
Bone Diagenesis in Short Timescales: Insights from an Exploratory Proteomic Analysis

 ,
,
Abstract
:Simple Summary
Abstract
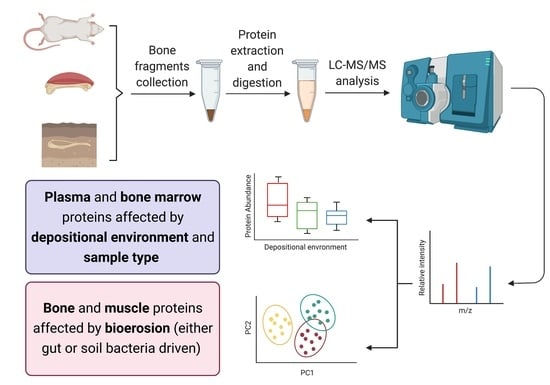
1. Introduction
2. Materials and Methods
2.1. Field Experiment and Sample Composition
2.2. Protein Extraction
2.3. LC–MS/MS Analysis
2.4. Protein Identification and Statistical Analysis
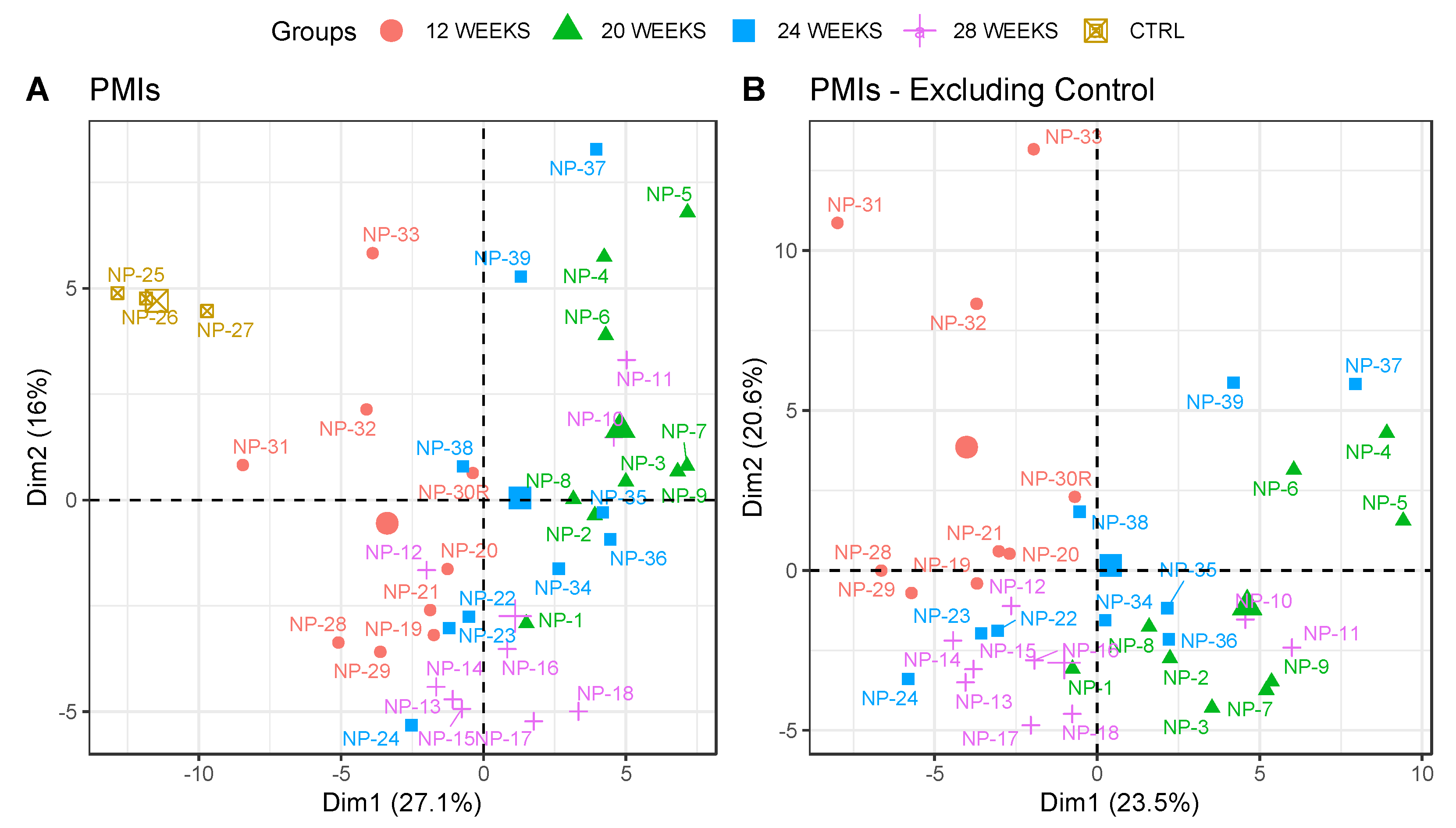
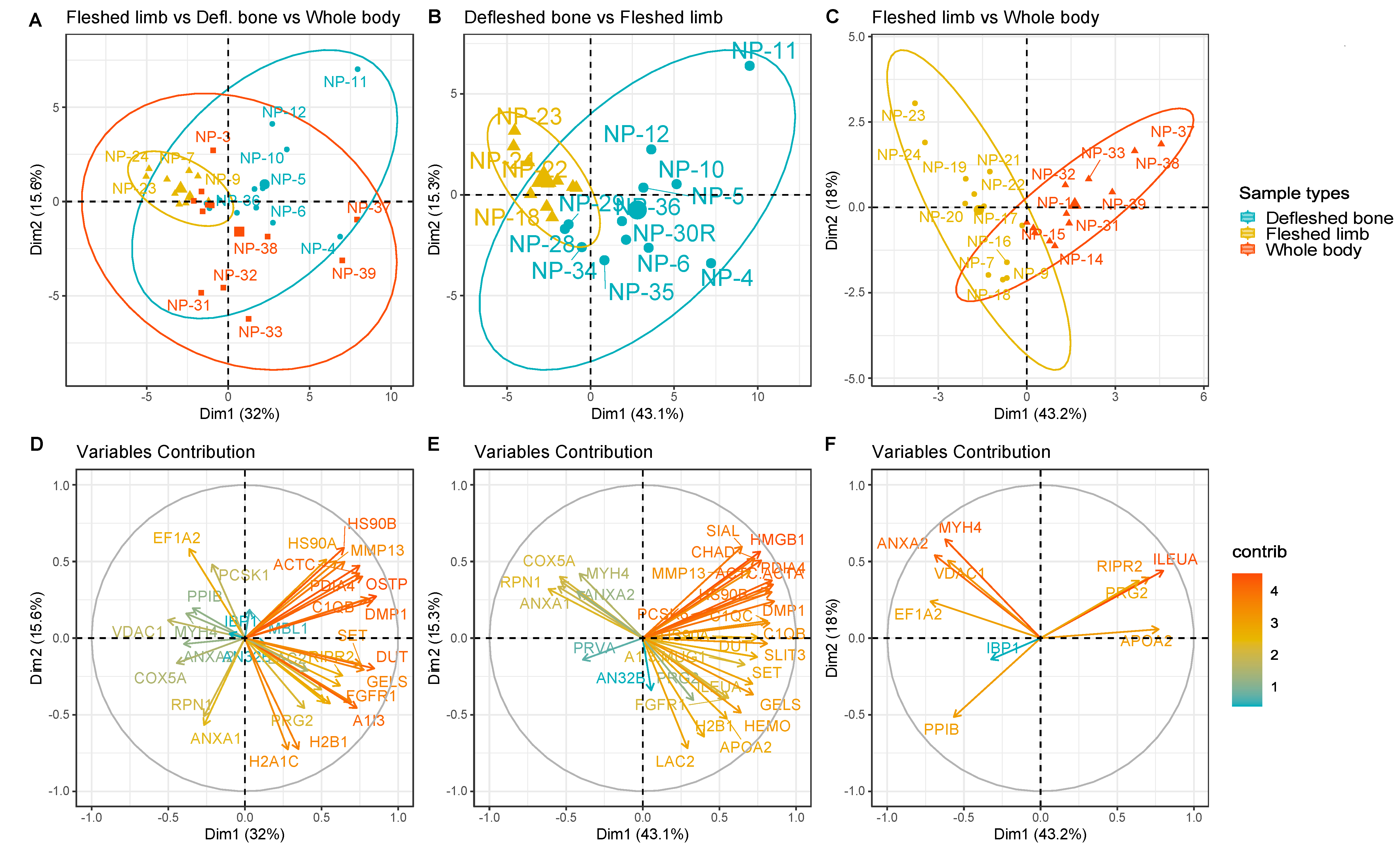
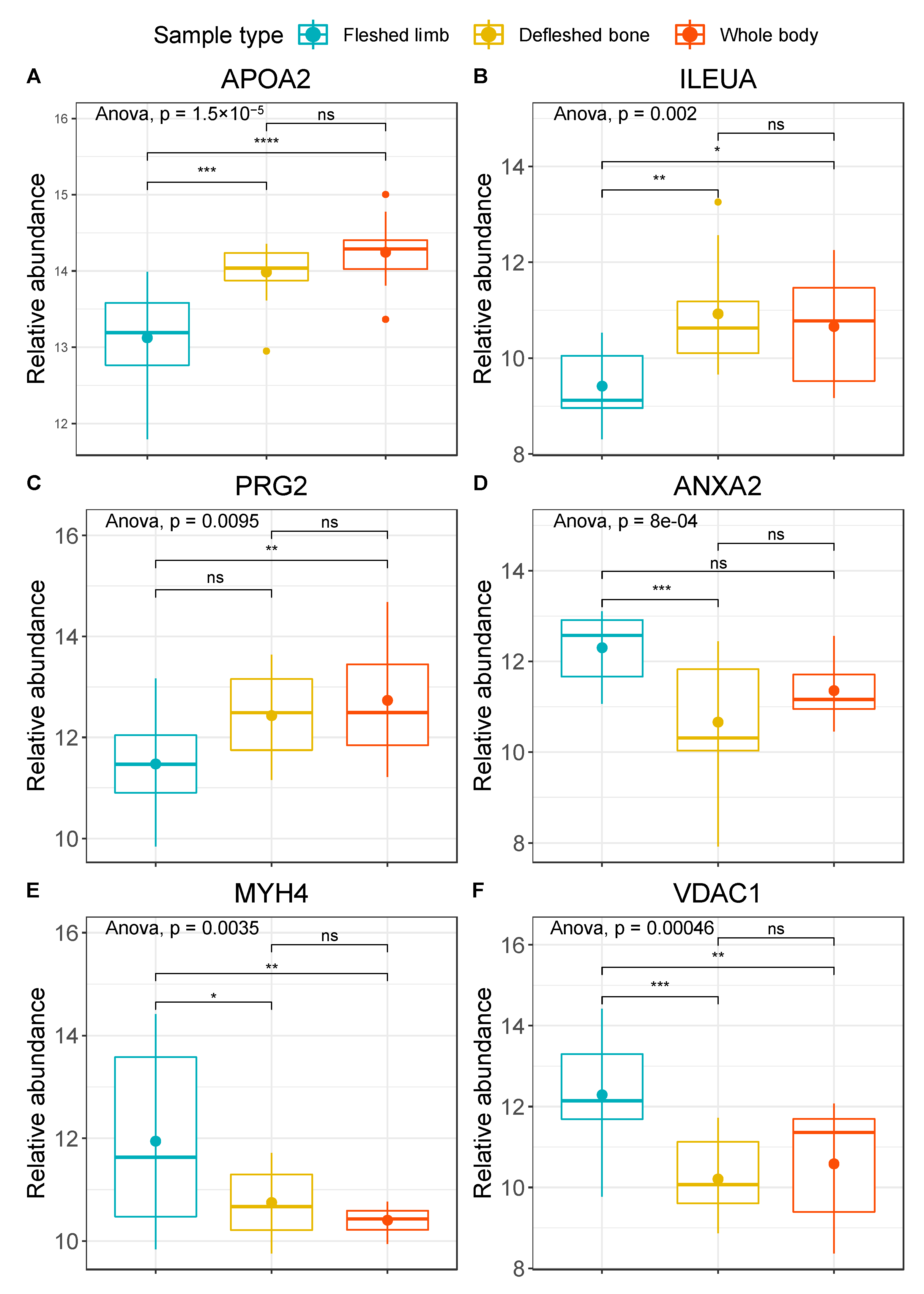
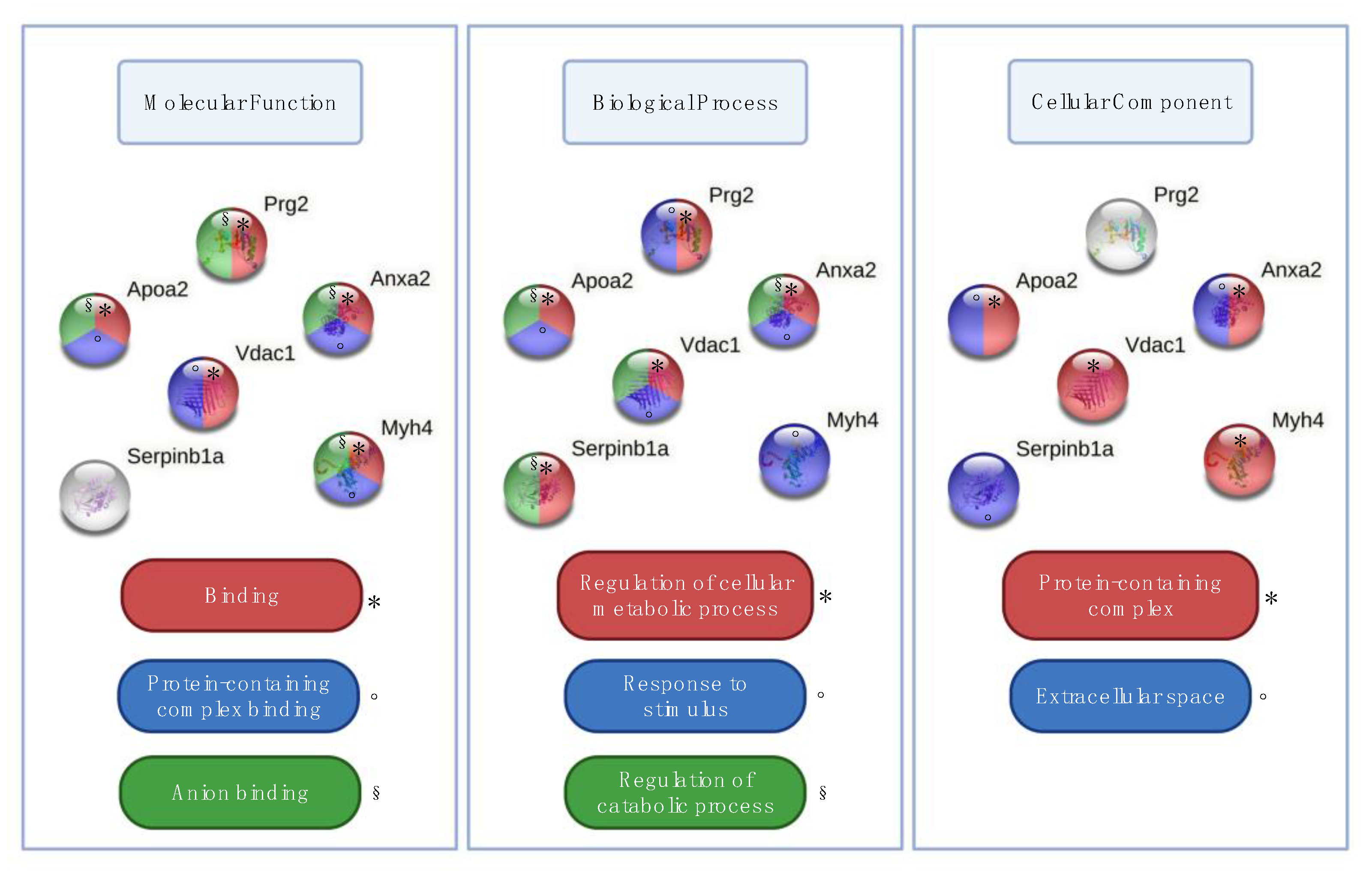
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, L.; Booth, T.J. The origin of bacteria responsible for bioerosion to the internal bone microstructure: Results from experimentally-deposited pig carcasses. Forensic Sci. Int. 2014, 239, 92–102. [Google Scholar] [CrossRef]
- Collins, C.J.; Kozyrev, M.; Frank, M.; Andriotis, O.G.; Byrne, R.A.; Kiener, H.P.; Pretterklieber, M.L.; Thurner, P.J. The impact of age, mineralization, and collagen orientation on the mechanics of individual osteons from human femurs. Materialia 2020, 9, 100573. [Google Scholar] [CrossRef]
- Jans, M.M.E.; Kars, H.; Nielsen-Marsh, C.M.; Smith, C.I.; Nord, A.G.; Arthur, P.; Earl, N. In situ preservation of archaeological bone: A histological study within a multidisciplinary approach. Archaeometry 2002, 44, 343–352. [Google Scholar] [CrossRef]
- Trueman, C.N.; Martill, D.M. The long-term survival of bone: The role of bioerosion. Archaeometry 2002, 44, 371–382. [Google Scholar] [CrossRef]
- Turner-Walker, G. Light at the end of the tunnels? The origins of microbial bioerosion in mineralised collagen. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 529, 24–38. [Google Scholar] [CrossRef]
- Jans, M.; Nielsen-Marsh, C.; Smith, C.; Collins, M.; Kars, H. Characterisation of microbial attack on archaeological bone. J. Archaeol. Sci. 2004, 31, 87–95. [Google Scholar] [CrossRef]
