
Influence of Varying Dietary ω6 to ω3 Fatty Acid Ratios on the Hepatic Transcriptome, and Association with Phenotypic Traits (Growth, Somatic Indices, and Tissue Lipid Composition), in Atlantic Salmon (Salmo salar)

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Experimental Diets
2.2. Sample Collection
2.3. RNA Extraction, DNase Treatment, Column Purification and cDNA Synthesis
2.4. Microarray Hybridization and Data Acquisition
2.5. Microarray Data Analysis
2.6. qPCR Study and Data Analysis
2.7. Statistical Analyses
2.7.1. qPCR Data
2.7.2. Correlation Analyses of qPCR and Lipid Composition Data
3. Results
3.1. Liver Microarray Analysis
3.2. qPCR Study
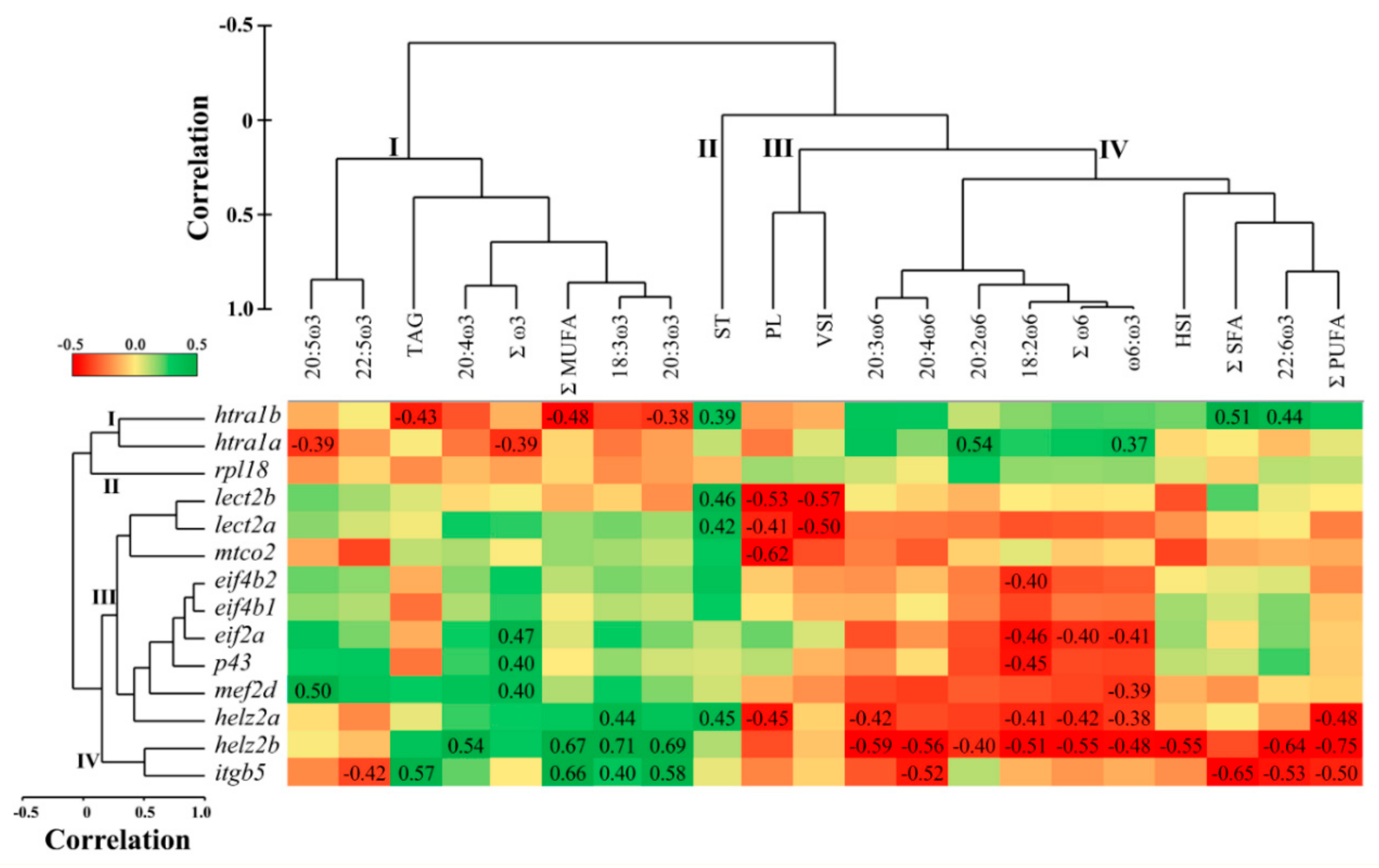
3.3. Correlations between Hepatic qPCR Transcript Expression and Liver Lipid Composition
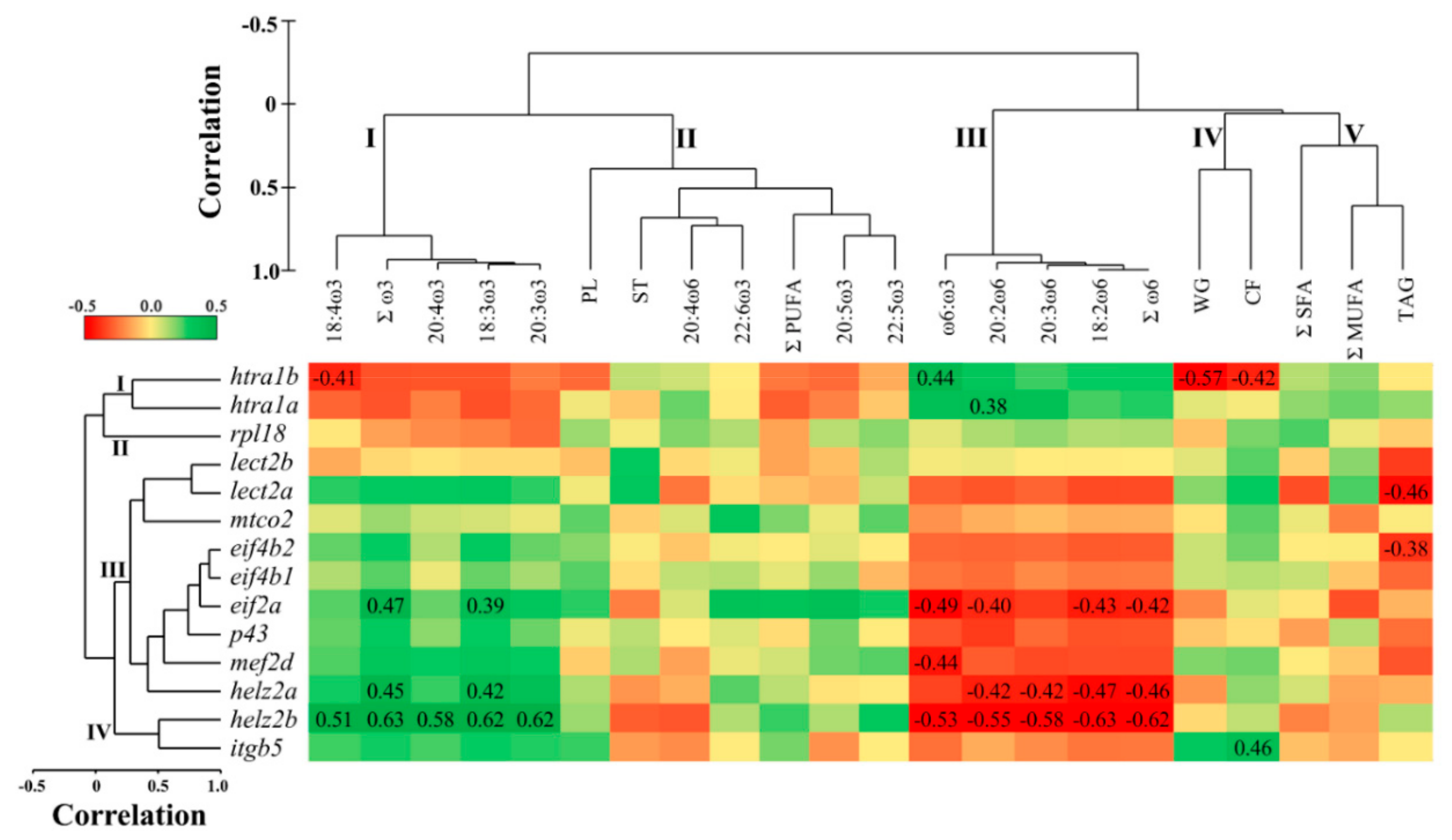
3.4. Correlations between Hepatic qPCR Transcript Expression and Muscle Lipid Composition
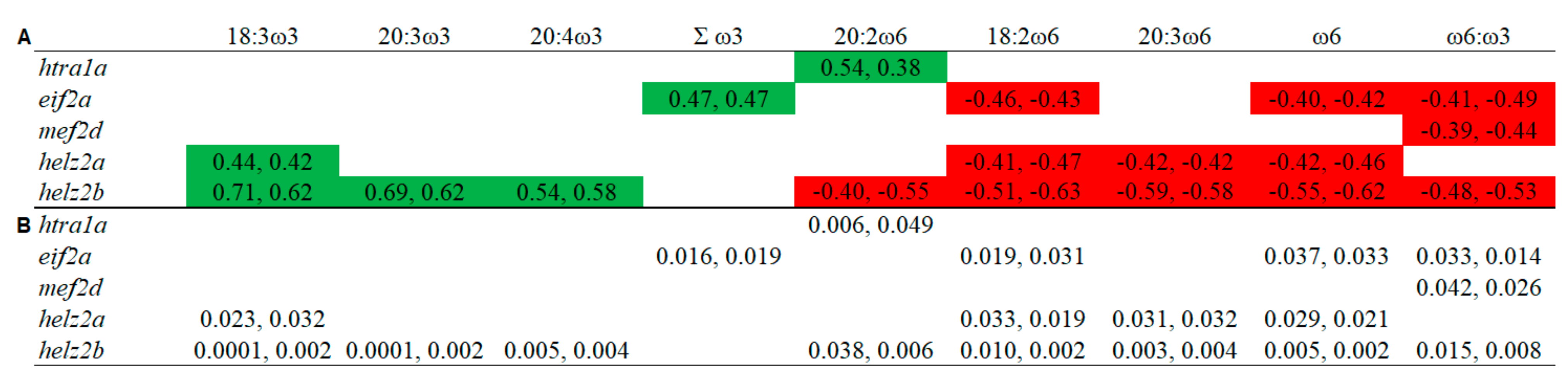
3.5. Overlapping Lipid–Gene Correlations between the Liver and Muscle Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilberg, M.J.; Miller, T.J. Comment on “Impacts of biodiversity loss on ocean ecosystem services”. Science 2007, 316, 787–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding aquaculture in an era of finite resources. Proc. Natl. Acad. Sci. USA 2009, 106, 15103–15110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchini, G.M.; Francis, D.S. Fatty acid metabolism (desaturation, elongation and β-oxidation) in rainbow trout fed fish oil- or linseed oil-based diets. Br. J. Nutr. 2009, 102, 69–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bransden, M.P.; Carter, C.G.; Nichols, P.D. Replacement of fish oil with sunflower oil in feeds for Atlantic salmon (Salmo salar L.): Effect on growth performance, tissue fatty acid composition and disease resistance. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 135, 611–625. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Liland, N.S.; Rosenlund, G.; Berntssen, M.H.G.; Brattelid, T.; Madsen, L.; Torstensen, B.E. Net production of Atlantic salmon (FIFO, Fish in Fish out <1) with dietary plant proteins and vegetable oils. Aquac. Nutr. 2013, 19, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Tocher, D.R. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Alhazzaa, R.; Bridle, A.R.; Nichols, P.D.; Carter, C.G. Replacing dietary fish oil with Echium oil enriched barramundi with C18 PUFA rather than long-chain PUFA. Aquaculture 2011, 312, 162–171. [Google Scholar] [CrossRef]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Sprague, M.; Dick, J.R.; Tocher, D.R. Impact of sustainable feeds on omega-3 long-chain fatty acid levels in farmed Atlantic salmon, 2006–2015. Sci. Rep. 2016, 6, 21892. [Google Scholar] [CrossRef] [Green Version]
- Montero, D.; Kalinowski, T.; Obach, A.; Robaina, L.; Tort, L.; Caballero, M.J.; Izquierdo, M.S. Vegetable lipid sources for gilthead seabream (Sparus aurata): Effects on fish health. Aquaculture 2003, 225, 353–370. [Google Scholar] [CrossRef]
- Ruyter, B.; Moya-Falcón, C.; Rosenlund, G.; Vegusdal, A. Fat content and morphology of liver and intestine of Atlantic salmon (Salmo salar): Effects of temperature and dietary soybean oil. Aquaculture 2006, 252, 441–452. [Google Scholar] [CrossRef]
- Jordal, A.E.O.; Lie, Ø.; Torstensen, B.E. Complete replacement of dietary fish oil with a vegetable oil blend affect liver lipid and plasma lipoprotein levels in Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2007, 13, 114–130. [Google Scholar] [CrossRef]
- Liland, N.S. Atlantic Salmon (Salmo salar L.) Sterol Metabolism and Metabolic Health Impact of Dietary Lipids. Ph.D. Thesis, Department of Biology, University of Bergen, National Institute of Nutrition and Seafood Research, Bergen, Norway, December 2014. [Google Scholar]
- Caballero-Solares, A.; Hall, J.R.; Xue, X.; Eslamloo, K.; Taylor, R.G.; Parrish, C.C.; Rise, M.L. The dietary replacement of marine ingredients by terrestrial animal and plant alternatives modulates the antiviral immune response of Atlantic salmon (Salmo salar). Fish Shellfish Immunol. 2017, 64, 24–38. [Google Scholar] [CrossRef]
- Pickova, J.; Mørkøre, T. Alternate oils in fish feeds. Eur. J. Lipid Sci. Technol. 2007, 109, 256–263. [Google Scholar] [CrossRef]
- Weaver, K.L.; Ivester, P.; Chilton, J.A.; Wilson, M.D.; Pandey, P.; Chilton, F.H. The content of favorable and unfavorable polyunsaturated fatty acids found in commonly eaten fish. J. Am. Diet. Assoc. 2008, 108, 1178–1185. [Google Scholar] [CrossRef]
