
Soil Fungal Community Composition Correlates with Site-Specific Abiotic Factors, Tree Community Structure, and Forest Age in Regenerating Tropical Rainforests

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Molecular Work
2.3. Bioinformatic Work
2.4. Statistical Analyses
3. Results
3.1. Environmental Differences between Regions and Between Primary and Secondary Forests
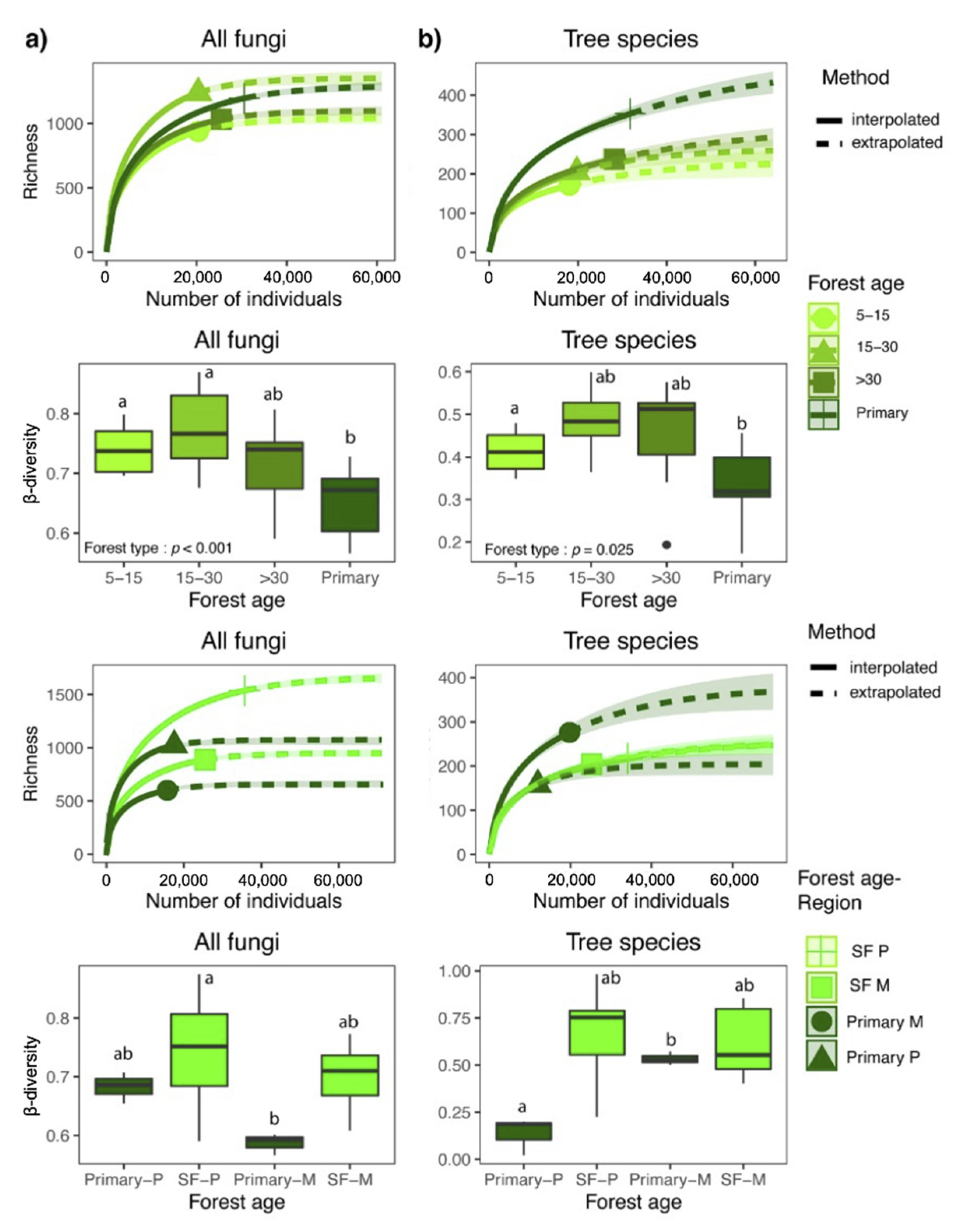
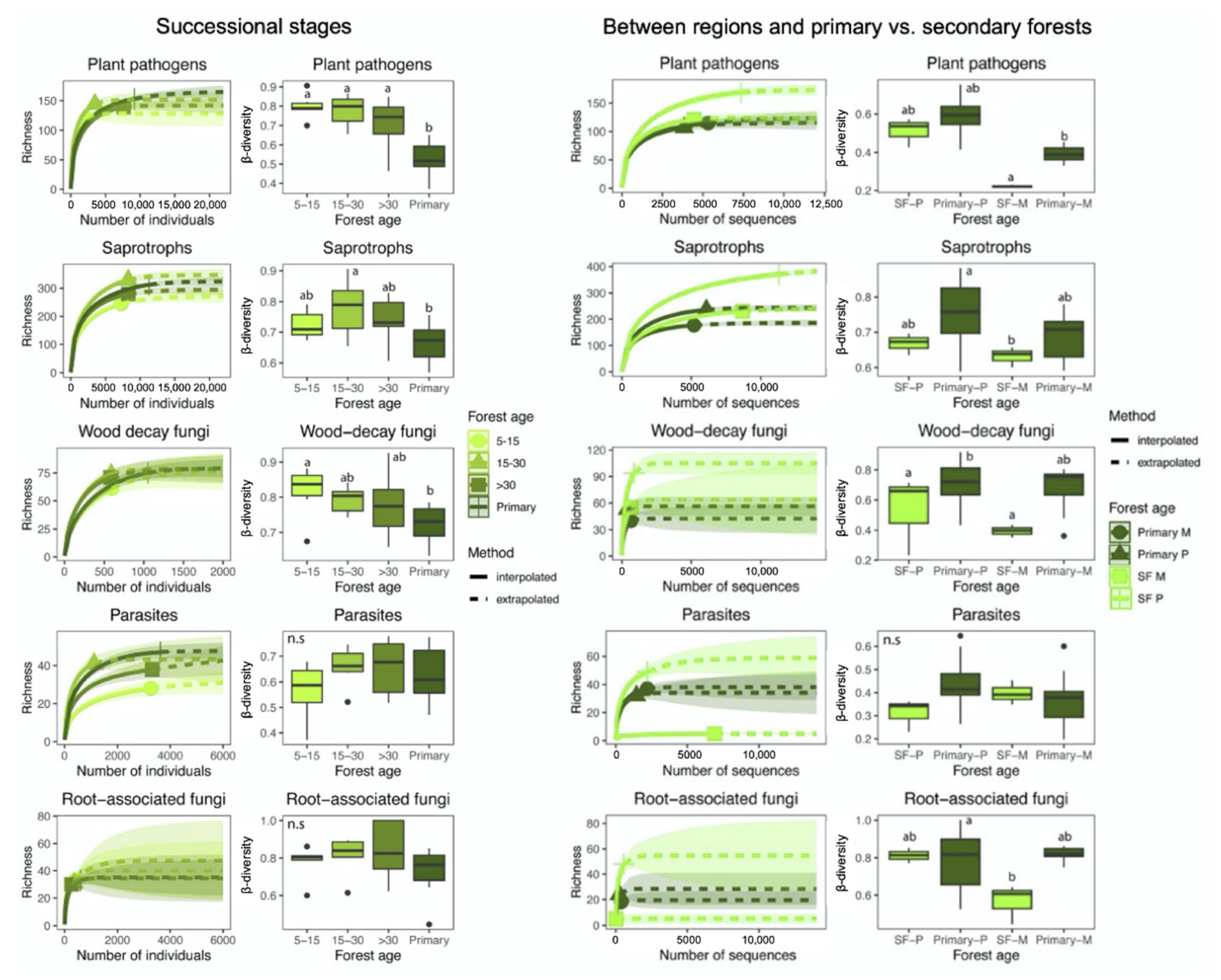
3.2. Patterns of Fungal Richness
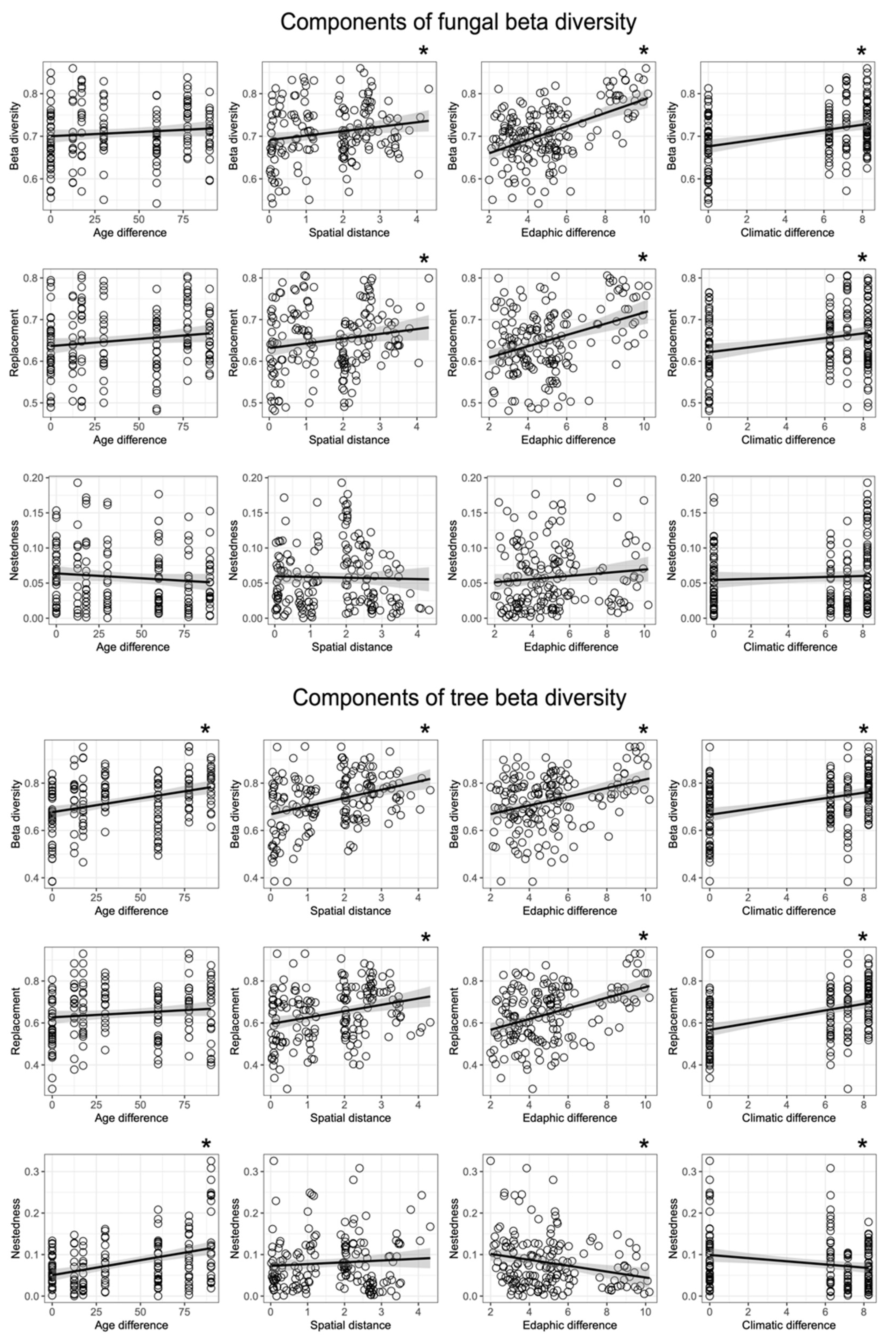
3.3. Community Composition and β-Diversity
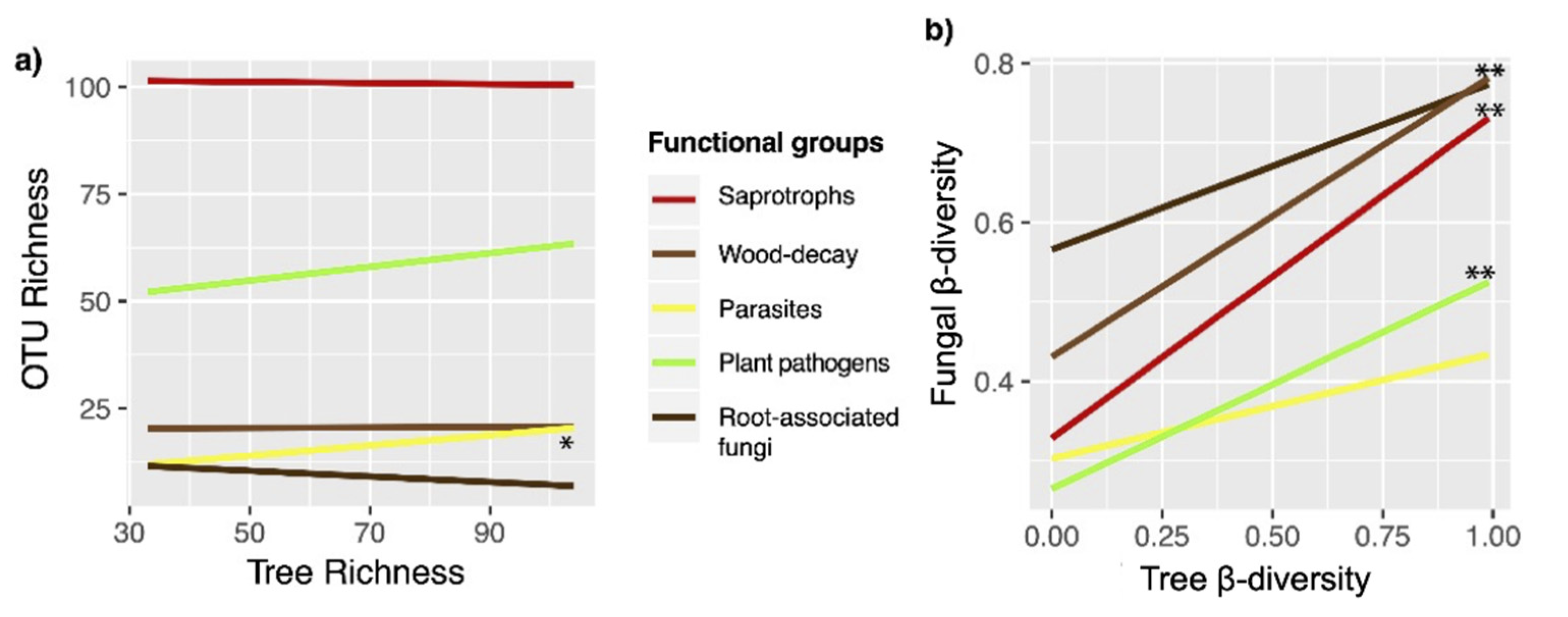
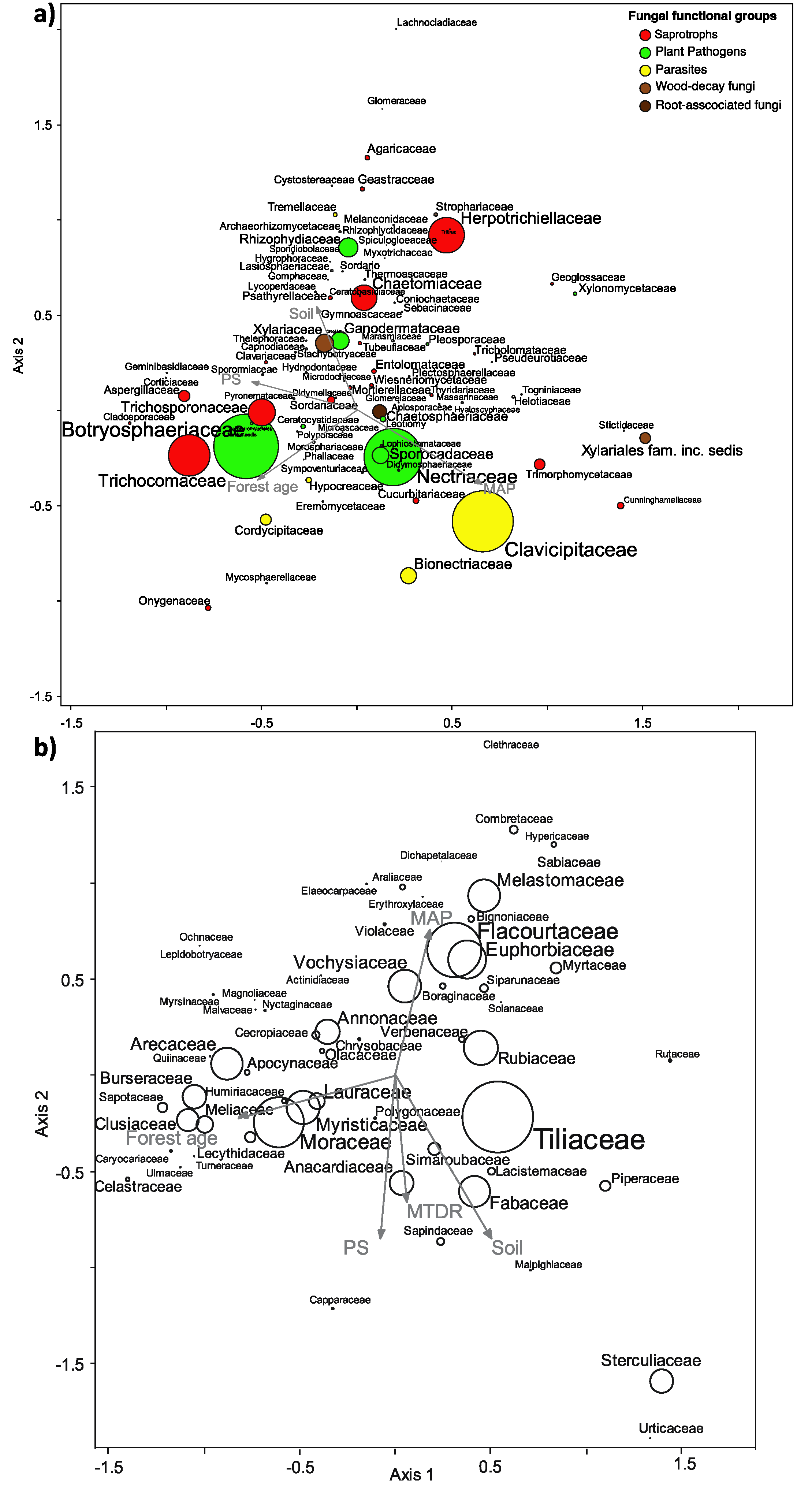
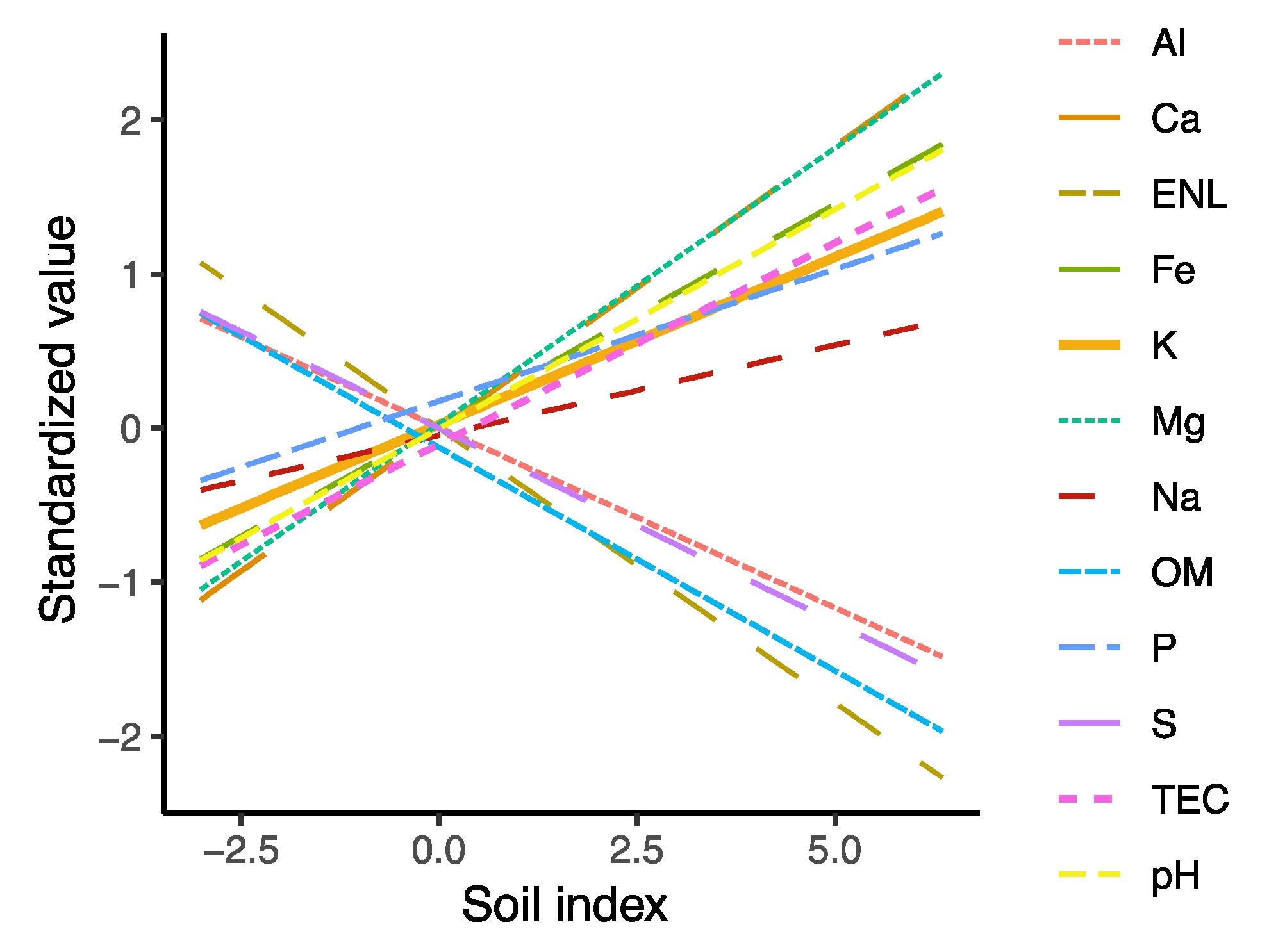
3.4. Effects of Environmental Factors on Fungal Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chazdon, R.L.; Letcher, S.G.; van Breugel, M.; Martinez-Ramos, M.; Bongers, F.; Finegan, B. Rates of change in tree communities of secondary Neotropical forests following major disturbances. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 273–289. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Finegan, B.; Capers, R.S.; Salgado-Negret, B.; Casanoves, F.; Boukili, V.; Norden, N. Composition and dynamics of functional groups of trees during tropical forest succession in northeastern Costa Rica. Biotropica 2010, 42, 31–40. [Google Scholar] [CrossRef]
- Finegan, B. Pattern and process in neotropoical secondary rain forests: The first 100 years of succession. Trends Ecol. Evol. 1996, 11, 119–124. [Google Scholar] [CrossRef]