- Brown, S.; Higham, T.; Slon, V.; Pääbo, S.; Meyer, M.; Douka, K.; Brock, F.; Comeskey, D.; Procopio, N.; Shunkov, M.; et al. Identification of a new hominin bone from Denisova Cave, Siberia using collagen fingerprinting and mitochondrial DNA analysis. Sci. Rep. 2016, 6, 23559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.; Kircher, M.; Gansauge, M.-T.; Li, H.; Racimo, F.; Mallick, S.; Schraiber, J.G.; Jay, F.; Prüfer, K.; De Filippo, C.; et al. A High-Coverage Genome Sequence from an Archaic Denisovan Individual. Science 2012, 338, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Buckley, M.; Anderung, C.; Penkman, K.; Raney, B.J.; Götherström, A.; Thomas-Oates, J.; Collins, M.J. Comparing the survival of osteocalcin and mtDNA in archaeological bone from four European sites. J. Archaeol. Sci. 2008, 35, 1756–1764. [Google Scholar] [CrossRef]
- Buckley, M. Ancient collagen reveals evolutionary history of the endemic South American ‘ungulates’. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142671. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Bonete, G.; Pérez-Cárceles, M.D.; Maurandi-López, A.; Pérez-Martínez, C.; Luna, A. Association between protein profile and postmortem interval in human bone remains. J. Proteom. 2019, 192, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martínez, C.; Pérez-Cárceles, M.D.; Legaz, I.; Prieto-Bonete, G.; Luna, A. Quantification of nitrogenous bases, DNA and Collagen type I for the estimation of the postmortem interval in bone remains. Forensic Sci. Int. 2017, 281, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Procopio, N.; Williams, A.; Chamberlain, A.T.; Buckley, M. Forensic proteomics for the evaluation of the post-mortem decay in bones. J. Proteom. 2018, 177, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizukami, H.; Hathway, B.; Procopio, N. Aquatic Decomposition of Mammalian Corpses: A Forensic Proteomic Approach. J. Proteome Res. 2020, 19, 2122–2135. [Google Scholar] [CrossRef] [PubMed]
- Nielsen-Marsh, C. Biomolecules in fossil remains: Multidisciplinary approach to endurance. Biochemist 2002, 24, 12–14. [Google Scholar] [CrossRef] [Green Version]
- Nielsen-Marsh, C.M.; Hedges, R.E.; Mann, T.; Collins, M.J. A preliminary investigation of the application of differential scanning calorimetry to the study of collagen degradation in archaeological bone. Thermochim. Acta 2000, 365, 129–139. [Google Scholar] [CrossRef]
