
Heterotypic Multicellular Spheroids as Experimental and Preclinical Models of Sprouting Angiogenesis

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Sprouting Angiogenesis
3. Endothelial and Perivascular Cell Types
4. 3D Tissue Spheroids Comprised of ECs and Perivascular Cells
5. The Functionality of Prevascularized 3D Tissue Spheroids
6. 3D Spheroid Sprouting Model
7. Role of ECM in 3D Spheroid Sprouting Model
8. The Application of 3D Heterotypic Endothelial-Perivascular Spheroid System
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Folkman, J.; Cole, P.; Zimmerman, S. Tumor behavior in isolated perfused organs: In vitro growth and metastases of biopsy material in rabbit thyroid and canine intestinal segment. Ann. Surg. 1966, 164, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Gimbrone, M.A.; Leapman, S.B.; Cotran, R.S.; Folkman, J. Tumor angiogenesis: Iris neovascularization at a distance from experimental intraocular tumors. J. Natl. Cancer Inst. 1973, 50, 219–228. [Google Scholar] [CrossRef]
- Nicosia, R.F.; Tchao, R.; Leighton, J. Angiogenesis-dependent Tumor Spread in Reinforced Fibrin Clot Culture. Cancer Res. 1983, 43, 2159–2166. [Google Scholar]
- Nicosia, R.F.; Ottinetti, A. Modulation of Microvascular Growth and Morphogenesis by Reconstituted Basement Membrane Gel in Three-Dimensional Cultures of Rat Aorta: A Comparative Study of Angiogenesis in Matrigel, Collagen, Fibrin, and Plasma Clot. Vitr. Cell. Dev. Biol.-Anim. 1990, 26, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Nicosia, R.F. The aortic ring model of angiogenesis: A quarter century of search and discovery. J. Cell. Mol. Med. 2009, 13, 4113–4136. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J. Clin. Investig. 1973, 52, 2745–2756. [Google Scholar] [CrossRef] [PubMed]
- Gimbrone, M.A.; Cotran, R.S.; Folkman, J. Endothelial regeneration: Studies with human endothelial cells in culture. Ser. Haematol. 1973, 6, 453–455. [Google Scholar]
- Stryker, Z.I.; Rajabi, M.; Davis, P.J.; Mousa, S.A. Evaluation of Angiogenesis Assays. Biomedicines 2019, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Patel-Hett, S.; D’Amore, P.A. Signal Transduction in Vasculogenesis and Developmental Angiogenesis. Int. J. Dev. Biol. 2011, 55, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Bayless, K.J.; Davis, G.E. Sphingosine-1-phosphate markedly induces matrix metalloproteinase and integrin-dependent human endothelial cell invasion and lumen formation in three-dimensional collagen and fibrin matrices. Biochem. Biophys. Res. Commun. 2003, 312, 903–913. [Google Scholar] [CrossRef]
- Bayless, K.J.; Kwak, H.-I.; Su, S.C. Investigating endothelial invasion and sprouting behavior in three-dimensional collagen matrices. Nat. Protoc. 2009, 4, 1888–1898. [Google Scholar] [CrossRef]
- Davis, G.E.; Bayless, K.J.; Mavila, A. Molecular basis of endothelial cell morphogenesis in three-dimensional extracellular matrices. Anat. Rec. 2002, 268, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, M.N.; Davis, J.; Hughes, C.C.W. Optimized Fibrin Gel Bead Assay for the Study of Angiogenesis. J. Vis. Exp. 2007, 3, 186. [Google Scholar] [CrossRef] [Green Version]
- Korff, T.T. Tensional forces in fibrillar extracellular matrices control directional capillary sprouting. J. Cell Sci. 1999, 112, 3249–3258. [Google Scholar] [CrossRef]
- Korff, T.; Kimmina, S.; Martiny-Baron, G.; Augustin, H.G. Blood vessel maturation in a 3-dimensional spheroidal coculture model: Direct contact with smooth muscle cells regulates endothelial cell quiescence and abrogates VEGF responsiveness. FASEB J. 2001, 15, 447–457. [Google Scholar] [CrossRef]
- Gianni-Barrera, R.; Trani, M.; Reginato, S.; Banfi, A. To sprout or to split? VEGF, Notch and vascular morphogenesis. Biochem. Soc. Trans. 2011, 39, 1644–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribatti, D.; Crivellato, E. “Sprouting angiogenesis”, a reappraisal. Dev. Biol. 2012, 372, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amico, G.; Muñoz-Félix, J.M.; Pedrosa, A.R.; Hodivala-Dilke, K.M. “Splitting the matrix”: Intussusceptive angiogenesis meets MT1-MMP. EMBO Mol. Med. 2020, 12, e11663. [Google Scholar] [CrossRef]