- Peña-Claros, M. Changes in Forest Structure and Species Composition during Secondary Forest Succession in the Bolivian Amazon. Biotropica 2003, 35, 450–461. [Google Scholar] [CrossRef]
- Dunn, R.R. Recovery of faunal communities during tropical forest regeneration. Conserv. Biol. 2004, 18, 302–309. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Maurenza, D.; Prieto, P.V.; Barros, F.S.M.; Jakovac, C.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Brancalion, P.H.S.; Ceccon, E.; et al. Associations between socio-environmental factors and landscape-level biodiversity recovery in naturally regenerating tropical and subtropical forests. Conserv. Lett. 2021, 14, e12768. [Google Scholar] [CrossRef]
- Dent, D.H.; Joseph Wright, S. The future of tropical species in secondary forests: A quantitative review. Biol. Conserv. 2009, 142, 2833–2843. [Google Scholar] [CrossRef]
- Nascimento, L.M.; de Sá Barretto Sampaio, E.V.; Rodal, M.J.N.; Lins-e-Silva, A.C.B. Secondary succession in a fragmented Atlantic Forest landscape: Evidence of structural and diversity convergence along a chronosequence. J. For. Res. 2014, 19, 501–513. [Google Scholar] [CrossRef]
- Whitfeld, T.J.S.; Lasky, J.R.; Damas, K.; Sosanika, G.; Molem, K.; Montgomery, R.A. Species richness, forest structure, and functional diversity during succession in the New Guinea Lowlands. Biotropica 2014, 46, 538–548. [Google Scholar] [CrossRef]
- Hernández-Ordóñez, O.; Urbina-Cardona, N.; Martínez-Ramos, M. Recovery of amphibian and reptile assemblages during old-field succession of tropical rain forests. Biotropica 2015, 47, 377–388. [Google Scholar] [CrossRef]