- Buckley, M.; Wadsworth, C. Proteome degradation in ancient bone: Diagenesis and phylogenetic potential. Palaeogeogr. Palaeoclim. Palaeoecol. 2014, 416, 69–79. [Google Scholar] [CrossRef]
- Sawafuji, R.; Cappellini, E.; Nagaoka, T.; Fotakis, A.K.; Jersie-Christensen, R.R.; Olsen, J.V.; Hirata, K.; Ueda, S. Proteomic profiling of archaeological human bone. R. Soc. Open Sci. 2017, 4, 161004. [Google Scholar] [CrossRef] [Green Version]
- Wadsworth, C.; Procopio, N.; Anderung, C.; Carretero, J.-M.; Iriarte, E.; Valdiosera, C.; Elburg, R.; Penkman, K.; Buckley, M. Comparing ancient DNA survival and proteome content in 69 archaeological cattle tooth and bone samples from multiple European sites. J. Proteom. 2017, 158, 1–8. [Google Scholar] [CrossRef]
- Collins, M.J.; Riley, M.S.; Child, A.M.; Turner-Walker, G. A Basic Mathematical Simulation of the Chemical Degradation of Ancient Collagen. J. Archaeol. Sci. 1995, 22, 175–183. [Google Scholar] [CrossRef]
- Hedges, R.E.M. Bone diagenesis: An overview of processes. Archaeometry 2002, 44, 319–328. [Google Scholar] [CrossRef]
- Jans, M.M.E. Microbial bioerosion of bone—A review. In Current Developments in Bioerosion; Wisshak, M., Tapanila, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 397–413. [Google Scholar] [CrossRef]
- Collins, M.; Nielsen-Marsh, C.M.; Hiller, J.; I Smith, C.; Roberts, J.P.; Prigodich, R.V.; Wess, T.J.; Csapo, J.; Millard, A.R.; Turner-Walker, G. The survival of organic matter in bone: A review. Archaeometry 2002, 44, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Wadsworth, C.; Buckley, M. Proteome degradation in fossils: Investigating the longevity of protein survival in ancient bone. Rapid Commun. Mass Spectrom. 2014, 28, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masters, P.M. Preferential preservation of noncollagenous protein during bone diagenesis: Implications for chronometric and stable isotopic measurements. Geochim. Cosmochim. Acta 1987, 51, 3209–3214. [Google Scholar] [CrossRef]