- Sweeney, M.; Foldes, G. It Takes Two: Endothelial-Perivascular Cell Cross-Talk in Vascular Development and Disease. Front. Cardiovasc. Med. 2018, 5, 154. [Google Scholar] [CrossRef]
- Betz, C.; Lenard, A.; Belting, H.G.; Affolter, M. Cell behaviors and dynamics during angiogenesis. Development 2016, 143, 2249–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiaverina, G.; di Blasio, L.; Monica, V.; Accardo, M.; Palmiero, M.; Peracino, B.; Vara-Messler, M.; Puliafito, A.; Primo, L. Dynamic Interplay between Pericytes and Endothelial Cells during Sprouting Angiogenesis. Cells 2019, 8, 1109. [Google Scholar] [CrossRef] [Green Version]
- Kamei, M.; Brian Saunders, W.; Bayless, K.J.; Dye, L.; Davis, G.E.; Weinstein, B.M. Endothelial tubes assemble from intracellular vacuoles in vivo. Nature 2006, 442, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.E.; Stratman, A.N.; Sacharidou, A.; Koh, W. Molecular Basis for Endothelial Lumen Formation and Tubulogenesis During Vasculogenesis and Angiogenic Sprouting. Int. Rev. Cell Mol. Biol. 2011, 288, 101–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billroth, T. Untersuchungen uber die Entwicklung der Blutgefasse nebst Beobachtungen aus der Koniglichen Chirurgishen Universitats-Klinik zu Berlin; Reimer: Berlin, Germany, 1856; pp. 1–87. [Google Scholar]
- Sabin, F. Studies on the origin of blood vessels and of red corpuscles as seen in the living blastoderm of the chick during the second day of incubation. Contrib. Embryol. 1920, 9, 213–262. [Google Scholar]
- Xu, K.; Cleaver, O. Tubulogenesis during blood vessel formation. Semin. Cell Dev. Biol. 2011, 22, 993–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebala, V.; Collins, R.; Geudens, I.; Phng, L.K.; Gerhardt, H. Blood flow drives lumen formation by inverse membrane blebbing during angiogenesis in vivo. Nat. Cell Biol. 2016, 18, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P.; De Smet, F.; Loges, S.; Mazzone, M. Branching morphogenesis and antiangiogenesis candidates: Tip cells lead the way. Nat. Rev. Clin. Oncol. 2009, 6, 315–326. [Google Scholar] [CrossRef]
- Smith, A.O.; Bowers, S.L.K.; Stratman, A.N.; Davis, G.E. Hematopoietic Stem Cell Cytokines and Fibroblast Growth factor-2 Stimulate Human Endothelial Cell-Pericyte Tube Co-Assembly in 3D Fibrin Matrices under Serum-Free Defined Conditions. PLoS ONE 2013, 8, e85147. [Google Scholar] [CrossRef]
- Gerhardt, H. VEGF and endothelial guidance in angiogenic sprouting. Organogenesis 2008, 4, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Sliwinska, P.N.; Alitalo, K.; Allen, E.; Anisimov, A.; Aplin, A.C.; Auerbach, R.; Augustin, H.G.; Bates, D.O.; Van Beijnum, J.R.; Bender, R.H.F. Consensus guidelines for the use and interpretation of angiogenesis assays. Angiogenesis 2018, 21, 425–532. [Google Scholar] [CrossRef] [Green Version]
- Medina-Leyte, D.J.; Domínguez-Pérez, M.; Mercado, I.; Villarreal-Molina, M.T.; Jacobo-Albavera, L. Use of Human Umbilical Vein Endothelial Cells (HUVEC) as a Model to Study Cardiovascular Disease: A Review. Appl. Sci. 2020, 10, 938. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.; Kang, K.T. Two-cell spheroid angiogenesis assay system using both endothelial colony forming cells and mesenchymal stem cells. Biomol. Ther. 2018, 26, 474–480. [Google Scholar] [CrossRef]
- Shah, S.; Lee, H.; Park, Y.H.; Jeon, E.; Chung, H.K.; Lee, E.S.; Shim, J.H.; Kang, K.T. Three-dimensional angiogenesis assay system using co-culture spheroids formed by endothelial colony forming cells and mesenchymal stem cells. J. Vis. Exp. 2019, 151, e60032. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.W.; Tian, L.; Krick, S.; Helton, E.S.; Denson, R.S.; Comhair, S.A.A.; Dweik, R.A. O-GlcNAc transferase regulates angiogenesis in idiopathic pulmonary arterial hypertension. Int. J. Mol. Sci. 2019, 20, 6299. [Google Scholar] [CrossRef] [Green Version]