- Young, K. Omega-6 (n-6) and omega-3 (n-3) fatty acids in tilapia and human health: A review. Int. J. Food Sci. Nutr. 2009, 60 (Suppl. 5), 203–211. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Importance of the ratio of omega-6/omega-3 essential fatty acids: Evolutionary aspects. World Rev. Nutr. Diet. 2003, 92, 1–22. [Google Scholar] [CrossRef]
- Wijendran, V.; Hayes, K.C. Dietary n-6 and n-3 fatty acid balance and cardiovascular health. Annu. Rev. Nutr. 2004, 24, 597–615. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Gómez Candela, C.; Bermejo López, L.M.; Loria Kohen, V. Importance of a balanced omega 6/omega 3 ratio for the maintenance of health: Nutritional recommendations. Nutr. Hosp. 2011, 26, 323–329. [Google Scholar] [CrossRef]
- De Pablo Martinez, M.A.; Álvarez De Cienfuegos, G. Modulatory effects of dietary lipids on immune system functions. Immunol. Cell Biol. 2000, 78, 31–39. [Google Scholar] [CrossRef]
- Wymann, M.P.; Schneiter, R. Lipid signalling in disease. Nat. Rev. Mol. Cell Biol. 2008, 9, 162–176. [Google Scholar] [CrossRef]
- Duan, Y.; Li, F.; Li, L.; Fan, J.; Sun, X.; Yin, Y. n-6:n-3 PUFA ratio is involved in regulating lipid metabolism and inflammation in pigs. Br. J. Nutr. 2014, 111, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Katan, T.; Caballero-Solares, A.; Taylor, R.G.; Rise, M.L.; Parrish, C.C. Effect of plant-based diets with varying ratios of ω6 to ω3 fatty acids on growth performance, tissue composition, fatty acid biosynthesis and lipid-related gene expression in Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 30, 290–304. [Google Scholar] [CrossRef]
- Jantzen, S.G.; Sanderson, D.S.; Von Schalburg, K.R.; Yasuike, M.; Marass, F.; Koop, B.F. A 44K microarray dataset of the changing transcriptome in developing Atlantic salmon (Salmo salar L.). BMC Res. Notes 2011, 4, 88. [Google Scholar] [CrossRef] [Green Version]
- Sahlmann, C.; Sutherland, B.J.G.; Kortner, T.M.; Koop, B.F.; Krogdahl, Å.; Bakke, A.M. Early response of gene expression in the distal intestine of Atlantic salmon (Salmo salar L.) during the development of soybean meal induced enteritis. Fish Shellfish Immunol. 2013, 34, 599–609. [Google Scholar] [CrossRef]
- Xue, X.; Hixson, S.M.; Hori, T.S.; Booman, M.; Parrish, C.C.; Anderson, D.M.; Rise, M.L. Atlantic salmon (Salmo salar) liver transcriptome response to diets containing Camelina sativa products. Comp. Biochem. Physiol. Part D Genom. Proteom. 2015, 14, 1–15. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Xu, Q.; Feng, C.Y.; Hori, T.S.; Plouffe, D.A.; Buchanan, J.T.; Rise, M.L. Family-specific differences in growth rate and hepatic gene expression in juvenile triploid growth hormone (GH) transgenic Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. Part D Genom. Proteom. 2013, 8, 317–333. [Google Scholar] [CrossRef]
- Booman, M.; Borza, T.; Feng, C.Y.; Hori, T.S.; Higgins, B.; Culf, A.; Léger, D.; Chute, I.C.; Belkaid, A.; Rise, M.L.; et al. Development and experimental validation of a 20K Atlantic cod (Gadus morhua) oligonucleotide microarray based on a collection of over 150,000 ESTs. Mar. Biotechnol. 2011, 13, 733–750. [Google Scholar] [CrossRef] [Green Version]
- Celton, M.; Malpertuy, A.; Lelandais, G.; de Brevern, A.G. Comparative analysis of missing value imputation methods to improve clustering and interpretation of microarray experiments. BMC Genom. 2010, 11, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bø, T.H.; Dysvik, B.; Jonassen, I. LSimpute: Accurate estimation of missing values in microarray data with least squares methods. Nucleic Acids Res. 2004, 32, e34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwender, H.; Krause, A.; Ickstadt, K. Identifying interesting genes with siggenes. RNews 2006, 6, 45–50. [Google Scholar]
- Breitling, R.; Armengaud, P.; Amtmann, A.; Herzyk, P. Rank products: A simple, yet powerful, new method to detect differentially regulated genes in replicated microarray experiments. FEBS Lett. 2004, 573, 83–92. [Google Scholar] [CrossRef]
- Jeffery, I.B.; Higgins, D.G.; Culhane, A.C. Comparison and evaluation of methods for generating differentially expressed gene lists from microarray data. BMC Bioinform. 2006, 7, 359. [Google Scholar] [CrossRef]
- Hong, F.; Breitling, R.; McEntee, C.W.; Wittner, B.S.; Nemhauser, J.L.; Chory, J. RankProd: A bioconductor package for detecting differentially expressed genes in meta-analysis. Bioinformatics 2006, 22, 2825–2827. [Google Scholar] [CrossRef] [Green Version]
- Caballero-Solares, A.; Xue, X.; Parrish, C.C.; Foroutani, M.B.; Taylor, R.G.; Rise, M.L. Changes in the liver transcriptome of farmed Atlantic salmon (Salmo salar) fed experimental diets based on terrestrial alternatives to fish meal and fish oil. BMC Genom. 2018, 19, 796. [Google Scholar] [CrossRef] [Green Version]