- Boukili, V.K.; Chazdon, R.L. Environmental filtering, local site factors and landscape context drive changes in functional trait composition during tropical forest succession. Perspect. Plant Ecol. Evol. Syst. 2017, 24, 37–47. [Google Scholar] [CrossRef]
- Rocha, R.; Ovaskainen, O.; López-Baucells, A.; Farneda, F.Z.; Sampaio, E.M.; Bobrowiec, P.E.D.; Cabeza, M.; Palmeirim, J.M.; Meyer, C.F.J. Secondary forest regeneration benefits old-growth specialist bats in a fragmented tropical landscape. Sci. Rep. 2018, 8, 3819. [Google Scholar] [CrossRef]
- Farneda, F.Z.; Rocha, R.; López-Baucells, A.; Sampaio, E.M.; Palmeirim, J.M.; Bobrowiec, P.E.D.; Grelle, C.E.V.; Meyer, C.F.J. Functional recovery of Amazonian bat assemblages following secondary forest succession. Biol. Conserv. 2018, 218, 192–199. [Google Scholar] [CrossRef]
- Connell, J. Diversity in tropical rain forest and coral reefs: High diversity of trees and corals is maintained only in a nonequilibrium state. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guariguata, M.R.; Ostertag, R. Neotropical secondary forest succession: Changes in structural and functional characteristics. For. Ecol. Manage 2001, 148, 185–206. [Google Scholar] [CrossRef]
- Lasky, J.R.; Uriarte, M.; Boukili, V.K.; Erickson, D.L.; John Kress, W.; Chazdon, R.L. The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession. Ecol. Lett. 2014, 17, 1158–1167. [Google Scholar] [CrossRef]