- Vorwald, C.E.; Joshee, S.; Leach, J.K. Spatial localization of endothelial cells in heterotypic spheroids influences NOTCH signaling. J. Mol. Med. 2020, 93, 425–435. [Google Scholar] [CrossRef]
- Fortunato, T.M.; Beltrami, C.; Emanueli, C.; De Bank, P.A.; Pula, G. Platelet lysate gel and endothelial progenitors stimulate microvascular network formation in vitro: Tissue engineering implications. Sci. Rep. 2016, 6, 25326. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Weisdorf, D.J.; Solovey, A.; Hebbel, R.P. Origins of circulating endothelial cells and endothelial outgrowth from blood. J. Clin. Investig. 2000, 105, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murohara, T. Cord blood-derived early outgrowth endothelial progenitor cells. Microvasc. Res. 2010, 79, 174–177. [Google Scholar] [CrossRef]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–967. [Google Scholar] [CrossRef]
- Asahara, T.; Masuda, H.; Takahashi, T.; Kalka, C.; Pastore, C.; Silver, M.; Kearne, M.; Magner, M.; Isner, J.M. Bone marrow origin of endothelial progenitor cells responsible for postnatal vasculogenesis in physiological and pathological neovascularization. Circ. Res. 1999, 85, 221–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, D.A.; Mead, L.E.; Tanaka, H.; Meade, V.; Fenoglio, A.; Mortell, K.; Ferkowicz, M.J.; Gilley, D.; Yoder, M.C.; Dc, W.; et al. Identification of a novel hierarchy of endothelial progenitor cells using human peripheral and umbilical cord blood. Blood 2004, 104, 2752–2760. [Google Scholar] [CrossRef] [PubMed]
- Prasain, N.; Meador, J.L.; Yoder, M.C. Phenotypic and Functional Characterization of Endothelial Colony Forming Cells Derived from Human Umbilical Cord Blood. J. Vis. Exp. 2012, 62, e3872. [Google Scholar] [CrossRef] [Green Version]
- Jaminon, A.; Reesink, K.; Kroon, A.; Schurgers, L. The Role of Vascular Smooth Muscle Cells in Arterial Remodeling: Focus on Calcification-Related Processes. Int. J. Mol. Sci. 2019, 20, 5694. [Google Scholar] [CrossRef] [Green Version]
- Birbrair, R.; Zhang, T.; Wang, Z.M.; Messi, M.L.; Mintz, A.; Delbono, O. Pericytes at the intersection between tissue regeneration and pathology. Clin. Sci. 2015, 128, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.G.; Zhu, S.Y.; Heng, B.C.; Dissanayaka, W.L.; Zhang, C.F. TGF-β1-induced differentiation of SHED into functional smooth muscle cells. Stem Cell Res. Ther. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, J.; Zou, T.; Qi, Y.; Yi, B.; Dissanayaka, W.L.; Zhang, C. DPSCs treated by TGF-β1 regulate angiogenic sprouting of three-dimensionally co-cultured HUVECs and DPSCs through VEGF-Ang-Tie2 signaling. Stem Cell Res. Ther. 2021, 12, 281. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Prolif. 1970, 3, 393–403. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef]
- Hass, R. Role of MSC in the Tumor Microenvironment. Cancers 2020, 12, 2107. [Google Scholar] [CrossRef]
- Ghajar, C.M.; Kachgal, S.; Kniazeva, E.; Mori, H.; Costes, S.V.; George, S.C.; Putnam, A.J. Mesenchymal cells stimulate capillary morphogenesis via distinct proteolytic mechanisms. Exp. Cell Res. 2010, 316, 813–825. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.Y.; Salazar, M.G.; Miller, S.; Meyers, C.; Ding, C.; Hardy, W.; Péault, B.; James, A.W. Comparison of human tissue microarray to human pericyte transcriptome yields novel perivascular cell markers. Stem Cells Dev. 2019, 28, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Nwadozi, E.; Rudnicki, M.; Haas, T.L. Metabolic Coordination of Pericyte Phenotypes: Therapeutic Implications. Front. Cell Dev. Biol. 2020, 8, 77. [Google Scholar] [CrossRef]
- Lin, R.Z.; Chang, H.Y. Recent advances in three-dimensional multicellular spheroid culture for biomedical research. Biotechnol. J. 2008, 3, 1172–1184. [Google Scholar] [CrossRef]