- Rise, M.L.; Hall, J.R.; Rise, M.; Hori, T.S.; Browne, M.J.; Gamperl, A.K.; Hubert, S.; Kimball, J.; Bowman, S.; Johnson, S.C. Impact of asymptomatic nodavirus carrier state and intraperitoneal viral mimic injection on brain transcript expression in Atlantic cod (Gadus morhua). Physiol. Genom. 2010, 42, 266–280. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Olsvik, P.A.; Lie, K.K.; Jordal, A.E.O.; Nilsen, T.O.; Hordvik, I. Evaluation of potential reference genes in real-time RT-PCR studies of Atlantic salmon. BMC Mol. Biol. 2005, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rise, M.L.; Hall, J.R.; Nash, G.W.; Xue, X.; Booman, M.; Katan, T.; Gamperl, A.K. Transcriptome profiling reveals that feeding wild zooplankton to larval Atlantic cod (Gadus morhua) influences suites of genes involved in oxidation-reduction, mitosis, and selenium homeostasis. BMC Genom. 2015, 16, 1016. [Google Scholar] [CrossRef] [Green Version]
- Booman, M.; Xu, Q.; Rise, M.L. Evaluation of the impact of camelina oil-containing diets on the expression of genes involved in the innate anti-viral immune response in Atlantic cod (Gadus morhua). Fish Shellfish Immunol. 2014, 41, 52–63. [Google Scholar] [CrossRef]
- Brown, T.D.; Hori, T.S.; Xue, X.; Ye, C.L.; Anderson, D.M.; Rise, M.L. Functional genomic analysis of the impact of Camelina (Camelina sativa) meal on Atlantic salmon (Salmo salar) distal intestine gene expression and physiology. Mar. Biotechnol. 2016, 18, 418–435. [Google Scholar] [CrossRef] [Green Version]
- Eslamloo, K.; Xue, X.; Hall, J.R.; Smith, N.C.; Caballero-Solares, A.; Parrish, C.C.; Taylor, R.G.; Rise, M.L. Transcriptome profiling of antiviral immune and dietary fatty acid dependent responses of Atlantic salmon macrophage-like cells. BMC Genom. 2017, 18, 706. [Google Scholar] [CrossRef]
- Morais, S.; Pratoomyot, J.; Taggart, J.B.; Bron, J.E.; Guy, D.R.; Bell, J.G.; Tocher, D.R. Genotype-specific responses in Atlantic salmon (Salmo salar) subject to dietary fish oil replacement by vegetable oil: A liver transcriptomic analysis. BMC Genom. 2011, 12, 255. [Google Scholar] [CrossRef] [Green Version]
- Leaver, M.J.; Villeneuve, L.A.N.; Obach, A.; Jensen, L.; Bron, J.E.; Tocher, D.R.; Taggart, J.B. Functional genomics reveals increases in cholesterol biosynthetic genes and highly unsaturated fatty acid biosynthesis after dietary substitution of fish oil with vegetable oils in Atlantic salmon (Salmo salar). BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Surapureddi, S.; Yu, S.; Bu, H.; Hashimoto, T.; Yeldandi, A.V.; Kashireddy, P.; Cherkaoui-Malki, M.; Qi, C.; Zhu, Y.J.; Rao, M.S.; et al. Identification of a transcriptionally active peroxisome proliferator-activated receptor α-interacting cofactor complex in rat liver and characterization of PRIC285 as a coactivator. Proc. Natl. Acad. Sci. USA 2002, 99, 11836–11841. [Google Scholar] [CrossRef] [Green Version]
- Katano-Toki, A.; Satoh, T.; Tomaru, T.; Yoshino, S.; Ishizuka, T.; Ishii, S.; Ozawa, A.; Shibusawa, N.; Tsuchiya, T.; Saito, T.; et al. THRAP3 interacts with HELZ2 and plays a novel role in adipocyte differentiation. Mol. Endocrinol. 2013, 27, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, S.; Satoh, T.; Yamada, M.; Hashimoto, K.; Tomaru, T.; Katano-Toki, A.; Kakizaki, S.; Okada, S.; Shimizu, H.; Ozawa, A.; et al. Protection against high-fat diet-induced obesity in Helz2-deficient male mice due to enhanced expression of hepatic leptin receptor. Endocrinology 2014, 155, 3459–3472. [Google Scholar] [CrossRef] [Green Version]
- Kersten, S. Integrated physiology and systems biology of PPARα. Mol. Metab. 2014, 3, 354–371. [Google Scholar] [CrossRef]
- Fusco, D.N.; Pratt, H.; Kandilas, S.; Cheon, S.S.Y.; Lin, W.; Cronkite, D.A.; Basavappa, M.; Jeffrey, K.L.; Anselmo, A.; Sadreyev, R.; et al. HELZ2 is an IFN effector mediating suppression of dengue virus. Front. Microbiol. 2017, 8, 240. [Google Scholar] [CrossRef]
- Fu, M.; Blackshear, P.J. RNA-binding proteins in immune regulation: A focus on CCCH zinc finger proteins. Nat. Rev. Immunol. 2017, 17, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Levraud, J.; Jouneau, L.; Briolat, V.; Laghi, V.; Boudinot, P. IFN-stimulated genes in zebrafish and humans define an ancient arsenal of antiviral immunity. J. Immunol. 2019, 203, 3361–3373. [Google Scholar] [CrossRef]
- Kersten, S.; Desvergne, B.; Wahli, W. Roles of PPARS in health and disease. Nature 2000, 405, 421–424. [Google Scholar] [CrossRef]
- Van Raalte, D.H.; Li, M.; Pritchard, P.H.; Wasan, K.M. Peroxisome proliferator-activated receptor (PPAR)-α: A pharmacological target with a promising future. Pharm. Res. 2004, 21, 1531–1538. [Google Scholar] [CrossRef]
