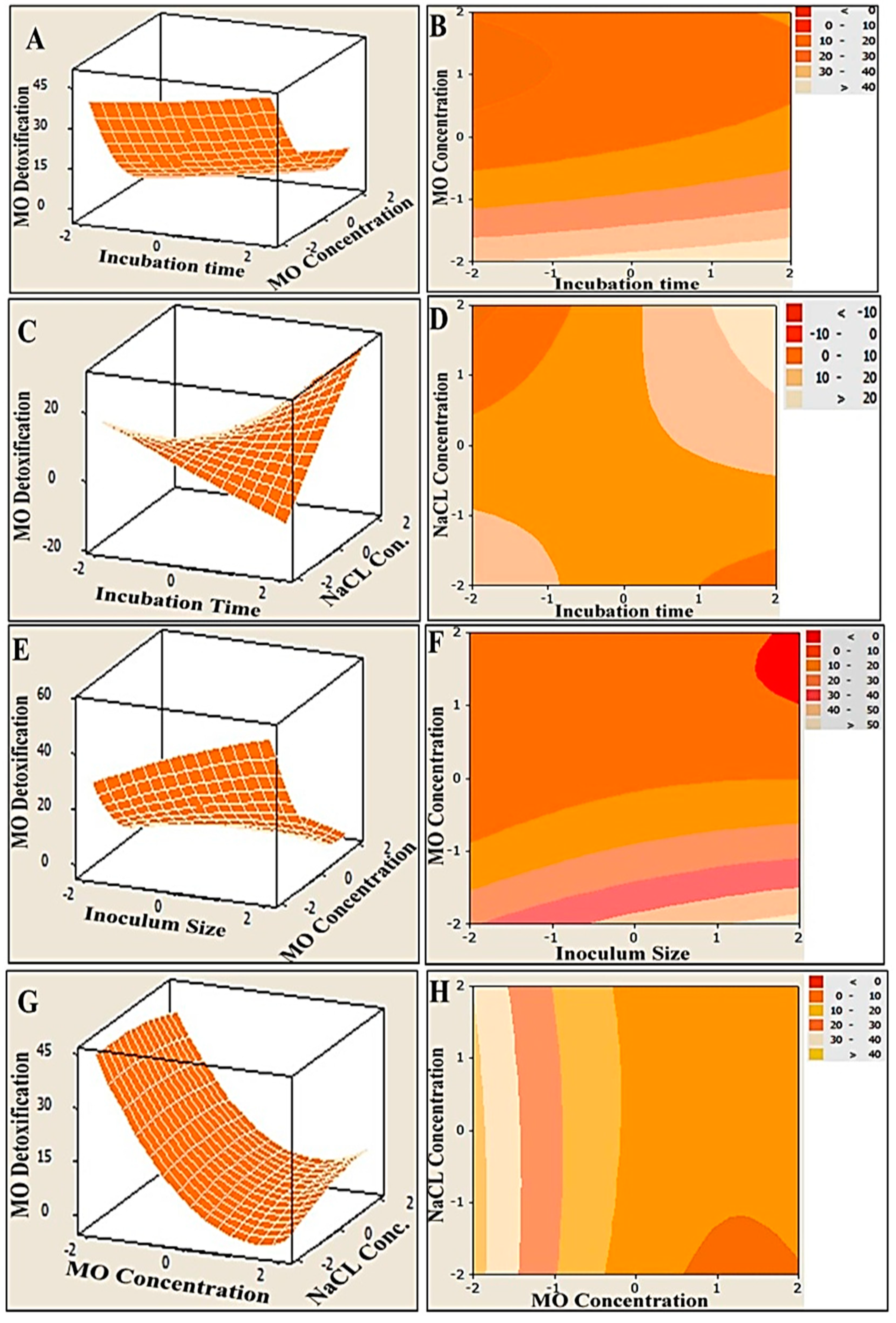
3.2. MO Degradation by Consortium Immobilized in Alginate Beads Using Central Composite Design (CCD)
3.3. Multiple Regression Analysis and ANOVA
3.4. Graphical Interpretation of the Response Surface Model
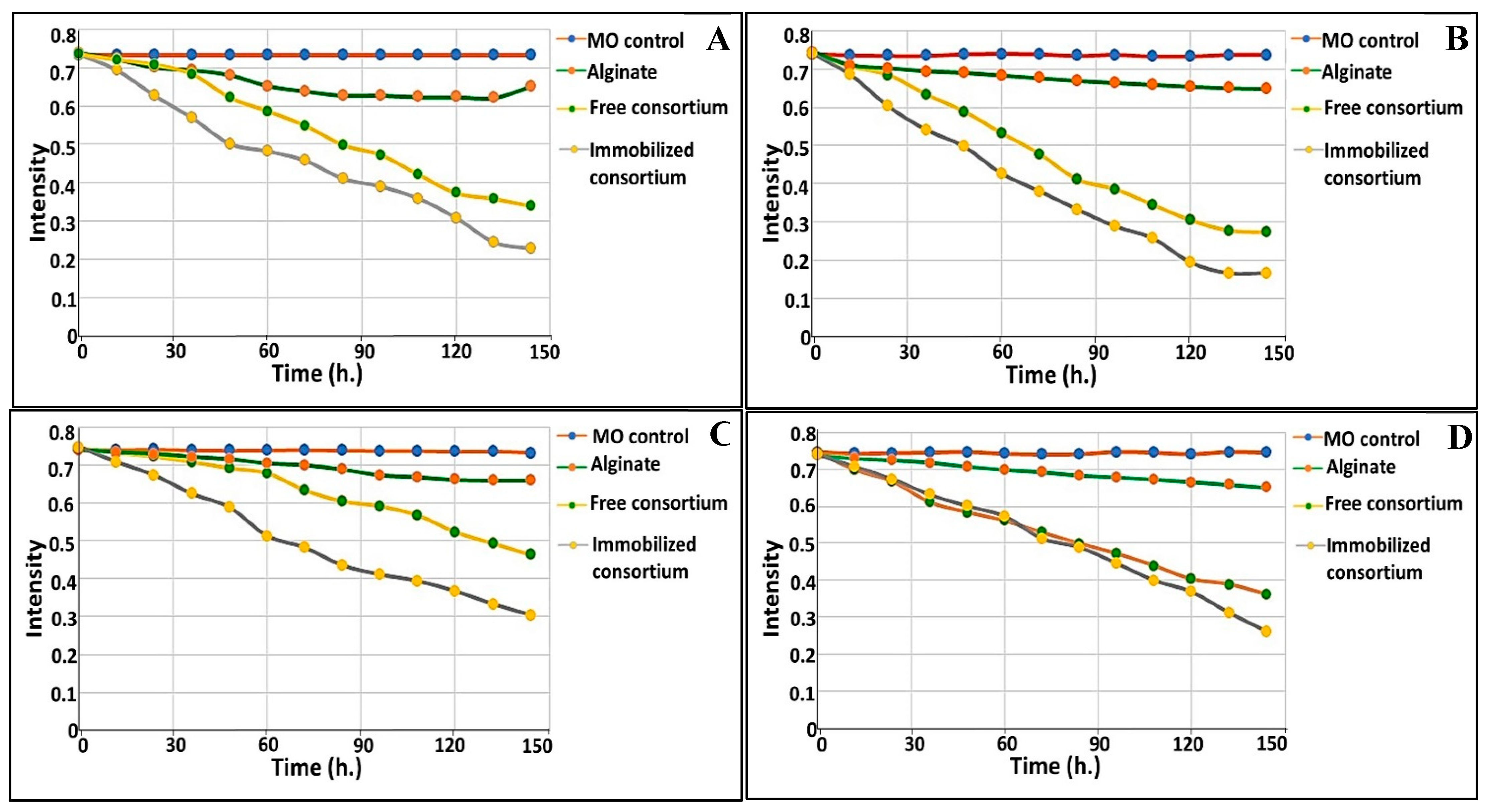
3.5. Analysis of Degradation By-Products
3.5.1. UV−Visible Spectrophotometric Analysis
3.5.2. FTIR Analysis
3.6. Enzymes Activity Assays
3.7. Acute Toxicity Assessment
3.7.1. Phytotoxicity Bioassays with C. vulgaris
3.7.2. Cytotoxicity against Normal Human and Animal Cell Lines
3.8. Bioaugmentation of Free and Immobilized Cells in Real Waste Effluents
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Asad, S.; Amoozegar, M.A.; Pourbabaee, A.; Sarbolouki, M.; Dastgheib, S. Decolorization of textile azo dyes by newly isolated halophilic and halotolerant bacteria. Bioresour. Technol. 2007, 98, 2082–2088. [Google Scholar] [CrossRef]
- El-Sayed, G.O.; Hazaa, M.M.; El-Komy, A.M. Biotreatment of water polluted with methyl orange dye by using different forms of yeast. J. Basic Environ. Sci. 2018, 5, 217–221. [Google Scholar]
- Mishra, S.; Nayak, J.K.; Maiti, A. Bacteria-mediated bio-degradation of reactive azo dyes coupled with bio-energy generation from model wastewater. Clean Technol. Environ. Policy 2020, 22, 651–667. [Google Scholar] [CrossRef]
- Sinha, S.; Chattopadhyay, P.; Pan, I.; Chatterjee, S.; Chanda, P.; Bandyopadhyay, D.; Das, K.; Sen, S. Microbial transformation of xenobiotics for environmental bioremediation. Afr. J. Biotechnol. 2011, 8, 6016–6027. [Google Scholar] [CrossRef]
- Fernandez, M.; Paisio, C.E.; Perotti, R.; Pereira, P.P.; Agostini, E.; González, P.S. Laboratory and field microcosms as useful experimental systems to study the bioaugmentation treatment of tannery effluents. J. Environ. Manag. 2019, 234, 503–511. [Google Scholar] [CrossRef]
- Gaur, N.; Narasimhulu, K.; PydiSetty, Y. Recent advances in the bio-remediation of persistent organic pollutants and its effect on environment. J. Clean. Prod. 2018, 198, 1602–1631. [Google Scholar] [CrossRef]
- Sayed, K.; Baloo, L.; Sharma, N. Bioremediation of Total Petroleum Hydrocarbons (TPH) by Bioaugmentation and Biostimulation in Water with Floating Oil Spill Containment Booms as Bioreactor Basin. Int. J. Environ. Res. Public Health 2021, 18, 2226. [Google Scholar] [CrossRef] [PubMed]
- Magan, N.; Fragoeiro, S.; Bastos, A.C. Environmental Factors and Bioremediation of Xenobiotics Using White Rot Fungi. Mycobiology 2010, 38, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Herrero, M.; Stuckey, D. Bioaugmentation and its application in wastewater treatment: A review. Chemosphere 2015, 140, 119–128. [Google Scholar] [CrossRef]
- Raper, E.; Stephenson, T.; Anderson, D.; Fisher, R.; Soares, A. Industrial wastewater treatment through bioaugmentation. Process. Saf. Environ. Prot. 2018, 118, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Basturk, I.; Varank, G.; Murat-Hocaoglu, S.; Yazici-Guvenc, S.; Oktem-Olgun, E.E.; Canli, O. Characterization and Treatment of Medical Laboratory Wastewater by Ozonation: Optimization of Toxicity Removal by Central Composite Design. Ozone Sci. Eng. 2021, 43, 213–227. [Google Scholar] [CrossRef]
- Costa, A.F.S.; Albuquerque, C.D.C.; Salgueiro, A.A.; Sarubbo, L.A. Color removal from industrial dyeing and laundry effluent by microbial consortium and coagulant agents. Process. Saf. Environ. Prot. 2018, 118, 203–210. [Google Scholar] [CrossRef]
- Asfaram, A.; Ghaedi, M.; Dashtian, K.; Ghezelbash, G.R. Preparation and Characterization of Mn0.4Zn0.6Fe2O4 Nanoparticles Supported on Dead Cells of Yarrowia lipolytica as a Novel and Efficient Adsorbent/Biosorbent Composite for the Removal of Azo Food Dyes: Central Composite Design Optimization Study. ACS Sustain. Chem. Eng. 2018, 6, 4549–4563. [Google Scholar] [CrossRef]