- Shpichka, A.I.; Konarev, P.V.; Efremov, Y.M.; Kryukova, A.E.; Aksenova, N.A.; Kotova, S.L.; Frolova, A.A.; Kosheleva, N.V.; Zhigalina, O.M.; Yusupov, V.I.; et al. Digging deeper: Structural background of PEGylated fibrin gels in cell migration and lumenogenesis. RSC Adv. 2020, 10, 4190–4200. [Google Scholar] [CrossRef]
- Carson, A.E.; Barker, T.H. Emerging concepts in engineering extracellular matrix variants for directing cell phenotype. Regen. Med. 2009, 4, 593. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.Y.; Kofron, C.M.; King, M.E.; Markes, A.R.; Okundaye, A.O.; Qu, Z.; Mende, U.; Choi, B.-R. Directed fusion of cardiac spheroids into larger heterocellular microtissues enables investigation of cardiac action potential propagation via cardiac fibroblasts. PLoS ONE 2018, 13, e0196714. [Google Scholar] [CrossRef] [Green Version]
- Kumarasamy, M.; Sosnik, A. Heterocellular spheroids of the neurovascular blood-brain barrier as a platform for personalized nanoneuromedicine. iScience 2021, 24, 102183. [Google Scholar] [CrossRef]
- Franchi-Mendes, T.; Lopes, N.; Brito, C. Heterotypic Tumor Spheroids in Agitation-Based Cultures: A Scaffold-Free Cell Model That Sustains Long-Term Survival of Endothelial Cells. Front. Bioeng. Biotechnol. 2021, 9, 649949. [Google Scholar] [CrossRef] [PubMed]
- Ryu, N.E.; Lee, S.H.; Park, H. Spheroid Culture System Methods and Applications for Mesenchymal Stem Cells. Cells 2019, 8, 1620. [Google Scholar] [CrossRef] [Green Version]
- Białkowska, K.; Komorowski, P.; Bryszewska, M.; Miłowska, K. Spheroids as a Type of Three-Dimensional Cell Cultures—Examples of Methods of Preparation and the Most Important Application. Int. J. Mol. Sci. 2020, 21, 6225. [Google Scholar] [CrossRef]
- Korff, T.; Augustin, H.G. Integration of endothelial cells in multicellular spheroids prevents apoptosis and induces differentiation. J. Cell Biol. 1998, 143, 1341–1352. [Google Scholar] [CrossRef]
- Heiss, M.; Hellström, M.; Kalén, M.; May, T.; Weber, H.; Hecker, M.; Augustin, H.G.; Korff, T. Endothelial cell spheroids as a versatile tool to study angiogenesis in vitro. FASEB J. 2015, 29, 3076–3084. [Google Scholar] [CrossRef] [PubMed]
- De Moor, L.; Merovci, I.; Baetens, S.; Verstraeten, J.; Kowalska, P.; Krysko, D.V.; De Vos, W.H.; Declercq, H. High-throughput fabrication of vascularized spheroids for bioprinting. Biofabrication 2018, 10, 35009. [Google Scholar] [CrossRef]
- Eckermann, C.W.; Lehle, K.; Schmid, S.A.; Wheatley, D.N.; Kunz-Schughart, L.A. Characterization and modulation of fibroblast/endothelial cell co-cultures for the in vitro preformation of three-dimensional tubular networks. Cell Biol. Int. 2011, 35, 1097–1110. [Google Scholar] [CrossRef]
- Kelm, J.M.; Lorber, V.; Snedeker, J.G.; Schmidt, D.; Broggini-Tenzer, A.; Weisstanner, M.; Odermatt, B.; Mol, A.; Zünd, G.; Hoerstrup, S.P. A novel concept for scaffold-free vessel tissue engineering: Self-assembly of microtissue building blocks. J. Biotechnol. 2010, 148, 46–55. [Google Scholar] [CrossRef]
- Rivron, N.C.; Raiss, C.C.; Liu, J.; Nandakumar, A.; Sticht, C.; Gretz, N. Sonic Hedgehog-activated engineered blood vessels enhance bone tissue formation. Proc. Natl. Acad. Sci. USA 2012, 109, 4413–4418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakhrushev, I.V.; (Laboratory of Cell Biology, Institute of Biomedical Chemistry, Moscow 119121, Russia); Tsvetkova, A.V.; (Laboratory of Cell Biology, Institute of Biomedical Chemistry, Moscow 119121, Russia); Nezhurina, E.K.; (P.A. Hertsen Moscow Oncology Research Center, National Medical Research Radiological Center, Moscow 125284, Russia); Karalkin, P.A.; (Institute for Cluster Oncology, Sechenov University, Moscow 119435, Russia). Unpublished work, 2021.
- Steinberg, M.S. Reconstruction of tissues by dissociated cells. Science 1963, 141, 401–408. [Google Scholar] [CrossRef]