- Smith, C.; Craig, O.; Prigodich, R.; Nielsen-Marsh, C.; Jans, M.; Vermeer, C.; Collins, M. Diagenesis and survival of osteocalcin in archaeological bone. J. Archaeol. Sci. 2005, 32, 105–113. [Google Scholar] [CrossRef]
- Creamer, J.I.; Buck, A.M. The assaying of haemoglobin using luminol chemiluminescence and its application to the dating of human skeletal remains. Lumin 2009, 24, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Costa, I.; Carvalho, F.; Magalhães, T.; De Pinho, P.G.; Silvestre, R.; Dinis-Oliveira, R.J. Promising blood-derived biomarkers for estimation of the postmortem interval. Toxicol. Res. 2015, 4, 1443–1452. [Google Scholar] [CrossRef] [Green Version]
- Takeichi, S.; Tokunaga, I.; Yoshima, K.; Maeiwa, M.; Bando, Y.; Kominami, E.; Katunuma, N. Mechanism of postmortem autolysis of skeletal muscle. Biochem. Med. 1984, 32, 341–348. [Google Scholar] [CrossRef]
- Lametsch, R.; Karlsson, A.; Rosenvold, K.; Andersen, H.J.; Roepstorff, P.; Bendixen, E. Postmortem Proteome Changes of Porcine Muscle Related to Tenderness. J. Agric. Food Chem. 2003, 51, 6992–6997. [Google Scholar] [CrossRef]
- Carter, D.O. The importance of microbial communities in the estimation of the time since death. In Estimation of the Time since Death: Current Research and Future Trends; Hayman, J., Oxenham, M., Eds.; Academic Press: London, UK, 2020; pp. 109–139. [Google Scholar] [CrossRef]
- Melvin, J.R.; Cronholm, L.S.; Simson, L.R.; Isaacs, A.M. Bacterial transmigration as an indicator of time of death. J. Forensic Sci. 1984, 29, 412–417. [Google Scholar] [CrossRef]
- Kellerma, G.D.; Waterman, N.G.; Scharfenberger, L.F. Demonstration in vitro of postmortem bacterial transmigration. Am. J. Clin. Pathol. 1976, 66, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Hyde, E.R.; Haarmann, D.P.; Lynne, A.M.; Bucheli, S.R.; Petrosino, J.F. The living dead: Bacterial community structure of a cadaver at the onset and end of the bloat stage of decomposition. PLoS ONE 2013, 8, e77733. [Google Scholar] [CrossRef]