- Denslow, J.S.; Guzman, G. Variation in stand structure, light and seedling abundance across a tropical moist forest chronosequence in Panama. J. Veg. Sci. 2000, 11, 201–212. [Google Scholar] [CrossRef]
- Bihn, J.H.; Verhaagh, M.; Brändle, M.; Brandl, R. Do secondary forests act as refuges for old growth forest animals? Recovery of ant diversity in the Atlantic forest of Brazil. Biol. Conserv. 2008, 141, 733–743. [Google Scholar] [CrossRef]
- Norden, N.; Angarita, H.A.; Bongers, F.; Martínez-Ramos, M.; Granzow-de la Cerda, I.; van Breugel, M.; Lebrija-Trejos, E.; Meave, J.A.; Vandermeer, J.; Williamson, G.B.; et al. Successional dynamics in Neotropical forests are as uncertain as they are predictable. Proc. Natl. Acad. Sci. USA 2015, 112, 8013–8018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, G.M.; Schmit, J.P. Fungal biodiversity: What do we know? What can we predict? Biodivers. Conserv. 2007, 16, 1–5. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Junninen, K.; Similä, M.; Kouki, J.; Kotiranta, H. Assemblages of wood-inhabiting fungi along the gradients of succession and naturalness in boreal pine-dominated forests in Fennoscandia. Ecography 2006, 29, 75–83. [Google Scholar] [CrossRef]
- Twieg, B.D.; Durall, D.M.; Simard, S.W. Ectomycorrhizal fungal succession in mixed temperate forests. New Phytol. 2007, 176, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Geml, J.; Laursen, G.A.; Herriott, I.; McFarland, J.M.; Booth, M.G.; Lennon, N.; Nusbaum, H.C.; Taylor, D.L. Phylogenetic and ecological analyses of soil and sporocarp DNA sequences reveal high diversity and strong habitat partitioning in the boreal ectomycorrhizal genus Russula Pers. (Russulales; Basidiomycota). New Phytol. 