- Foty, R.A.; Steinberg, M.S. The differential adhesion hypothesis: A direct evaluation. Dev. Biol. 2005, 278, 255–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tse, J.D.; Moore, R.; Meng, Y.; Tao, W.; Smith, E.R.; Xu, X.X. Dynamic conversion of cell sorting patterns in aggregates of embryonic stem cells with differential adhesive affinity. BMC Dev. Biol. 2021, 21, 2. [Google Scholar] [CrossRef]
- Marshall, J.; Barnes, A.; Genever, P. Analysis of the intrinsic self-organising properties of mesenchymal stromal cells in three-dimensional co-culture models with endothelial cells. Bioengineering 2018, 5, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, S.T.; Douglas, A.M.; Chadid, T.; Kuo, K.; Rajabalan, A.; Li, H.; Copland, I.B.; Barker, T.H.; Galipeau, J.; Brewster, L.P. A novel platelet lysate hydrogel for endothelial cell and mesenchymal stem cell-directed neovascularization. Acta Biomater. 2016, 36, 86–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.G.; Andrejecsk, J.W.; Kluger, M.S.; Saltzman, W.M.; Pober, J.S. Pericytes modulate endothelial sprouting. Cardiovasc. Res. 2013, 100, 492–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, D.N.; Hospodiuk, M.; Ozbolat, I.T. Synergistic interplay between human MSCs and HUVECs in 3D spheroids laden in collagen/fibrin hydrogels for bone tissue engineering. Acta Biomater. 2019, 95, 348–356. [Google Scholar] [CrossRef]
- Saleh, F.A.; Whyte, M.; Genever, P.G. Effects of endothelial cells on human mesenchymal stem cell activity in a three-dimensional in vitro model. Eur. Cells Mater. 2011, 22, 242–257. [Google Scholar] [CrossRef] [PubMed]
- Straface, G.; Aprahamian, T.; Flex, A.; Gaetani, E.; Biscetti, F.; Smith, R.C.; Pecorini, G.; Pola, E.; Angelini, F.; Stigliano, E.; et al. Sonic hedgehog regulates angiogenesis and myogenesis during post-natal skeletal muscle regeneration. J. Cell. Mol. Med. 2009, 13, 2424–2435. [Google Scholar] [CrossRef]
- Le, H.; Kleinerman, R.; Lerman, O.Z.; Brown, D.; Galiano, R.; Gurtner, G.C.; Warren, S.M.; Levine, J.P.; Saadeh, P.B. Hedgehog signaling is essential for normal wound healing. Wound Repair Regen. 2008, 16, 768–773. [Google Scholar] [CrossRef]
- Wenger, A.; Kowalewski, N.; Stahl, A.; Mehlhorn, A.T.; Schmal, H.; Stark, G.B.; Finkenzeller, G. Development and characterization of a spheroidal coculture model of endothelial cells and fibroblasts for improving angiogenesis in tissue engineering. Cells Tissues Organs 2005, 181, 80–88. [Google Scholar] [CrossRef]
- Bauman, E.; Feijão, T.; Carvalho, D.T.O.; Granja, P.L.; Barrias, C.C. Xeno-free pre-vascularized spheroids for therapeutic applications. Sci. Rep. 2018, 8, 230. [Google Scholar] [CrossRef]
- Wenger, A.; Stahl, A.; Weber, H.; Finkenzeller, G.; Augustin, H.G. Modulation of in vitro angiogenesis in a three-dimensional spheroidal coculture model for bone tissue engineering. Tissue Eng. 2004, 10, 1536–1547. [Google Scholar] [CrossRef]
- Duran, C.L.; Howell, D.W.; Dave, J.M.; Smith, R.L.; Torrie, M.E. Molecular Regulation of Sprouting Angiogenesis. Compr. Physiol. 2018, 8, 153–235. [Google Scholar] [CrossRef]
- Wirz, W.; Antoine, M.; Tag, C.G.; Gressner, A.M.; Korff, T.; Hellerbrand, C.; Kiefer, P. Hepatic stellate cells display a functional vascular smooth muscle cell phenotype in a three-dimensional co-culture model with endothelial cells. Differentiation 2008, 76, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Dissanayaka, W.L.; Zhu, L.; Hargreaves, K.M.; Jin, L.; Zhang, C. In Vitro Analysis of Scaffold-free Prevascularized Microtissue Spheroids Containing Human Dental Pulp Cells and Endothelial Cells. J. Endod. 2015, 41, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Levenberg, S.; Rouwkema, J.; Macdonald, M.; Garfein, E.S.; Kohane, D.S.; Darland, D.C.; Marini, R.; Van Blitterswijk, C.A.; Mulligan, R.C.; D’Amore, P.A.; et al. Engineering vascularized skeletal muscle tissue. Nat. Biotechnol. 2005, 23, 879–884. [Google Scholar] [CrossRef]