- Finley, S.J.; Benbow, M.E.; Javan, G.T. Microbial communities associated with human decomposition and their potential use as postmortem clocks. Int. J. Leg. Med. 2014, 129, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Damann, F.E.; Jans, M.M.E. Microbes, anthropology, and bones. In Forensic Microbiology; Carter, D.O., Tomberlin, J.K., Benbow, M.E., Metcalf, J.L., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; p. 319. [Google Scholar]
- Hyde, E.R.; Haarmann, D.P.; Petrosino, J.F.; Lynne, A.M.; Bucheli, S.R. Initial insights into bacterial succession during human decomposition. Int. J. Leg. Med. 2014, 129, 661–671. [Google Scholar] [CrossRef]
- Metcalf, J.L.; Parfrey, L.W.; Gonzalez, A.; Lauber, C.L.; Knights, D.; Ackermann, G.; Humphrey, G.C.; Gebert, M.J.; Van Treuren, W.; Berg-Lyons, D.; et al. A microbial clock provides an accurate estimate of the postmortem interval in a mouse model system. eLife 2013, 2, e01104. [Google Scholar] [CrossRef]
- Cobaugh, K.L.; Schaeffer, S.M.; DeBruyn, J.M. Functional and structural succession of soil microbial communities below decomposing human cadavers. PLoS ONE 2015, 10, e0130201. [Google Scholar] [CrossRef]
- Damann, F.E.; Williams, D.E.; Layton, A.C. Potential use of bacterial community succession in decaying human bone for estimating postmortem interval. J. Forensic Sci. 2015, 60, 844–850. [Google Scholar] [CrossRef]
- Kontopoulos, I.; Nystrom, P.; White, L. Experimental taphonomy: Post-mortem microstructural modifications in Sus scrofa domesticus bone. Forensic Sci. Int. 2016, 266, 320–328. [Google Scholar] [CrossRef]
- Brönnimann, D.; Portmann, C.; Pichler, S.L.; Booth, T.J.; Röder, B.; Vach, W.; Schibler, J.; Rentzel, P. Contextualising the dead—Combining geoarchaeology and osteo-anthropology in a new multi-focus approach in bone histotaphonomy. J. Archaeol. Sci. 2018, 98, 45–58. [Google Scholar] [CrossRef]
- Reiche, I.; Favre-Quattropani, L.; Vignaud, C.; Bocherens, H.; Charlet, L.; Menu, M. A multi-analytical study of bone diagenesis: The Neolithic site of Bercy (Paris, France). Meas. Sci. Technol. 2003, 14, 1608–1619. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; A Holden, P. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Nielsen-Marsh, C.M.; Hedges, R.E. Patterns of Diagenesis in Bone I: The Effects of Site Environments. J. Archaeol. Sci. 2000, 27, 1139–1150. [Google Scholar] [CrossRef]