2010, 187, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.L.; Kauserud, H.; Ohlson, M. Forestry impacts on the hidden fungal biodiversity associated with bryophytes. FEMS Microbiol. Ecol. 2014, 90, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Mcguire, K.L.; Fierer, N.; Bateman, C.; Treseder, K.K.; Turner, B.L.; Ecology, S.M.; May, N.; Mcguire, K.L.; Fierer, N.; Bateman, C.; et al. Fungal Community Composition in Neotropical Rain Forests: The Influence of Tree Diversity and Precipitation. Microb. Ecol. 2016, 63, 804–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachelot, B.; Uriarte, M.; Zimmerman, J.; Thompson, J.; Leff, J.W.; Asiaii, A.; Koshner, J.; McGuire, K. Long-lasting effects of land use history on soil fungal communities in second- growth tropical rain forests. Ecol. Appl. 2016, 26, 1881–1895. [Google Scholar] [CrossRef] [Green Version]
- Mueller, R.C.; Rodrigues, J.L.M.; Nüsslein, K.; Bohannan, B.J.M. Land use change in the Amazon rain forest favours generalist fungi. Funct. Ecol. 2016, 30, 1845–1853. [Google Scholar] [CrossRef] [Green Version]
- Janzen, D. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Connell, J.H. On the role of natural enemies in preventing competitive exclusion in some marine mammals and in rain forest trees. In Dynamics of Populations; Boer, P.J., Gradwell, G., Eds.; PUDOC: Wageningen, The Netherlands, 1971; pp. 298–310. [Google Scholar]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- CCT (Centro Científico Tropical, CR); WRI (World