- Huang, H.; Starodub, O.; McIntosh, A.; Kier, A.B.; Schroeder, F. Liver fatty acid-binding protein targets fatty acids to the nucleus. Real time confocal and multiphoton fluorescence imaging in living cells. J. Biol. Chem. 2002, 277, 29139–29151. [Google Scholar] [CrossRef] [Green Version]
- Hostetler, H.A.; McIntosh, A.L.; Atshaves, B.P.; Storey, S.M.; Payne, H.R.; Kier, A.B.; Schroeder, F. L-FABP directly interacts with PPARα in cultured primary hepatocytes. J. Lipid Res. 2009, 50, 1663–1675. [Google Scholar] [CrossRef] [Green Version]
- Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptors: Nuclear control of metabolism. Endocr. Rev. 1999, 20, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, F.; Petrescu, A.D.; Huang, H.; Atshaves, B.P.; McIntosh, A.L.; Martin, G.G.; Hostetler, H.A.; Vespa, A.; Landrock, D.; Landrock, K.K.; et al. Role of fatty acid binding proteins and long chain fatty acids in modulating nuclear receptors and gene transcription. Lipids 2008, 43, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ticho, B.S.; Stainier, D.Y.R.; Fishman, M.C.; Breitbart, R.E. Three zebrafish MEF2 genes delineate somitic and cardiac muscle development in wild-type and mutant embryos. Mech. Dev. 1996, 59, 205–218. [Google Scholar] [CrossRef]
- He, M.; Zhou, D.; Ding, N.Z.; Teng, C.B.; Yan, X.C.; Liang, Y. Common carp MEF2 genes: Evolution and expression. Genes 2019, 10, 588. [Google Scholar] [CrossRef] [Green Version]
- Black, B.L.; Olson, E.N. Transcriptional control of muscle development by myocyte enhancer factor-2 (MEF2) proteins. Annu. Rev. Cell Dev. Biol. 1998, 14, 167–196. [Google Scholar] [CrossRef]
- Flavell, S.W.; Cowan, C.W.; Kim, T.K.; Greer, P.L.; Lin, Y.; Paradis, S.; Griffith, E.C.; Hu, L.S.; Chen, C.; Greenberg, M.E. Activity-dependent regulation of MEF2 transcription factors suppresses excitatory synapse number. Science 2006, 311, 1008–1012. [Google Scholar] [CrossRef] [Green Version]
- Haberland, M.; Arnold, M.A.; McAnally, J.; Phan, D.; Kim, Y.; Olson, E.N. Regulation of HDAC9 gene expression by MEF2 establishes a negative-feedback loop in the transcriptional circuitry of muscle differentiation. Mol. Cell. Biol. 2007, 27, 518–525. [Google Scholar] [CrossRef] [Green Version]
- Flavell, S.W.; Kim, T.K.; Gray, J.M.; Harmin, D.A.; Hemberg, M.; Hong, E.J.; Markenscoff-Papadimitriou, E.; Bear, D.M.; Greenberg, M.E. Genome-wide analysis of MEF2 transcriptional program reveals synaptic target genes and neuronal activity-dependent polyadenylation site selection. Neuron 2008, 60, 1022–1038. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Zhou, Y.; Jiang, S.; Huang, F.; Peng, J.; Jiang, S. Transcriptional response of porcine skeletal muscle to feeding a linseed-enriched diet to growing pigs. J. Anim. Sci. Biotechnol. 2016, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Grammes, F.; Rørvik, K.A.; Takle, H. Tetradecylthioacetic acid modulates cardiac transcription in Atlantic salmon, Salmo salar L., suffering heart and skeletal muscle inflammation. J. Fish Dis. 2012, 35, 109–117. [Google Scholar] [CrossRef]
- Xiaomeng, M.; Zhang, L.; Lixin, D. Research progress on MEF2B gene in human and animals. Agric. Sci. Technol. 2016, 17, 2477–2482. [Google Scholar]
- Wang, X.; Tang, X.; Gong, X.; Albanis, E.; Friedman, S.L.; Mao, Z. Regulation of hepatic stellate cell activation and growth by transcription factor myocyte enhancer factor 2. Gastroenterology 2004, 127, 1174–1188. [Google Scholar] [CrossRef]
- Zhang, K.; Chang, Y.; Shi, Z.; Han, X.; Han, Y.; Yao, Q. ω-3 PUFAs ameliorate liver fibrosis and inhibit hepatic stellate cells proliferation and activation by promoting YAP/TAZ degradation. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, E.N.; Valdés, J.A.; Molina, A.; Björnsson, B.T. Regulation of skeletal muscle growth in fish by the growth hormone—Insulin-like growth factor system. Gen. Comp. Endocrinol. 2013, 192, 136–148. [Google Scholar] [CrossRef]
- Lien, C.L.; Schebesta, M.; Makino, S.; Weber, G.J.; Keating, M.T. Gene expression analysis of zebrafish heart regeneration. PLoS Biol. 2006, 4, 1386–1396. [Google Scholar] [CrossRef]
- Clausen, T.; Southan, C.; Ehrmann, M. The HtrA family of proteases: Implications for protein composition and cell fate. Mol. Cell 2002, 10, 443–455. [Google Scholar] [CrossRef]