- Machrouhi, A.; Alilou, H.; Farnane, M.; El Hamidi, S.; Sadiq, M.; Abdennouri, M.; Tounsadi, H.; Barka, N. Statistical optimization of activated carbon from Thapsia transtagana stems and dyes removal efficiency using central composite design. J. Sci. Adv. Mater. Devices 2019, 4, 544–553. [Google Scholar] [CrossRef]
- Azargohar, R.; Dalai, A. Production of activated carbon from Luscar char: Experimental and modeling studies. Microporous Mesoporous Mater. 2005, 85, 219–225. [Google Scholar] [CrossRef]
- Ghelich, R.; Jahannama, M.R.; Abdizadeh, H.; Torknik, F.S.; Vaezi, M.R. Central composite design (CCD)-Response surface methodology (RSM) of effective electrospinning parameters on PVP-B-Hf hybrid nanofibrous composites for synthesis of HfB2-based composite nanofibers. Compos. Part B Eng. 2019, 166, 527–541. [Google Scholar] [CrossRef]
- Ahmadi, M.; Jorfi, S.; Kujlu, R.; Ghafari, S.; Soltani, R.D.C.; Haghighifard, N.J. A novel salt-tolerant bacterial consortium for biodegradation of saline and recalcitrant petrochemical wastewater. J. Environ. Manag. 2017, 191, 198–208. [Google Scholar] [CrossRef]
- Phan, D.C.; Goodwin, D.G.; Frank, B.P.; Bouwer, E.J.; Fairbrother, D.H. Biodegradability of carbon nanotube/polymer nanocomposites under aerobic mixed culture conditions. Sci. Total Environ. 2018, 639, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Gangola, S.; Bhandari, G.; Zhang, W.; Maithani, D.; Mishra, S.; Chen, S. New insights into the degradation of synthetic pollutants in contaminated environments. Chemosphere 2021, 268, 128827. [Google Scholar] [CrossRef]
- Sarkar, P.; Dey, A. 4-Nitrophenol biodegradation by an isolated and characterized microbial consortium and statistical optimization of physicochemical parameters by Taguchi Methodology. J. Environ. Chem. Eng. 2020, 8, 104347. [Google Scholar] [CrossRef]
- Muñoz, R.; Alvarez, M.T.; Muñoz, A.; Terrazas, E.; Guieysse, B.; Mattiasson, B. Sequential removal of heavy metals ions and organic pollutants using an algal-bacterial consortium. Chemosphere 2006, 63, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, A.; Abbes, C.; Ben Mouhoub, R.; Ben Hassine, S.; Landoulsi, A. Enhancement of mixture pollutant biodegradation efficiency using a bacterial consortium under static magnetic field. PLoS ONE 2019, 14, e0208431. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Razek, M.A.; Abozeid, A.M.; Eltholth, M.M.; Abouelenien, F.A.; El-Midany, S.A.; Moustafa, N.Y.; Mohamed, R.A. others Bioremediation of a pesticide and selected heavy metals in wastewater from various sources using a consortium of microalgae and cyanobacteria. Slov. Vet. 2019, 56, 61–73. [Google Scholar]
- Allam, N. Bioremediation Efficiency of Heavy Metals and Azo Dyes by Individual or Consortium Bacterial Species Either as Free orImmobilized Cells: A Comparative Study. Egypt. J. Bot. 2017, 57, 555–564. [Google Scholar] [CrossRef]
- Pizarro-Tobias, P.; Niqui, J.L.; Roca, A.; Solano, J.; Fernández, M.; Bastida, F.; García, C.; Ramos, J.L. Field trial on removal of petroleum-hydrocarbon pollutants using a microbial consortium for bioremediation and rhizoremediation. Environ. Microbiol. Rep. 2014, 7, 85–94. [Google Scholar] [CrossRef]
- Nozari, M.; Samaei, M.R.; Dehghani, M.; Ebrahimi, A.A. Bioremediation of Alkane Hydrocarbons Using Bacterial Consortium From Soil. Health Scope 2018, 7, e12524. [Google Scholar] [CrossRef]
- Hameed, B.B.; Ismail, Z.Z. Decolorization, biodegradation and detoxification of reactive red azo dye using non-adapted immobilized mixed cells. Biochem. Eng. J. 2018, 137, 71–77. [Google Scholar] [CrossRef]