- Goff, M.L. Early post-mortem changes and stages of decomposition in exposed cadavers. Exp. Appl. Acarol. 2009, 49, 21–36. [Google Scholar] [CrossRef]
- Langley, N.R.; Tersigni-Tarrant, M.A. (Eds.) Forensic Anthropology. A Comprehensive Introduction; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Fernández-Jalvo, Y.; Andrews, P.; Pesquero, D.; Smith, C.; Marín-Monfort, D.; Sánchez, B.; Geigl, E.-M.; Alonso, A. Early bone diagenesis in temperate environments. Palaeogeogr. Palaeoclim. Palaeoecol. 2010, 288, 62–81. [Google Scholar] [CrossRef]
- Yoshino, M.; Kimijima, T.; Miyasaka, S.; Sato, H.; Seta, S. Microscopical study on estimation of time since death in skeletal remains. Forensic Sci. Int. 1991, 49, 143–158. [Google Scholar] [CrossRef]
- Hyde, E.R.; Metcalf, J.L.; Bucheli, S.R.; Lynne, A.M.; Knight, R. Microbial communities associated with decomposing corpses. In Forensic Microbiology; Wiley Online Books; John Wiley Sons Ltd.: Chichester, UK, 2017; pp. 245–273. [Google Scholar]
- Li, D.; Zhu, Z.; Sun, D.-W. Effects of freezing on cell structure of fresh cellular food materials: A review. Trends Food Sci. Technol. 2018, 75, 46–55. [Google Scholar] [CrossRef]
- Speck, M.L.; Ray, B. Effects of Freezing and Storage on Microorganisms in Frozen Foods: A Review. J. Food Prot. 1977, 40, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Procopio, N.; Ghignone, S.; Williams, A.; Chamberlain, A.; Mello, A.; Buckley, M. Metabarcoding to investigate changes in soil microbial communities within forensic burial contexts. Forensic Sci. Int. Genet. 2019, 39, 73–85. [Google Scholar] [CrossRef]
- Procopio, N.; Buckley, M. Minimizing Laboratory-Induced Decay in Bone Proteomics. J. Proteome Res. 2017, 16, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Huber, W.; Von Heydebreck, A.; Sültmann, H.; Poustka, A.; Vingron, M. Variance stabilization applied to microarray data calibration and to the quantification of differential expression. Bioinformatics 2002, 18, S96–S104. [Google Scholar] [CrossRef]