- Oka, C.; Tsujimoto, R.; Kajikawa, M.; Koshiba-Takeuchi, K.; Ina, J.; Yano, M.; Tsuchiya, A.; Ueta, Y.; Soma, A.; Kanda, H.; et al. HtrA1 serine protease inhibits signaling mediated by Tgfβ family proteins. Development 2004, 131, 1041–1053. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.R.; Chamberland, A.; Lin, Q.; Li, X.J.; Dai, D.; Zeng, W.; Ryan, M.S.; Rivera-Bermúdez, M.A.; Flannery, C.R.; Yang, Z. Serine protease HTRA1 antagonizes transforming growth factor-β signaling by cleaving its receptors and loss of HTRA1 in vivo enhances bone formation. PLoS ONE 2013, 8, e74094. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Jin, L.; Luo, T.P.; Luo, G.H.; Tan, Y.; Qin, X.H. Serine protease HtrA1 expression in human hepatocellular carcinoma. Hepatobiliary Pancreat. Dis. Int. 2010, 9, 508–512. [Google Scholar]
- Tacchi, L.; Secombes, C.J.; Bickerdike, R.; Adler, M.A.; Venegas, C.; Takle, H.; Martin, S.A.M. Transcriptomic and physiological responses to fishmeal substitution with plant proteins in formulated feed in farmed Atlantic salmon (Salmo salar). BMC Genom. 2012, 13, 363. [Google Scholar] [CrossRef] [Green Version]
- Bakay, M.; Zhao, P.; Chen, J.; Hoffman, E.P. A web-accessible complete transcriptome of normal human and DMD muscle. Neuromuscul. Disord. 2002, 12, S125–S141. [Google Scholar] [CrossRef]
- Tiaden, A.N.; Richards, P.J. The emerging roles of HTRA1 in musculoskeletal disease. Am. J. Pathol. 2013, 182, 1482–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokkinos, P.A.; Kazantzi, A.; Sfyroera, G.; Zarkadis, I.K. Molecular cloning of leukocyte cell-derived chemotaxin 2 in rainbow trout. Fish Shellfish Immunol. 2005, 18, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Guo, M.; Cui, H.; Yan, Y.; Ouyang, Z.; Qin, Q. A new leukocyte cell-derived chemotaxin-2 from marine fish grouper, Epinephelus coioides: Molecular cloning and expression analysis. Fish Shellfish Immunol. 2011, 31, 600–605. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Q.; Lu, X.J.; Li, C.H. LECT2 improves the outcomes in ayu with Vibrio anguillarum infection via monocytes/macrophages. Fish Shellfish Immunol. 2014, 41, 586–592. [Google Scholar] [CrossRef]
- Fu, G.H.; Bai, Z.Y.; Xia, J.H.; Liu, X.J.; Liu, F.; Wan, Z.Y.; Yue, G.H. Characterization of the LECT2 gene and its associations with resistance to the big belly disease in Asian seabass. Fish Shellfish Immunol. 2014, 37, 131–138. [Google Scholar] [CrossRef]
- Lan, F.; Misu, H.; Chikamoto, K.; Takayama, H.; Kikuchi, A.; Mohri, K.; Takata, N.; Hayashi, H.; Matsuzawa-Nagata, N.; Takeshita, Y.; et al. LECT2 functions as a hepatokine that links obesity to skeletal muscle insulin resistance. Diabetes 2014, 63, 1649–1664. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.W.; Chung, Y.H.; Kim, H.; Elaty, A.M.A.; Jeong, J.H. LECT2 promotes inflammation and insulin resistance in adipocytes via P38 pathways. J. Mol. Endocrinol. 2018, 61, 37–45. [Google Scholar] [CrossRef]
- Todorčević, M.; Škugor, S.; Krasnov, A.; Ruyter, B. Gene expression profiles in Atlantic salmon adipose-derived stromo-vascular fraction during differentiation into adipocytes. BMC Genom. 2010, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Kortner, T.M.; Björkhem, I.; Krasnov, A.; Timmerhaus, G.; Krogdahl, Å. Dietary cholesterol supplementation to a plant-based diet suppresses the complete pathway of cholesterol synthesis and induces bile acid production in Atlantic salmon (Salmo salar L.). Br. J. Nutr. 2014, 111, 2089–2103. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Immunoregulatory and anti-inflammatory effects of n-3 polyunsaturated fatty acids. Braz. J. Med. Biol. Res. 1998, 31, 467–490. [Google Scholar] [CrossRef] [Green Version]
- Pompéia, C.; Lopes, L.R.; Miyasaka, C.K.; Procópio, J.; Sannomiya, P.; Curi, R. Effect of fatty acids on leukocyte function. Braz. J. Med. Biol. Res. 2000, 33, 1255–1268. [Google Scholar] [CrossRef]
- Geay, F.; Ferraresso, S.; Zambonino-Infante, J.L.; Bargelloni, L.; Quentel, C.; Vandeputte, M.; Kaushik, S.; Cahu, C.L.; Mazurais, D. Effects of the total replacement of fish-based diet with plant-based diet on the hepatic transcriptome of two European sea bass (Dicentrarchus labrax) half-sibfamilies showing different growth rates with the plant-based diet. BMC Genom. 2011, 12, 522. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.H.; Wang, J.; Kang, J.X. Decreased n-6/n-3 fatty acid ratio reduces the invasive potential of human lung cancer cells by downregulation of cell adhesion/invasion-related genes. Carcinogenesis 2005, 26, 779–784. [Google Scholar] [CrossRef] [Green Version]
- Ivakhno, S.S.; Kornelyuk, A.I. Cytokine-like activities of some aminoacyl-tRna synthetases and auxiliary p43 cofactor of aminoacylation reaction and their role in oncogenesis. Exp. Oncol. 2004, 26, 250–255. [Google Scholar]