- Ismail, Z.Z.; Khudhair, H.A. Recycling of immobilized cells for aerobic biodegradation of phenol in a fluidized bed bioreactor. Syst. Cybern. Inform. 2015, 13, 81–86. [Google Scholar]
- Maniyam, M.N.; Yaacob, N.S.; Azman, H.H.; Ab Ghaffar, N.A.; Abdullah, H. Immobilized cells of Rhodococcus strain UCC 0004 as source of green biocatalyst for decolourization and biodegradation of methyl orange. Biocatal. Agric. Biotechnol. 2018, 16, 569–578. [Google Scholar] [CrossRef]
- Dogan, N.M.; Sensoy, T.; Doganli, G.A.; Bozbeyoglu, N.N.; Arar, D.; Akdogan, H.A.; Canpolat, M. Immobilization of Lycinibacillus fusiformis B26 cells in different matrices for use in turquoise blue HFG decolourization. Arch. Environ. Prot. 2016, 42, 92–99. [Google Scholar] [CrossRef] [Green Version]
- García, J.M.; Garcia-Martinez, T.; Mauricio, J.C.G.; Moreno, J. Yeast Immobilization Systems for Alcoholic Wine Fermentations: Actual Trends and Future Perspectives. Front. Microbiol. 2018, 9, 241. [Google Scholar] [CrossRef] [Green Version]
- Akpor, O.B. Research Article Dye Decolouration by Immobilized and Free Bacterial Cells at Different Glucose Concentration. 2018. Res. J. Environ. Sci. 2018, 12, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Susilowati, P.E.; A Rajiani, N.; Hermawan, H.; Zaeni, A.; Sudiana, I.N. The use Immobilized Bacteria-Alginate-Chitin for crack remediation. IOP Conf. Ser. Earth Environ. Sci. 2019, 299, 012010. [Google Scholar] [CrossRef]
- Jeon, Y.; Bissessur, A.; Singh, P. Novel immobilization techniques of Acinetobacter (V2) and Paenibacillus (D9) bacterial strains for waste oil degradation. Biotechnol. Biotechnol. Equip. 2019, 33, 911–920. [Google Scholar] [CrossRef]
- Staroń, P.; Chwastowski, J. Raphia-Microorganism Composite Biosorbent for Lead Ion Removal from Aqueous Solutions. Materials 2021, 14, 7482. [Google Scholar] [CrossRef] [PubMed]
- Vaiano, V.; Sacco, O.; Libralato, G.; Lofrano, G.; Siciliano, A.; Carraturo, F.; Guida, M.; Carotenuto, M. Degradation of anionic azo dyes in aqueous solution using a continuous flow photocatalytic packed-bed reactor: Influence of water matrix and toxicity evaluation. J. Environ. Chem. Eng. 2020, 8, 104549. [Google Scholar] [CrossRef]
- Mapukata, S.; Kobayashi, N.; Kimura, M.; Nyokong, T. Asymmetrical and symmetrical zinc phthalocyanine-cobalt ferrite conjugates embedded in electrospun fibers for dual photocatalytic degradation of azo dyes: Methyl Orange and Orange G. J. Photochem. Photobiol. A Chem. 2019, 379, 112–122. [Google Scholar] [CrossRef]
- Yang, H.-Y.; Liu, J.; Wang, Y.-X.; He, C.-S.; Zhang, L.-S.; Mu, Y.; Li, W.-H. Bioelectrochemical decolorization of a reactive diazo dye: Kinetics, optimization with a response surface methodology, and proposed degradation pathway. Bioelectrochemistry 2019, 128, 9–16. [Google Scholar] [CrossRef]
- Kolekar, Y.M.; Nemade, H.N.; Markad, V.L.; Adav, S.S.; Patole, M.S.; Kodam, K.M. Decolorization and biodegradation of azo dye, reactive blue 59 by aerobic granules. Bioresour. Technol. 2012, 104, 818–822. [Google Scholar] [CrossRef]
- Al-Baldawi, I.A.; Abdullah, S.R.S.; Almansoory, A.F.; Ismail, N.I.; Abu Hasan, H.; Anuar, N. Role of Salvinia molesta in biodecolorization of methyl orange dye from water. Sci. Rep. 2020, 10, 13980. [Google Scholar] [CrossRef]
- Croce, R.; Cinà, F.; Lombardo, A.; Crispeyn, G.; Cappelli, C.I.; Vian, M.; Maiorana, S.; Benfenati, E.; Baderna, D. Aquatic toxicity of several textile dye formulations: Acute and chronic assays with Daphnia magna and Raphidocelis subcapitata. Ecotoxicol. Environ. Saf. 2017, 144, 79–87. [Google Scholar] [CrossRef]
- Manzar, M.S.; Waheed, A.; Qazi, I.W.; Blaisi, N.I.; Ullah, N. Synthesis of a novel epibromohydrin modified crosslinked polyamine resin for highly efficient removal of methyl orange and eriochrome black T. J. Taiwan Inst. Chem. Eng. 2019, 97, 424–432. [Google Scholar] [CrossRef]