- Fennema, E.; Rivron, N.; Rouwkema, J.; van Blitterswijk, C.; De Boer, J. Spheroid culture as a tool for creating 3D complex tissues. Trends Biotechnol. 2013, 31, 108–115. [Google Scholar] [CrossRef]
- Rouwkema, J.; Boer, J.D.B.; Van Bitterswijk, C. Endothelial Cells Assemble into a 3-Dimensional Prevascular Network in a Bone Tissue Engineering Construct. Tissue Eng. 2006, 12, 2685–2693. [Google Scholar] [CrossRef]
- Walser, R.; Metzger, W.; Görg, A.; Pohlemann, T.; Menger, M.D.; Laschke, M.W. Generation of co-culture spheroids as vascularisation units for bone tissue engineering. Eur. Cells Mater. 2013, 26, 222–233. [Google Scholar] [CrossRef]
- Gluzman, Z.; Koren, B.; Preis, M.; Cohen, T.; Tsaba, A.; Cosset, F.L.; Shofti, R.; Lewis, B.S.; Virmani, R.; Flugelman, M.Y. Endothelial cells are activated by angiopoeitin-1 gene transfer and produce coordinated sprouting in vitro and arteriogenesis in vivo. Biochem. Biophys. Res. Commun. 2007, 359, 263–268. [Google Scholar] [CrossRef]
- Yoon, C.H.; Choi, Y.E.; Koh, S.J.; Choi, J.-I.; Park, Y.B.; Kim, H.S. High glucose-induced jagged 1 in endothelial cells disturbs notch signaling for angiogenesis: A novel mechanism of diabetic vasculopathy. J. Mol. Cell. Cardiol. 2014, 69, 52–66. [Google Scholar] [CrossRef]
- Blocki, A.; Wang, Y.; Koch, M.; Peh, P.; Beyer, S.; Law, P.; Hui, J.; Raghunath, M. Not all MSCs can act as pericytes: Functional in vitro assays to distinguish pericytes from other mesenchymal stem cells in angiogenesis. Stem Cells Dev. 2013, 22, 2347–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpentier, G.; Berndt, S.; Ferratge, S.; Rasband, W.; Cuendet, M.; Uzan, G.; Albanese, P. Angiogenesis Analyzer for ImageJ—A comparative morphometric analysis of “Endothelial Tube Formation Assay” and “Fibrin Bead Assay”. Sci. Rep. 2020, 10, 11568. [Google Scholar] [CrossRef]
- Martino, M.M.; Hubbell, J.A. The 12th–14th type III repeats of fibronectin function as a highly promiscuous growth factor-binding domain. FASEB J. 2010, 24, 4711–4721. [Google Scholar] [CrossRef]
- Miron-Mendoza, M.; Lin, X.; Ma, L.; Ririe, P.; Petroll, W.M. Individual versus collective fibroblast spreading and migration: Regulation by Matrix Composition in 3D Culture. Exp. Eye Res. 2012, 99, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Tonnesen, M.G.; Mousa, S.A.; Clark, R.A.F. Fibrin and collagen differentially but synergistically regulate sprout angiogenesis of human dermal microvascular endothelial cells in 3-dimensional matrix. Int. J. Cell Biol. 2013, 2013, 231279. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.Y.; Lin, D.; Jarman, E.H.; Polacheck, W.J.; Baker, B.M. Functional angiogenesis requires microenvironmental cues balancing endothelial cell migration and proliferation. Lab Chip 2020, 20, 1153–1166. [Google Scholar] [CrossRef] [PubMed]
- Shanbhag, S.; Rashad, A.; Nymark, E.H.; Suliman, S.; de Lange Davies, C.; Stavropoulos, A.; Bolstad, A.I.; Mustafa, K. Spheroid Coculture of Human Gingiva-Derived Progenitor Cells with Endothelial Cells in Modified Platelet Lysate Hydrogels. Front. Bioeng. Biotechnol. 2021, 9, 739225. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Lee, J.; Song, M.; Kim, M.; Hwang, S.J.; Jang, H.; Park, Y. Combination of three angiogenic growth factors has synergistic effects on sprouting of endothelial cell / mesenchymal stem cell-based spheroids in a 3D matrix. J. Biomed. Mater. Res. B Appl. Biomater. 2015, 104, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Correa de Sampaio, P.; Auslaender, D.; Krubasik, D.; Failla, A.V.; Skepper, J.N.; Murphy, G.; English, W.R. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS ONE 2012, 7, e30753. [Google Scholar] [CrossRef] [Green Version]
- Ehsan, S.M.; Welch-Reardon, K.M.; Waterman, M.L.; Hughes, C.C.W.; George, S.C. A three-dimensional in vitro model of tumor cell intravasation. Integr. Biol. 2014, 6, 603–610. [Google Scholar] [CrossRef]