- Ko, Y.G.; Park, H.; Kim, T.; Lee, J.W.; Park, S.G.; Seol, W.; Kim, J.E.; Lee, W.H.; Kim, S.H.; Park, J.E.; et al. A cofactor of mRNA synthetase, p43, is secreted to up-regulate proinflammatory genes. J. Biol. Chem. 2001, 276, 23028–23033. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Park, S.G.; Kim, J.; Ko, Y.G.; Kim, S. Signaling pathways for TNF production induced by human aminoacyl-tRNA synthetase-associating factor, p43. Cytokine 2002, 20, 148–153. [Google Scholar] [CrossRef]
- Sang, G.P.; Shin, H.; Young, K.S.; Lee, Y.; Choi, E.C.; Park, B.J.; Kim, S. The novel cytokine p43 stimulates dermal fibroblast proliferation and wound repair. Am. J. Pathol. 2005, 166, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Le, X.; Pugach, E.K.; Hettmer, S.; Storer, N.Y.; Liu, J.; Wills, A.A.; DiBiase, A.; Chen, E.Y.; Ignatius, M.S.; Poss, K.D.; et al. A novel chemical screening strategy in zebrafish identifies common pathways in embryogenesis and rhabdomyosarcoma development. Development 2012, 140, 2354–2364. [Google Scholar] [CrossRef] [Green Version]
- Faught, E.; Vijayan, M.M. Loss of the glucocorticoid receptor in zebrafish improves muscle glucose availability and increases growth. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E1093–E1104. [Google Scholar] [CrossRef]
- Sen, N.D.; Zhou, F.; Harris, M.S.; Ingolia, N.T.; Hinnebusch, A.G. EIF4B stimulates translation of long mRNAs with structured 5′ UTRs and low closed-loop potential but weak dependence on eIF4G. Proc. Natl. Acad. Sci. USA 2016, 113, 10464–10472. [Google Scholar] [CrossRef] [Green Version]
- Harding, H.P.; Zhang, Y.; Bertolotti, A.; Zeng, H.; Ron, D. Perk is essential for translational regulation and cell survival during the unfolded protein response. Mol. Cell 2000, 5, 897–904. [Google Scholar] [CrossRef]
- Howarth, D.L.; Lindtner, C.; Vacaru, A.M.; Sachidanandam, R.; Tsedensodnom, O.; Vasilkova, T.; Buettner, C.; Sadler, K.C. Activating transcription factor 6 is necessary and sufficient for alcoholic fatty liver disease in zebrafish. PLoS Genet. 2014, 10, e1004335. [Google Scholar] [CrossRef]
- Kavaliauskis, A.; Arnemo, M.; Rishovd, A.L.; Gjøen, T. Activation of unfolded protein response pathway during infectious salmon anemia virus (ISAV) infection in vitro and in vivo. Dev. Comp. Immunol. 2016, 54, 46–54. [Google Scholar] [CrossRef]
- Saelens, X.; Kalai, M.; Vandenabeele, P. Translation inhibition in apoptosis: Caspase-dependent PKR activation and eIF2-α phosphorylation. J. Biol. Chem. 2001, 276, 41620–41628. [Google Scholar] [CrossRef] [Green Version]
- Morley, S.J.; Coldwell, M.J.; Clemens, M.J. Initiation factor modifications in the preapoptotic phase. Cell Death Differ. 2005, 12, 571–584. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rankin, D.; Rennie, M.J.; Mittendorfer, B. Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: A randomized controlled trial. Am. J. Clin. Nutr. 2011, 93, 402–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panserat, S.; Hortopan, G.A.; Plagnes-Juan, E.; Kolditz, C.; Lansard, M.; Skiba-Cassy, S.; Esquerré, D.; Geurden, I.; Médale, F.; Kaushik, S.; et al. Differential gene expression after total replacement of dietary fish meal and fish oil by plant products in rainbow trout (Oncorhynchus mykiss) liver. Aquaculture 2009, 294, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Houlihan, D.F.; Carter, C.G. Protein synthesis. Fish Physiol. 2001, 20, 31–75. [Google Scholar] [CrossRef]
- Kaushik, S.J.; Seiliez, I. Protein and amino acid nutrition and metabolism in fish: Current knowledge and future needs. Aquac. Res. 2010, 41, 322–332. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Name (Symbol) a | Nucleotide Sequence (5′-3′) b | Amplification Efficiency (%) | Amplicon Size (bp) | GenBank Accession Number |
|---|---|---|---|---|
| Serine protease HTRA1 a (htra1a) c | F:GCTGATGTGGTGGAGGAGAT | 113.3 | 127 | NM001141717 |
| R:TCAAGCCGTCCTCTGACAC | - | - | - | |
| Serine protease HTRA1 b (htra1b) c | F:ATGATGACTCTCACACCAATGC | 95.4 | 104 | EG831192 |
| R:GTTTTTGGGATGACCTCGATT | - | - | - | |
| Aminoacyl tRNA synthase complex-interacting multifunctional protein 1 (p43) | F:GGAAGACGAATGCAGAGGAC | 97.2 | 82.0 | BT044000 |
| R:GGAGCGGTCATTCACACTTT | - | - | - | |
| Eukaryotic translation initiation factor 2A (eif2a) | F:TAAACCCAGATGCCCTTGAG | 94.9 | 143 | NM001140088 |
| R:GGCTTTCAGCTCGTCGATAG | - | - | - | |
| Eukaryotic translation initiation factor 4B 1 (eif4b1) | F:CGCAGGGACCGGGATGAT | 85.3 | 123 | BT072661 |
| R:TCGGTCCTC5CTGTCCGC | - | - | - | |
| Eukaryotic translation initiation factor 4B 2 (eif4b2) | F:CACATCCAGGAAGTACCTCT | 87.4 | 94.0 | DY739566 |
| R:TCGTCCTCCTTACCGCTGA | - | - | - | |
| Cytochrome c oxidase subunit 2 (mtco2) d | F:CACCGATTACGAAGACTTAGGC | 107.9 | 136 | DW554935 |
| R:TGAAACTAGGACCCGGATTG | - | - | - | |
| Leukocyte cell-derived chemotaxin 2 precursor a (lect2a) c | F:CAGATGGGGACAAGGACACT | 94.6 | 150 | BT059281 |