- Prabhakar, Y.; Gupta, A.; Kaushik, A. Enhanced decolorization of reactive violet dye 1 by halo-alkaliphilic Nesterenkonia strain: Process optimization, short acclimatization and reusability analysis in batch cycles. Process. Saf. Environ. Prot. 2019, 131, 116–126. [Google Scholar] [CrossRef]
- Mirbolooki, H.; Amirnezhad, R.; Pendashteh, A.R. Treatment of high saline textile wastewater by activated sludge microorganisms. J. Appl. Res. Technol. 2017, 15, 167–172. [Google Scholar] [CrossRef]
- Benkhaya, S.; M’Rabet, S.; El Harfi, A. Classifications, properties, recent synthesis and applications of azo dyes. Heliyon 2020, 6, e03271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masarbo, R.S. Decolorization of Sulfonated Azo Dye Methyl Orange by Bacterial Species. Int. J. Curr. Res. Acad. Rev. 2016, 4, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhang, J.; Zhang, C.; Yue, Q.; Li, Y.; Li, C. Equilibrium and kinetic studies of methyl orange and methyl violet adsorption on activated carbon derived from Phragmites australis. Desalination 2010, 252, 149–156. [Google Scholar] [CrossRef]
- Zabłocka-Godlewska, E.; Przystaś, W.; Grabińska-Sota, E. Dye Decolourisation Using Two Klebsiella Strains. Water Air Soil Pollut. 2015, 226, 2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, M.; Haque, A.; Mosharaf, K.; Marcus, P.K. Decolorization, degradation and detoxification of carcinogenic sulfonated azo dye methyl orange by newly developed biofilm consortia. Saudi J. Biol. Sci. 2021, 28, 793–804. [Google Scholar] [CrossRef]
- Rahman, M.M.; Khan, A.A.; Ali, M.E. others Screening of antagonistic bacteria against bacterial wilt of tomato, eggplant and potato in Bangladesh. Int. J. Agric. Biol. 2013, 15, 973–975. [Google Scholar]
- Eltarahony, M.; Zaki, S.; Kheiralla, Z.; Abd-El-Haleem, D. NAP enzyme recruitment in simultaneous bioremediation and nanoparticles synthesis. Biotechnol. Rep. 2018, 18, e00257. [Google Scholar] [CrossRef] [PubMed]
- Devi, P.; Wahidullah, S.; Sheikh, F.; Pereira, R.; Narkhede, N.; Amonkar, D.; Tilvi, S.; Meena, R.M. Biotransformation and Detoxification of Xylidine Orange Dye Using Immobilized Cells of Marine-Derived Lysinibacillus sphaericus D3. Mar. Drugs 2017, 15, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, M.C.; López, O.V.; Ciolino, A.E.; Morata, V.I.; Villar, M.A.; Ninago, M.D. Immobilization of enological pectinase in calcium alginate hydrogels: A potential biocatalyst for winemaking. Biocatal. Agric. Biotechnol. 2019, 18, 101091. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Xu, Y.; Xu, L.; Shi, T.; Ma, X.; Wu, X.; Li, Q.X.; Hua, R. Enhanced biodegradation of organophosphorus insecticides in industrial wastewater via immobilized Cupriavidus nantongensis X1T. Sci. Total Environ. 2020, 755, 142505. [Google Scholar] [CrossRef]
- Ruan, B.; Wu, P.; Chen, M.; Lai, X.; Chen, L.; Yu, L.; Gong, B.; Kang, C.; Dang, Z.; Shi, Z.; et al. Immobilization of Sphingomonas sp. GY2B in polyvinyl alcohol–alginate–kaolin beads for efficient degradation of phenol against unfavorable environmental factors. Ecotoxicol. Environ. Saf. 2018, 162, 103–111. [Google Scholar] [CrossRef]
- Abu-Elreesh, G.; El-Shall, H.; Eltarahony, M.; Abdelhaleem, D. Conversion of Cost-Effective agricultural wastes into valued oil using the fungus Curvularia Sp: Isolation, Optimization and Statistical analysis. Biosci. Res. 2019, 16, 3006–3024. [Google Scholar]
- Chen, K.-C.; Wu, J.-Y.; Liou, D.-J.; Hwang, S.-C.J. Decolorization of the textile dyes by newly isolated bacterial strains. J. Biotechnol. 2003, 101, 57–68. [Google Scholar] [CrossRef]