- Blair, H.C.; Kalyvioti, E.; Papachristou, I.N.; Tourkova, I.L.; A Syggelos, S.; Deligianni, D.; Orkoula, M.G.; Kontoyannis, C.G.; A Karavia, E.; Kypreos, E.K.; et al. Apolipoprotein A-1 regulates osteoblast and lipoblast precursor cells in mice. Lab. Investig. 2016, 96, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Fouret, P.; Du Bois, R.M.; Bernaudin, J.F.; Takahashi, H.; Ferrans, V.J.; Crystal, R.G. Expression of the neutrophil elastase gene during human bone marrow cell differentiation. J. Exp. Med. 1989, 169, 833–845. [Google Scholar] [CrossRef]
- Overgaard, M.T.; Oxvig, C.; Christiansen, M.; Lawrence, J.B.; Conover, C.A.; Gleich, G.J.; Sottrup-Jensen, L.; Haaning, J. Messenger Ribonucleic Acid Levels of Pregnancy-Associated Plasma Protein-A and the Proform of Eosinophil Major Basic Protein: Expression in Human Reproductive and Nonreproductive Tissues1. Biol. Reprod. 1999, 61, 1083–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopwood, B.; Tsykin, A.; Findlay, D.; Fazzalari, N. Gene expression profile of the bone microenvironment in human fragility fracture bone. Bone 2009, 44, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Anflous-Pharayra, K.; Cai, Z.-J.; Craigen, W.J. VDAC1 serves as a mitochondrial binding site for hexokinase in oxidative muscles. Biochim. Biophys. Acta BBA Bioenerg. 2007, 1767, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Niu, Y.; Liu, X.; Yang, F.; Niu, W.; Fu, L. Proteomic Analysis of Skeletal Muscle in Insulin-Resistant Mice: Response to 6-Week Aerobic Exercise. PLoS ONE 2013, 8, e53887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adlam, R.E.; Simmons, T. The Effect of Repeated Physical Disturbance on Soft Tissue Decomposition?Are Taphonomic Studies an Accurate Reflection of Decomposition? J. Forensic Sci. 2007, 52, 1007–1014. [Google Scholar] [CrossRef]
- Genetos, D.C.; Wong, A.; Watari, S.; Yellowley, C.E. Hypoxia increases Annexin A2 expression in osteoblastic cells via VEGF and ERK. Bone 2010, 47, 1013–1019. [Google Scholar] [CrossRef] [Green Version]
- Bárány, M. ATPase Activity of Myosin Correlated with Speed of Muscle Shortening. J. Gen. Physiol. 1967, 50, 197–218. [Google Scholar] [CrossRef] [Green Version]