- Rahimnejad, M.; Nasrollahi Boroujeni, N.; Jahangiri, S.; Rabiee, N.; Rabiee, M.; Makvandi, P.; Akhavan, O.; Varma, R.S. Prevascularized Micro-/Nano-Sized Spheroid/Bead Aggregates for Vascular Tissue Engineering. Nano-Micro Lett. 2021, 13, 182. [Google Scholar] [CrossRef]
- De Moor, L.; Smet, J.; Plovyt, M.; Bekaert, B.; Vercruysse, C.; Asadian, M.; De Geyter, N.; Van Vlierberghe, S.; Dubruel, P.; Declercq, H. Engineering microvasculature by 3D bioprinting of prevascularized spheroids in photo-crosslinkable gelatin. Biofabrication 2021, 13, 045021. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.; Meeks, C.; Vézina, A.; Robey, R.W.; Tanner, K.; Gottesman, M.M. Model systems for studying the blood-brain barrier: Applications and challenges. Biomaterials 2019, 214, 119217. [Google Scholar] [CrossRef]
- Lee, S.; Chung, M.; Lee, S.-R.; Jeon, N.L. 3D brain angiogenesis model to reconstitute functional human blood–brain barrier in vitro. Biotechnol. Bioeng. 2020, 117, 748–762. [Google Scholar] [CrossRef]
- Yarygin, K.N.; Namestnikova, D.D.; Sukhinich, K.K.; Gubskiy, I.L.; Majouga, A.G.; Kholodenko, I.V. Cell Therapy of Stroke: Do the Intra-Arterially Transplanted Mesenchymal Stem Cells Cross the Blood–Brain Barrier? Cells 2021, 10, 2997. [Google Scholar] [CrossRef]
- Hatakeyama, M.; Ninomiya, I.; Kanazawa, M. Angiogenesis and neuronal remodeling after ischemic stroke. Neural Regen. Res. 2020, 15, 16–19. [Google Scholar] [CrossRef]
- Tetzlaff, F.; Fischer, A. Human Endothelial Cell Spheroid-based Sprouting Angiogenesis Assay in Collagen. Bio-Protoc. 2018, 8, e2995. [Google Scholar] [CrossRef]
- Pfisterer, L.; Korff, T. Spheroid-Based In Vitro Angiogenesis Model. Methods Mol. Biol. 2016, 1430, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Nie, M.; Miura, S.; Takeuchi, S. Microfluidic Device for the Analysis of Angiogenic Sprouting under Bidirectional Biochemical Gradients. Micromachines 2020, 11, 1049. [Google Scholar] [CrossRef]
- Hiepen, C.; Osterland, M.; Schütte, C.; Zwijsen, A.; Knaus, P. Role of bone morphogenetic proteins in sprouting angiogenesis: Differential BMP receptor-dependent signaling pathways balance stalk vs. tip cell competence. FASEB J. 2017, 31, 4720–4733. [Google Scholar] [CrossRef] [Green Version]
- Eglinger, J.; Karsjens, H.; Lammert, E. Quantitative assessment of angiogenesis and pericyte coverage in human cell-derived vascular sprouts. Inflamm. Regen. 2017, 37, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
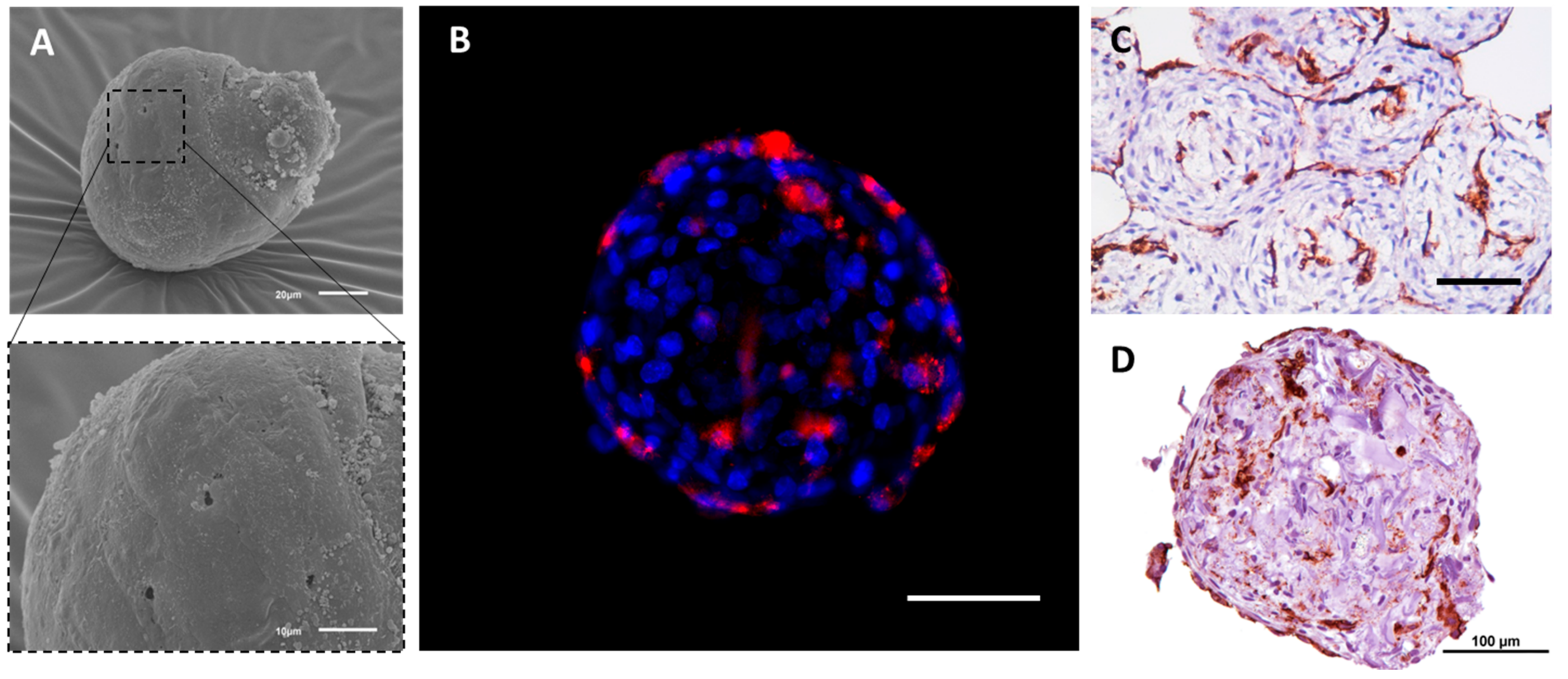{kind=link}
{kind=link}
| Author(s) | Year | Type of ECs | Type of Perivascular Cells | Hydrogel | EC-Perivascular Sprouting Regulation in 3D Coculture Spheroid Sprouting Model (Main Findings) | Reference |
|---|---|---|---|---|---|---|
| Korff T. et al. | 2001 | HUVECs | Umbilical artery SMCs | Collagen (from rat tails) | ECs form layer of cells on the surface of the coculture spheroid. The presence of SMCs decreases EC sensitiveness to VEGF and bFGF stimulation and apoptotic rate in long-term culture. | [16] |
| Wenger et al. | 2004 | HUVECs | Osteoblasts | Collagen | Osteoblasts decreased ECs sensitiveness to VEGFR and bFGF stimulation. The sprouts in co-culture spheroids are mainly composed of osteoblasts and do not form lumens comparing to ECs monoculture spheroids. | [82] |
| Wenger A. et al. | 2005 | HUVECs | Dermal fibroblasts | Collagen | Coculture with osteoblasts decreased heterotypic spheroid sprouting compared to ECs monoculture spheroids. The inhibitory effect of fibroblasts was not mediated by paracrine regulation. | [80] |
| Gluzman Z. et al. | 2007 | Saphenous vein ECs | Saphenous vein SMCs | Collagen | The activation of Ang-1 in ECs and VEGF in SMCs in coculture sprouting model leads to extensive sprouting. | [90] |
| Witz et al. | 2008 | HUVECs, liver sinusoidal ECs (LSECs) | Umbilical artery SMCs | Collagen (from rat tails) | Coculture with SMCs inhibited HUVECs sensitiveness to VEGF and bFGF stimulation. In contrast to HUVECs, LSECs sensitiveness was not affected by coculture with SMCs. | [84] |