- Eltarahony, M.; Ibrahim, A.; El-Shall, H.; Ibrahim, E.; Althobaiti, F.; Fayad, E. Antibacterial, Antifungal and Antibiofilm Activities of Silver Nanoparticles Supported by Crude Bioactive Metabolites of Bionanofactories Isolated from Lake Mariout. Molecules 2021, 26, 3027. [Google Scholar] [CrossRef] [PubMed]
- Glenn, J.K.; Gold, M.H. Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-degrading basidiomycete, Phanerochaete chrysosporium. Arch. Biochem. Biophys. 1985, 242, 329–341. [Google Scholar] [CrossRef]
- Redinbaugh, M.G.; Campbell, W. Quaternary structure and composition of squash NADH:nitrate reductase. J. Biol. Chem. 1985, 260, 3380–3385. [Google Scholar] [CrossRef]
- Shanmugam, V.; Kumari, M.; Yadav, K.D. n-Propanol as a substrate for assaying the ligninperoxidase activity of Phanerochaete chrysoporium. Indian J. Biochem. Biophys. 1999, 36. [Google Scholar]
- Sambasiva Rao, K.R.S.; Tripathy, N.K.; Srinivasa Rao, D.; Prakasham, R.S. Production, characterization, catalytic and inhibitory activities of tyrosinase. Res. J. Biotechnol. 2013, 8, 1. [Google Scholar]
- Zhao, M.; Sun, P.-F.; Du, L.-N.; Wang, G.; Jia, X.-M.; Zhao, Y.-H. Biodegradation of methyl red by Bacillus sp. strain UN2: Decolorization capacity, metabolites characterization, and enzyme analysis. Environ. Sci. Pollut. Res. 2014, 21, 6136–6145. [Google Scholar] [CrossRef] [PubMed]
- El Fakharany, E.; Hassan, M.A.; Taha, T.H. Production and Application of Extracellular Laccase Produced by Fusarium oxysporum EMT. Int. J. Agric. Biol. 2016, 18, 939–947. [Google Scholar] [CrossRef]
- Rekik, H.; Jaouadi, N.Z.; Bouacem, K.; Zenati, B.; Kourdali, S.; Badis, A.; Annane, R.; Bouanane-Darenfed, A.; Bejar, S.; Jaouadi, B. Physical and enzymatic properties of a new manganese peroxidase from the white-rot fungus Trametes pubescens strain i8 for lignin biodegradation and textile-dyes biodecolorization. Int. J. Biol. Macromol. 2019, 125, 514–525. [Google Scholar] [CrossRef]
- Sari, I.P.; Simarani, K. Decolorization of selected azo dye by Lysinibacillus fusiformis W1B6: Biodegradation optimization, isotherm, and kinetic study biosorption mechanism. Adsorpt. Sci. Technol. 2019, 37, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, X.; Geng, A. Expression of a novel manganese peroxidase from Cerrena unicolor BBP6 in Pichia pastoris and its application in dye decolorization and PAH degradation. Biochem. Eng. J. 2020, 153, 107402. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zaki, S.A.; Eltarahony, M.; Abd-El-Haleem, D.A. Disinfection of water and wastewater by biosynthesized magnetite and zerovalent iron nanoparticles via NAP-NAR enzymes of Proteus mirabilis 10B. Environ. Sci. Pollut. Res. 2019, 26, 23661–23678. [Google Scholar] [CrossRef]
- Shebany, Y.M.; El-Dawy, E.G.; Gherbawy, Y.A. Potential of Different Fungi Species in Biodegradation Field of Phenolic Compounds. Annu. Res. Rev. Biol. 2019, 1–9. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Bharti, V.; Vikrant, K.; Goswami, M.; Tiwari, H.; Sonwani, R.K.; Lee, J.; Tsang, D.C.; Kim, K.-H.; Saeed, M.; Kumar, S.; et al. Biodegradation of methylene blue dye in a batch and continuous mode using biochar as packing media. Environ. Res. 2019, 171, 356–364. [Google Scholar] [CrossRef]
- Fuentes, M.S.; Briceño, G.E.; Saez, J.M.; Benimeli, C.; Diez, M.C.; Amoroso, M.J. Enhanced Removal of a Pesticides Mixture by Single Cultures and Consortia of Free and Immobilized Streptomyces Strains. BioMed Res. Int. 2013, 2013, 392573. [Google Scholar] [CrossRef] [Green Version]
- El-Naggar, N.E.-A.; Rabei, N.H.; El-Malkey, S.E. Eco-friendly approach for biosorption of Pb2+ and carcinogenic Congo red dye from binary solution onto sustainable Ulva lactuca biomass. Sci. Rep. 2020, 10, 16021. [Google Scholar] [CrossRef]