| R:GCCTTCTTCGGGTCTGTGTA | - | - | - | |
| Leukocyte cell-derived chemotaxin 2 precursor b (lect2b) c | F:ACAACTGGGGACAAGGACAG | 84.8 | 125 | DV106130 |
| R:CACTTTGCCGTTGAGTTTCA | - | - | - | |
| 60S ribosomal protein L18 (rpl18) | F:AGTTCCACGACTCGAAGATC | 93.8 | 143 | DW535031 |
| R:TTTTATTGTGCCGCACAAGGT | - | - | - | |
| Myocyte-specific enhancer factor 2D (mef2d) | F:GCAGCAACATCAACAACAGC | 89.5 | 160 | XM014177143 |
| R:CTCATCTCTACCCAAGAGGA | - | - | - | |
| Helicase with zinc finger domain 2 a (helz2a, alias pric285a) e | F:GCAAGGTTGGGTATGAGGAA | 91.3 | 149 | BT072427 |
| R:TTCGGAGTTGCTCCAGTCTT | - | - | - | |
| Helicase with zinc finger domain 2 b (helz2b, alias pric285b) e | F:AGACGTAGTGGTTCGGATCG | 82.0 | 145 | EG928625 |
| R:GACCGTGATTTCGTCCAGTT | - | - | - | |
| Integrin beta-5-like (itgb5) f | F:CCTGCCAGCGGCTATGCAA | 94.1 | 147 | DW540995/ XM014165323 |
| R:AGGACTGACATGCCGTTGG | - | - | ||
| Elongation factor 1 alpha-2 (eef1α-2) g | F:GCACAGTAACACCGAAACGA | 86.4 | 132 | BG933853 |
| R:ATGCCTCCGCACTTGTAGAT | - | - | - | |
| 60S ribosomal protein 32 (rpl32) g | F:AGGCGGTTTAAGGGTCAGAT | 96.1 | 119 | BT043656 |
| R:TCGAGCTCCTTGATGTTGTG | - | - | - |
| Probe ID a | BLASTx Identification b | Gene Ontology (GO) of Putative Human Orthologues d | Fold-Change e | |||
|---|---|---|---|---|---|---|
| Best Named BLASTx Hit (Species) c | Accession No. | E-Value | % ID (AA) | |||
| C187R103 | Lipoma HMGIC fusion partner-like 4 protein (lhpl4) (Salmo salar) | NP_001140142 | 0 | 272/272 (100%) | BP: regulation of inhibitory synapse assembly, gamma-aminobutyric acid receptor clustering. MF: protein binding, GABA receptor binding. CC: inhibitory synapse, postsynaptic membrane, cell projection, plasma membrane, cell junction. | 4.78 |
| C231R170 | Serine protease HTRA1 (htra1b) (Salvelinus alpinus) | XP_023864611 | 4e−171 | 248/256 (97%) | BP: proteolysis, extracellular matrix disassembly, negative regulation of transforming growth factor beta receptor signaling pathway, negative regulation of defense response to virus, positive regulation of epithelial cell proliferation. MF: serine-type endopeptidase and peptidase activity, insulin-like growth factor binding, hydrolase activity. CC: collagen-containing extracellular matrix, extracellular space, plasma membrane, cytoplasm. | 3.57 |
| C103R052 | Aminoacyl tRNA synthase complex-interacting multifunctional protein 1 (p43) (Salmo trutta) | XP_029622221 | 0 | 321/326 (98%) | BP: inflammatory response, apoptotic process, response to wounding, tRNA aminoacylation for protein translation, defense response to virus, leukocyte migration, angiogenesis, chemotaxis, positive regulation of glucagon secretion. MF: RNA binding, tRNA binding, protein binding, cytokine activity, protein homodimerization activity. CC: aminoacyl-tRNA synthetase multienzyme complex, nucleus, cytosol, endoplasmic reticulum, extracellular region. | −2.79 |
| C067R040 | Eukaryotic translation initiation factor 2A (eif2a) (Salmo salar) | NP_001133560 | 0 | 576/576 (100%) | BP: translational initiation, ribosome assembly, protein phosphorylation, SREBP signaling pathway, response to amino acid starvation. MF: translation initiation factor activity, cadherin binding, ribosome binding, tRNA binding, protein binding. CC: blood microparticle, extracellular space, cytosolic small ribosomal subunit. | −3.13 |
| C253R093 | Eukaryotic translation initiation factor 4B (eif4b1) (Salvelinus alpinus) | XP_023852969 | 6e−11 | 37/40 (93%) | BP: translational initiation, eukaryotic translation initiation factor 4F complex assembly. MF: RNA binding, protein binding, translation initiation factor activity, RNA strand annealing activity. CC: polysome, cytosol, eukaryotic translation initiation factor 4F complex. | −3.23 |
| C060R108 | Cytochrome c oxidase subunit 2 (mtco2) (Oncorhynchus masou masou) | ASB29545 | 7e−74 | 115/182 (63%) | BP: electron transport chain, oxidation-reduction process. MF: cytochrome-c oxidase activity, copper ion binding, metal ion binding, oxidoreductase activity. CC: membrane, respirasome, mitochondrion. | −3.27 |
| C159R112 | Leukocyte cell-derived chemotaxin 2 precursor (lect2a) (Salmo salar) | ACI67916 | 6e−102 | 155/156 (99%) | BP: chemotaxis, skeletal system development. MF: protein binding, metal ion binding. CC: cytoplasm, extracellular space. | −3.48 |
| C152R057 | 60S ribosomal protein L18 (rpl18) (Salmo trutta) | XP_029599741 | 3e−122 | 172/173 (99%) | BP: translation, viral transcription, SRP-dependent cotranslational protein targeting to membrane. MF: structural constituent of ribosome, protein binding, RNA binding. CC: ribosome, cytosolic large ribosomal subunit, cytosol. | −4.37 |