Resources Institute, US). La Depreciación de los Recursos Naturales en Costa Rica y su Relación con el Sistema de Cuentas Nacionales [The Depreciation of Natural Resources in Costa Rica and Its Relationship with the System of National Accounts]; WRI: Washington, DC, USA, 1991; 160p. [Google Scholar]
- Vílchez, B.; Chazdon, R. Tipos de Bosque del Corredor Biológico Osa (Entrevista) [Forest Types of the Osa Biological Corridor (Interview)]; Instituto Tecnológico de Costa Rica: Cartago, Costa Rica, 2009. [Google Scholar]
- Morales-Salazar, M.; Vílchez-Alvarado, B.; Ortega-Gutiérrez, M.; Ortiz-Malavasi, E.; Guevara-Bonilla, M.; Chazdon, R.I. Diversidad y estructura horizontal en los bosques tropicales del Corredor Biológico de Osa, Costa Rica [Diversity and horizontal structure of tropical forests of the Osa Biological Corridor, Costa Rica]. Rev. For. Mesoam. Kurú 2012, 9, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.; Asner, G.; Dahlin, K.; Anderson, C.; Knapp, D.; Martin, R.; Mascaro, J.; Chazdon, R.; Cole, R.; Wanek, W.; et al. Landscape-scale controls on aboveground forest carbon stocks on the Osa Peninsula, Costa Rica. PLoS ONE 2015, 10, e0126748. [Google Scholar] [CrossRef] [Green Version]
- Weissenhofer, A.; Huber, W.; Koukal, T.; Immitzer, M.; Schembera, E.; Sontag, S.; Zamora, N.; Weber, A. Ecosystem diversity in the Piedras Blancas National Park and adjacent areas (Costa Rica), with the first vegetation map of the area. Nat. Cult. Hist. Golf. Dulce Reg. Costa Rica 2008, 80, 65–96. [Google Scholar]