- Eltarahony, M.; Zaki, S.; Abd-El-Haleem, D. Aerobic and anaerobic removal of lead and mercury via calcium carbonate precipitation mediated by statistically optimized nitrate reductases. Sci. Rep. 2020, 10, 4029. [Google Scholar] [CrossRef] [Green Version]
- Akansha, K.; Chakraborty, D.; Sachan, S.G. Decolorization and degradation of methyl orange by Bacillus stratosphericus SCABiocatal. Agric. Biotechnol. 2019, 18, 101044. [Google Scholar] [CrossRef]
- Kalyani, D.C.; Telke, A.A.; Govindwar, S.P.; Jadhav, J.P. Biodegradation and Detoxification of Reactive Textile Dye by IsolatedPseudomonassp. SUK1. Water Environ. Res. 2009, 81, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Harikumar, P.S.; Joseph, L.; Dhanya, A. Photocatalytic degradation of textile dyes by hydrogel supported titanium dioxide nanoparticles. J. Environ. Eng. Ecol. Sci. 2013, 2. [Google Scholar]
- Masarbo, R.S.; Ismailsab, M.; Monisha, T.R.; Nayak, A.S.; Karegoudar, T.B. Enhanced decolorization of sulfonated azo dye methyl orange by single and mixed bacterial strains AK1, AK2 and VKY1. Bioremediat. J. 2018, 22, 136–146. [Google Scholar] [CrossRef]
- Baena-Baldiris, D.; Montes-Robledo, A.; Baldiris-Avila, R. Franconibacter sp., 1MS: A New Strain in Decolorization and Degradation of Azo Dyes Ponceau S Red and Methyl Orange. ACS Omega 2020, 5, 28146–28157. [Google Scholar] [CrossRef]
- Raghukumar, C.; Chandramohan, D.; Michel, F.C.; Redd, C.A. Degradation of lignin and decolorization of paper mill bleach plant effluent (BPE) by marine fungi. Biotechnol. Lett. 1996, 18, 105–106. [Google Scholar] [CrossRef]
- Sghaier, I.; Ouertani, R.; Mahjoubi, M.; El-Hidri, D.; Hassen, W.; Chamkhi, A.; Chouchane, H.; Jaouani, A.; Cherif, A.; Neifar, M. Application of a mixture design to optimize textile azo-dye decolorization using a bacterial consortium. Biom. Biostat. Int. J. 2019, 8, 58–63. [Google Scholar] [CrossRef]
- Tang, W.; Xu, X.; Ye, B.-C.; Cao, P.; Ali, A. Decolorization and degradation analysis of Disperse Red 3B by a consortium of the fungus Aspergillus sp. XJ-2 and the microalgae Chlorella sorokiniana XJK. RSC Adv. 2019, 9, 14558–14566. [Google Scholar] [CrossRef] [Green Version]
- Contreras, M.; Grande-Tovar, C.D.; Vallejo, W.; Chaves-López, C. Bio-Removal of Methylene Blue from Aqueous Solution by Galactomyces geotrichum KL20A. Water 2019, 11, 282. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.P.; Patel, K.A.; Darji, A.M. Microbial Degradation and Decolorization of Methyl Orange Dye by an Application of Pseudomonas Spp. ETL-1982. Int. J. Environ. Bioremediation Biodegrad. 2013, 1, 26–36. [Google Scholar] [CrossRef]
- Zommere, Ž.; Nikolajeva, V. others Immobilization of bacterial association in alginate beads for bioremediation of oil-contaminated lands. Environ. Exp. Biol. 2017, 15, 105–111. [Google Scholar]
- Giese, E.C. Mining Applications of Immobilized Microbial Cells in an Alginate Matrix: An Overview. Rev. Int. Contam. Ambient. 2020, 36, 775–787. [Google Scholar] [CrossRef]
- Das, A.; Mishra, S. Removal of textile dye reactive green-19 using bacterial consortium: Process optimization using response surface methodology and kinetics study. J. Environ. Chem. Eng. 2017, 5, 612–627. [Google Scholar] [CrossRef]
- Chenthamara, D.; Subramaniam, S.; Ramakrishnan, S.G.; Krishnaswamy, S.; Essa, M.M.; Lin, F.H.; Qoronfleh, M.W. Therapeutic efficacy of nanoparticles and routes of administration. Biomater. Res. 2019, 23, 20. [Google Scholar] [CrossRef]
- Cui, D.; Zhang, H.; He, R.; Hao, Z. The Comparative Study on the Rapid Decolorization of Azo, Anthraquinone and Triphenylmethane Dyes by Anaerobic Sludge. Int. J. Environ. Res. Public Health 2016, 13, 1053. [Google Scholar] [CrossRef]