| Chang et al. | 2013 | HUVECs | Placenta-derived pericytes | Collagen | PCs promote endothelial sprouting by elaborating HGF, but when recruited to invest endothelial sprouts by PDGF-BB, limit the extent of sprouting in vitro. | [75] |
| Blocki et al. | 2013 | HUVECs | Placenta-derived pericytes/ bone marrow-derived MSCs /fetal lung fibroblast cell line IMR-90 | Collagen | Placenta-derived pericytes improved the integrity of sprouts, while MSCs and fibroblasts migrated larger distances away from ECs and, thus, segregating from sprouts. | [92] |
| Chang-Hwan et al. | 2014 | ECs from mesenteric and gastroepiploic arteries | SMCs from mesenteric and gastroepiploic arteries | Collagen | Vessel growth was aligned with MSCs expressing PDGFR-β (a pericyte marker). When MSCs were depleted after lumen formation, vascular structures were collapsed. | [91] |
| Kim et al. | 2015 | HUVECs | Cord blood-derived MSCs | Collagen | The combination of three angiogenic GFs PDGF+VEGF+FGF increases the length and number of branches comparing to mono VEGF and dual combinations (VEGF+FGF and VEGF+PDGF). | [99] |
| Robinson et al. | 2016 | HUVECs | MSCs (undefined) | Fibrin/PL gel | MSCs influence HUVECs sprouting ability decreasing it. | [74] |
| Bauman et al. | 2018 | Umbilical cord OECs | Bone marrow-derived MSCs | Fibrin | OEC-MSC coculture spheroids deposit ECM (fibronectin, collagen type IV) and exhibit robust sprouting. | [81] |
| Shah et al. | 2018 | Endothelial progenitor cells | Bone marrow-derived MSCs | Collagen | ECs coculture with perivascular cells in spheroids exhibits IC50 of vatalanib (VEGF inhibitor) correlating with in vivo results | [34] |
| Shah et al. | 2019 | Human ECFCs | Bone marrow-derived MSCs | Collagen | The presence of perivascular cells in coculture spheroids affects the sensitiveness to bevacizumab (VEGF inhibitor) which results in relevant IC50 in comparison to EC monoculture spheroids. | [35] |
| Barnes et al. | 2019 | Pulmonary arterial ECs from healthy donors and patients with idiopathic pulmonary arterial hypertension (IPAH) | Pulmonary arterial SMCs from healthy donors and patients with IPAH | Collagen type I (from rat tails) | ECs and SMCs from patients with IPAH in coculture spheroids exhibit more robust sprouting due to altered glucose uptake and dysregulation in OGT/O-GlcNAc axis. | [36] |
| Vorwand et al. | 2020 | Human ECFCs | Bone marrow-derived MSCs | Fibrin | Spatial localization of ECs within EC-MSC co-culture spheroids affects sprouting potential and NOTCH3 expression. | [37] |
| Zhang et al. | 2021 | HUVECs | Dental pulp MSCs | Collagen type I | Pre-treatment with TGF-β1 affects angiogenic properties of dental pulp MSCs on coculture spheroid sprouting assay in comparison to untreated MSCs. | [48] |
| Shanbhag et al. | 2021 | HUVECs | GPCs | PL gel | Direct coculture with GPCs in mixed spheroids in a 5:1 ratio significantly improves HUVEC sprouting. | [98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vakhrushev, I.V.; Nezhurina, E.K.; Karalkin, P.A.; Tsvetkova, A.V.; Sergeeva, N.S.; Majouga, A.G.; Yarygin, K.N. Heterotypic Multicellular Spheroids as Experimental and Preclinical Models of Sprouting Angiogenesis. Biology 2022, 11, 18. https://doi.org/10.3390/biology11010018
Vakhrushev IV, Nezhurina EK, Karalkin PA, Tsvetkova AV, Sergeeva NS, Majouga AG, Yarygin KN. Heterotypic Multicellular Spheroids as Experimental and Preclinical Models of Sprouting Angiogenesis. Biology. 2022; 11(1):18. https://doi.org/10.3390/biology11010018
Chicago/Turabian StyleVakhrushev, Igor V., Elizaveta K. Nezhurina, Pavel A. Karalkin, Anastasia V. Tsvetkova, Nataliya S. Sergeeva, Alexander G. Majouga, and Konstantin N. Yarygin. 2022. "Heterotypic Multicellular Spheroids as Experimental and Preclinical Models of Sprouting Angiogenesis" Biology 11, no. 1: 18. https://doi.org/10.3390/biology11010018