- Huber, W.; Weissenhofer, A.; Zamora, N.; Weber, A. Plant diversity and biogeography of the Golfo Dulce region, Costa Rica. Stapfia 2008, 88, 97–104. [Google Scholar]
- Hietz, P. Plant life forms in the Golfo Dulce region and other neotropical rainforests. Stapfia 2008, 88, 129–142. [Google Scholar]
- Cornejo, X.; Mori, S.A.; Aguilar, R.; Stevens, H.; Douwes, F. Phytogeography of the trees of the Osa Peninsula, Costa Rica. Brittonia 2012, 64, 76–101. [Google Scholar] [CrossRef]
- Buchs, D.M.; Baumgartner, P.O.; Baumgartner-Mora, C.; Bandini, A.N.; Jackett, S.-J.; Diserens, M.-O.; Stucki, J. Late Cretaceous to Miocene seamount accretion and mélange formation in the Osa and Burica Peninsulas (Southern Costa Rica): Episodic growth of a convergent margin. Geol. Soc. Lond. Spec. Publ. 2009, 328, 411–456. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, R.; Cornejo, X. Plants endemic to the Osa Peninsula (En Liínea); The New York Botanical Garden: Bronx, NY, USA, 2010. [Google Scholar]
- Morales-Salazar, M. Composición Florística, Estructura, Muestreo Diagnóstico y Estado de Conservación de una Cronosecuencia Bosques Tropicales del Corredor Biológico Osa [Floristic Composition, Structure, Diagnostic Sampling and State of Conservation of a Chronosequence of Tropical Forests in the Osa Biological Corridor, Costa Rica]. Diploma Thesis, Escuela de Ingegnería Forestal, Cartago, Costa Rica, 2010; 115p. [Google Scholar]