- Ayed, L.; Chaieb, K.; Cheref, A.; Bakhrouf, A. Biodegradation of triphenylmethane dye Malachite Green by Sphingomonas paucimobilis. World J. Microbiol. Biotechnol. 2009, 25, 705–711. [Google Scholar] [CrossRef]
- Sheela, T.; Sadasivam, S.K. Dye degradation potential and its degradative enzymes synthesis of Bacillus cereus SKB12 isolated from a textile industrial effluent. J. Appl. Biol. Biotechnol. 2020, 8, 42–46. [Google Scholar]
- Thanavel, M.; Kadam, S.K.; Biradar, S.P.; Govindwar, S.P.; Jeon, B.-H.; Sadasivam, S.K. Combined biological and advanced oxidation process for decolorization of textile dyes. SN Appl. Sci. 2018, 1, 97. [Google Scholar] [CrossRef] [Green Version]
- Bedekar, P.A.; Kshirsagar, S.D.; Gholave, A.R.; Govindwar, S.P. Degradation and detoxification of methylene blue dye adsorbed on water hyacinth in semi continuous anaerobic–aerobic bioreactors by novel microbial consortium-SB. RSC Adv. 2015, 5, 99228–99239. [Google Scholar] [CrossRef]
- Oukarroum, A.; Zaidi, W.; Samadani, M.; Dewez, D. Toxicity of Nickel Oxide Nanoparticles on a Freshwater Green Algal Strain ofChlorella vulgaris. BioMed Res. Int. 2017, 2017, 9528180. [Google Scholar] [CrossRef] [Green Version]
- Demmig-Adams, B.; Adams, W.W. Antioxidants in Photosynthesis and Human Nutrition. Science 2002, 298, 2149–2153. [Google Scholar] [CrossRef] [PubMed]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.-F. Microalgal Carotenoids: A Review of Production, Current Markets, Regulations, and Future Direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.-S.; Hung, C.-H.; Chao, S.-L. Decolorization of azo dye reactive black B by Bacillus cereus strain HJ-1. Chemosphere 2013, 90, 2109–2114. [Google Scholar] [CrossRef]
- Qu, Y.; Zhou, J.; Wang, J.; Song, Z.; Xing, L.; Fu, X. Bioaugmentation of Bromoamine Acid Degradation with Sphingomonas xenophaga QYY and DNA Fingerprint Analysis of Augmented Systems. Biodegradation 2006, 17, 83–91. [Google Scholar] [CrossRef]
- Poi, G.; Shahsavari, E.; Aburto-Medina, A. Large Scale Bioaugmentation of Municipal Waste Water Contaminated with Petroleum Hydrocarbons. JSM Environ. Sci. Ecol. 2016, 4, 1036. [Google Scholar]
- Jasmine, J.; Mukherji, S. Evaluation of bioaugmentation and biostimulation effects on the treatment of refinery oily sludge using 2n full factorial design. Environ. Sci. Process. Impacts 2014, 16, 1889–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Coded Levels/Experimental Values | ||||
|---|---|---|---|---|---|
| −2 | −1 | 0 | 1 | 2 | |
| Incubation time (days) | 1 | 2 | 3 | 4 | 5 |
| Inoculum Size (%) | 1 | 3 | 5 | 10 | 20 |
| MO Conc. (mg/L) | 200 | 250 | 300 | 350 | 400 |
| NaCl Conc. (g/L) | 5 | 10 | 15 | 20 | 40 |
| Run Order | Incubation Time (Day) | Inoculum Size | MO Conc. | NaCl Conc. | Experimental MO Degradation (%) | Predicted MO Degradation (%) | St. Residual |
|---|---|---|---|---|---|---|---|
| 1 | 0 | 0 | 2 | 0 | 2.25 | 5.825 | −2.17 |
| 2 | 0 | 0 | −2 | 0 | 46.38 | 43.61 | 1.68 |
| 3 | −1 | 1 | −1 | −1 | 25.14 | 22.029 | 1.89 |
| 4 | −1 | −1 | −1 | 1 | 11.37 | 11.834 | −0.28 |
| 5 | 0 | 0 | 0 | 0 | 8.31 | 8.641 | −0.14 |
| 6 | −2 | 0 | 0 | 0 | 0.27 | 2.122 | −1.13 |
| 7 | −1 | −1 | 1 | 1 | 0.52 | −0.638 | 0.7 |
| 8 | 1 | −1 | −1 | 1 | 22.4 | 25.081 | −1.63 |
| 9 | 0 | 0 | 0 | 0 | 11.5 | 8.641 | 1.21 |
| 10 | 0 | 0 | 0 | −2 | 1.1 | 3.7 | −1.58 |
| 11 | 0 | −2 | 0 | 0 | 2.81 | 1.7 | 0.67 |
| 12 | 1 | 1 | 1 | 1 | 5.24 | 5.171 | 0.04 |
| 13 | 0 | 2 | 0 | 0 | 8.58 | 10.495 | −1.16 |
| 14 | 1 | −1 | 1 | −1 | 3.49 | 2.771 | 0.44 |
| 15 | 0 | 0 | 0 | 0 | 9.11 | 8.641 | 0.2 |
| 16 | −1 | 1 | 1 | −1 | 2.36 | 1.067 | 0.79 |
| 17 | 1 | 1 | −1 | 1 | 32.77 | 32.082 | 0.42 |
| 18 | 0 | 0 | 0 | 2 | 11.08 | 9.285 | 1.09 |
| 19 | 1 | −1 | 1 | 1 | 7.34 | 8.257 | −0.56 |
| 20 | 2 | 0 | 0 | 0 | 16.62 | 15.574 | 0.64 |
| 21 | 0 | 0 | 0 | 0 | 8.31 | 8.641 | −0.14 |