- Shoshan-Barmatz, V.; Hadad, N.; Feng, W.; Shafir, I.; Orr, I.; Varsanyi, M.; Heilmeyer, L.M. VDAC/porin is present in sarcoplasmic reticulum from skeletal muscle. FEBS Lett. 1996, 386, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Pittner, S.; Ehrenfellner, B.; Monticelli, F.C.; Zissler, A.; Sänger, A.M.; Stoiber, W.; Steinbacher, P. Postmortem muscle protein degradation in humans as a tool for PMI delimitation. Int. J. Leg. Med. 2016, 130, 1547–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Codes | Depositional Environment | Sample Type | Timescale in Weeks | Proteomics Code |
|---|---|---|---|---|
| CTRL A-B-C | Control | Control | 0 | NP25-26-27 |
| w12 BB A-B-C | Buried | Defleshed | 12 | NP28-29-30R |
| w12 EW A-B-C | Exposed | Whole body | 12 | NP31-32-33 |
| w12 EF A-B-C | Exposed | Fleshed limb | 12 | NP19-20-21 |
| w20 BB A-B-C | Buried | Defleshed | 20 | NP4-5-6 |
| w20 EW A-B-C | Exposed | Whole body | 20 | NP1-2-3 |
| w20 EF A-B-C | Exposed | Fleshed limb | 20 | NP7-8-9 |
| w24 BB A-B-C | Buried | Defleshed | 24 | NP34-35-36 |
| w24 EW A-B-C | Exposed | Whole body | 24 | NP37-38-39 |
| w24 EF A-B-C | Exposed | Fleshed limb | 24 | NP22-23-24 |
| w28 BB A-B-C | Buried | Defleshed | 28 | NP10-11-12 |
| w28 EW A-B-C | Exposed | Whole body | 28 | NP13-14-15 |
| w28 EF A-B-C | Exposed | Fleshed limb | 28 | NP16-17-18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Procopio, N.; Mein, C.A.; Starace, S.; Bonicelli, A.; Williams, A. Bone Diagenesis in Short Timescales: Insights from an Exploratory Proteomic Analysis. Biology 2021, 10, 460. https://doi.org/10.3390/biology10060460
Procopio N, Mein CA, Starace S, Bonicelli A, Williams A. Bone Diagenesis in Short Timescales: Insights from an Exploratory Proteomic Analysis. Biology. 2021; 10(6):460. https://doi.org/10.3390/biology10060460
Chicago/Turabian StyleProcopio, Noemi, Caley A. Mein, Sefora Starace, Andrea Bonicelli, and Anna Williams. 2021. "Bone Diagenesis in Short Timescales: Insights from an Exploratory Proteomic Analysis" Biology 10, no. 6: 460. https://doi.org/10.3390/biology10060460