| C133R018 | Myocyte-specific enhancer factor 2D (mef2d) (Oncorhynchus mykiss) | XP_021427816 | 3e−70 | 193/193 (100%) | BP: positive regulation of vascular smooth muscle cell proliferation, muscle organ development, skeletal muscle and neuronal cell differentiation, apoptotic process, positive regulation of transcription by RNA polymerase II, adult heart development, nervous system development. MF: DNA-binding transcription factor activity, RNA polymerase II-specific, protein binding, histone deacetylase binding, protein heterodimerization activity. CC: nucleus, nuclear chromatin, intracellular membrane-bounded organelle, nucleoplasm. | −4.54 |
| C065R088 | Helicase with zinc finger domain 2 (helz2a alias, pric285a) (Salmo trutta) | XP_029548942 | 0 | 694/714 (97%) | BP: regulation of lipid metabolic process, positive regulation of transcription by RNA polymerase II, nuclear-transcribed mRNA catabolic process, nonsense-mediated decay. MF: nuclear receptor transcription activity, helicase activity, ribonuclease activity, hydrolase activity, RNA binding, ATP binding, protein binding, metal ion binding. CC: nucleus, membrane, nucleoplasm. | −4.71 |
| C002R106 * | Integrin beta-5-like (itgb5) (Oncorhynchus mykiss) | XP_021453113 | 0 | 283/315 (90%) | BP: cell adhesion mediated by integrin, integrin-mediated signaling pathway, muscle contraction, antigen processing and presentation of exogenous peptide antigen via MHC class I, TAP-dependent, viral process, transforming growth factor beta receptor signaling pathway. MF: protein binding, signaling receptor activity, virus receptor activity. CC: cell surface, extracellular exosome, phagocytic vesicle, plasma membrane, integrin complex. | −5.12 |
| C188R069 | Cytochrome c oxidase subunit 1 (mtco1) (Oncorhynchus tshawytscha) * | NP_148940 | 0 | 410/437 (94%) | BP: oxidation-reduction process, oxidative phosphorylation, electron transport chain, aerobic respiration. MF: oxidoreductase activity, cytochrome-c oxidase activity, heme binding, metal ion binding. CC: mitochondrial inner membrane, respiratory chain complex IV, respirasome. | −7.11 |
| Microarray Probe a | Transcript Name | qPCR RQ Values b | p-Value (qPCR) c | Fold-Change d | |||
|---|---|---|---|---|---|---|---|
| High ω3 | Balanced | High ω6 | Microarray | qPCR | |||
| N/A | htra1a | 2.2 ± 0.41 | 1.9 ± 0.29 | 3.0 ± 0.65 | 0.25 | N/A | 1.34 |
| C231R170 | htra1b | 6.0 ± 2.14 | 10.7 ± 3.56 | 22.4 ± 7.43 | 0.07 | 3.57 | 3.75 |
| C103R052 | p43 | 3.4 ± 0.66 | 2.9 ± 0.41 | 2.2 ± 0.47 | 0.24 | −2.79 | −1.59 |
| C067R040 | eif2a | 5.2 ± 0.40 | 5.3 ± 0.74 | 3.5 ± 0.92 | 0.19 | −3.13 | −1.47 |
| C253R093 | eif4b1 | 8.8 ± 1.78 | 6.7 ± 1.43 | 5.5 ± 1.53 | 0.29 | −3.23 | −1.59 |
| N/A | eif4b2 | 2.7 ± 0.30 | 2.7 ± 0.40 | 2.2 ± 0.30 | 0.55 | N/A | −1.22 |
| C060R108 | mtco2 | 1.4 ± 0.10 | 1.2 ± 0.06 | 1.4 ± 0.14 | 0.43 | −3.27 | 1.07 |
| C159R112 | lect2a | 7.6 ± 2.58 | 4.0 ± 1.20 | 4.2 ± 0.96 | 0.38 | −3.48 | −1.79 |
| N/A | lect2b | 3.4 ± 0.74 | 3.7 ± 0.89 | 3.8 ± 0.61 | 0.96 | N/A | 1.12 |
| C152R057 | rpl18 | 2.0 ± 0.17 | 2.1 ± 0.25 | 2.2 ± 0.16 | 0.88 | −4.37 | 1.08 |
| C133R018 | mef2d | 1.9 ± 0.10 a,b | 2.0 ± 0.11 a | 1.5 ± 0.13 b | 0.03 | −4.54 | −1.22 |
| C065R088 | helz2a | 2.3 ± 0.32 a | 1.6 ± 0.15 a,b | 1.5 ± 0.14 b | 0.04 | −4.71 | −1.49 |
| N/A | helz2b | 2.3 ± 0.21 a | 1.4 ± 0.09 b | 1.4 ± 0.11 b | 0.002 | N/A | −1.61 |
| C002R106 | itgb5 | 2.1 ± 0.23 | 1.9 ± 0.08 | 1.6 ± 0.08 | 0.13 | −5.25 | −1.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katan, T.; Xue, X.; Caballero-Solares, A.; Taylor, R.G.; Parrish, C.C.; Rise, M.L. Influence of Varying Dietary ω6 to ω3 Fatty Acid Ratios on the Hepatic Transcriptome, and Association with Phenotypic Traits (Growth, Somatic Indices, and Tissue Lipid Composition), in Atlantic Salmon (Salmo salar). Biology 2021, 10, 578. https://doi.org/10.3390/biology10070578
Katan T, Xue X, Caballero-Solares A, Taylor RG, Parrish CC, Rise ML. Influence of Varying Dietary ω6 to ω3 Fatty Acid Ratios on the Hepatic Transcriptome, and Association with Phenotypic Traits (Growth, Somatic Indices, and Tissue Lipid Composition), in Atlantic Salmon (Salmo salar). Biology. 2021; 10(7):578. https://doi.org/10.3390/biology10070578
Chicago/Turabian StyleKatan, Tomer, Xi Xue, Albert Caballero-Solares, Richard G. Taylor, Christopher C. Parrish, and Matthew L. Rise. 2021. "Influence of Varying Dietary ω6 to ω3 Fatty Acid Ratios on the Hepatic Transcriptome, and Association with Phenotypic Traits (Growth, Somatic Indices, and Tissue Lipid Composition), in Atlantic Salmon (Salmo salar)" Biology 10, no. 7: 578. https://doi.org/10.3390/biology10070578