| 22 | 1 | 1 | 1 | −1 | 3.93 | 1.272 | 1.62 |
| 23 | −1 | 1 | −1 | 1 | 20.02 | 22.128 | −1.28 |
| 24 | −1 | −1 | 1 | −1 | 0.78 | −0.726 | 0.92 |
| 25 | 1 | 1 | −1 | −1 | 24.04 | 26.586 | −1.55 |
| 26 | 0 | 0 | 0 | 0 | 7.75 | 8.641 | −0.38 |
| 27 | 0 | 0 | 0 | 0 | 6 | 8.641 | −1.12 |
| 28 | −1 | −1 | −1 | −1 | 8.69 | 10.148 | −0.89 |
| 29 | −1 | 1 | 1 | 1 | 1.04 | −0.431 | 0.89 |
| 30 | 1 | −1 | −1 | −1 | 18.72 | 17.997 | 0.44 |
| 31 | 0 | 0 | 0 | 0 | 9.51 | 8.641 | 0.37 |
| Term | Coef. | SE Coef. | T | p Value |
|---|---|---|---|---|
| Constant | 8.64143 | 0.9636 | 8.968 | 0 |
| Incubation days | 3.36292 | 0.5204 | 6.462 | 0 |
| Inoculum size | 2.19875 | 0.5204 | 4.225 | 0.001 |
| MO conc. | −9.44625 | 0.5204 | −18.152 | 0 |
| NaCl conc. | 1.39625 | 0.5204 | 2.683 | 0.016 |
| (Incubation days)2 | 0.05162 | 0.4768 | 0.108 | 0.915 |
| (Inoculum size)2 | −0.63588 | 0.4768 | −1.334 | 0.201 |
| (MO conc.)2 | 4.01912 | 0.4768 | 8.43 | 0 |
| (NaCl conc.)2 | −0.53713 | 0.4768 | −1.127 | 0.277 |
| Incubation days × Inoculum size | −0.82313 | 0.6374 | −1.291 | 0.215 |
| Incubation days × MO conc. | −1.08812 | 0.6374 | −1.707 | 0.107 |
| Incubation days × NaCl conc. | 1.34938 | 0.6374 | 2.117 | 0.05 |
| Inoculum size × MO conc. | −2.52188 | 0.6374 | −3.957 | 0.001 |
| Inoculum size × NaCl conc. | −0.39687 | 0.6374 | −0.623 | 0.542 |
| MO conc. × NaCl conc. | −0.39937 | 0.6374 | −0.627 | 0.54 |
| Source | Df | Seq. SS | Adj. SS | Adj. MS | F | p Value |
|---|---|---|---|---|---|---|
| Regression | 14 | 3259.04 | 3259.04 | 232.788 | 35.82 | 0 |
| Linear | 4 | 2575.8 | 2575.8 | 643.949 | 99.07 | 0 |
| Square | 4 | 517.49 | 517.49 | 129.373 | 19.9 | 0 |
| Interaction | 6 | 165.75 | 165.75 | 27.625 | 4.25 | 0.01 |
| Residual error | 16 | 103.99 | 103.99 | 6.5 | ||
| Lack of fit | 10 | 86.86 | 86.86 | 8.686 | 3.04 | 0.093 |
| Pure error | 6 | 17.14 | 17.14 | 2.856 | ||
| Total | 30 | 3363.03 |
| Parameter | Industrial Wastewater Concentration | Agricultural Wastewater Concentration |
|---|---|---|
| Total nitrogen (mg/L) | 113.44 | 411.2 |
| Total phosphates (mg/L) | 0.2 | 194.6 |
| T.D. (mg/L) | 1060 | 927 |
| Nitrate (mg/L) | 18.77 | 274.9 |
| Nitrite (mg/L) | 2.5 | 44.9 |
| Ammonia (mg/L) | 0.135 | 160.6 |
| Carbonate (mg/L) | 81.6 | 84 |
| Sulfate (mg/L) | 133.7 | 424.58 |
| Sulfide (mg/L) | 198.6 | 58.9 |
| Phenol (mg/L) | 73.5 | 0.04 |
| Oil (mg/L) | 185.2 | 13.3 |
| B.O.D (mg/L) | 120.8 | 183 |
| C.O.D (mg/L) | 310 | 204 |
| Turbidity (NTU) | 464.3 | 12.5 |
| E.C (μs) | 1003 | 901 |
| Total count (CFU/L) | 8.34 × 103 | 6.74 × 105 |
| Calcium (mg/L) | 152.52 | 440.68 |
| Zn (mg/L) | 13.4 | 1.2 |
| Fe (mg/L) | 28.6 | 9.8 |
| Cr (mg/L) | 6.6 | <0.01 |
| Cd (mg/L) | 3.5 | <0.01 |
| Cu (mg/L) | 11.2 | 2.3 |
| Co (mg/L) | 7.4 | 0.1 |
| Ag (mg/L) | 5.3 | <0.01 |
| Pb (mg/L) | 9.6 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, A.; El-Fakharany, E.M.; Abu-Serie, M.M.; ElKady, M.F.; Eltarahony, M. Methyl Orange Biodegradation by Immobilized Consortium Microspheres: Experimental Design Approach, Toxicity Study and Bioaugmentation Potential. Biology 2022, 11, 76. https://doi.org/10.3390/biology11010076
Ibrahim A, El-Fakharany EM, Abu-Serie MM, ElKady MF, Eltarahony M. Methyl Orange Biodegradation by Immobilized Consortium Microspheres: Experimental Design Approach, Toxicity Study and Bioaugmentation Potential. Biology. 2022; 11(1):76. https://doi.org/10.3390/biology11010076
Chicago/Turabian StyleIbrahim, Amany, Esmail M. El-Fakharany, Marwa M. Abu-Serie, Marwa F. ElKady, and Marwa Eltarahony. 2022. "Methyl Orange Biodegradation by Immobilized Consortium Microspheres: Experimental Design Approach, Toxicity Study and Bioaugmentation Potential" Biology 11, no. 1: 76. https://doi.org/10.3390/biology11010076