- Morales-Salazar, M.; Vílchez-Alvarado, B.; Ortega-Gutiérrez, M.; Ortiz-Malavasi, E.; Guevara-Bonilla, M.; Chazdon, R.I. Estructura, compsición y diversidad vegetal en los bosques tropicales del Corredor Biológico de Osa, Costa Rica [Structure, composition, and diversity of plants in tropical forests in the Osa Biological Corridor, Costa Rica]. Rev. For. Mesoam. Kurú 2013, 10, 1–13. [Google Scholar]
- Vargas-Fonseca, L. Análisis de una Cronosequencia de Bosques Tropicales de Corredor Biológico Osa, Costa Rica [Analyses of a Chronosequence of Tropical Forests in the Osa Biological Corridor, Costa Rica]. Diploma Thesis, Escuela de Ingegnería Forestal, Cartago, Costa Rica, 2012; 99p. [Google Scholar]
- Ihrmark, K.; Bödeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandström-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region—evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Geml, J.; Pastor, N.; Fernandez, L.; Pacheco, S.; Semenova, T.A.; Becerra, A.G.; Wicaksono, C.Y.; Nouhra, E.R. Large-scale fungal diversity assessment in the Andean Yungas forests reveals strong community turnover among forest types along an altitudinal gradient. Mol. Ecol. 2014, 23, 2452–2472. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2014, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “vegan”. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Chazdon, R.L.; Shen, T.J. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Lett. 2005, 8, 148–159. [Google Scholar] [CrossRef]
- Wheeler, B. CRAN—Package lmPerm. R Packag. version. 2010. Available online: https://cran.r-project.org/web/packages/lmPerm/lmPerm.pdf (accessed on 30 September 2021).
- Mccune, B.; Grace, J.B. Analysis of Ecological Communities. MjM Software: Gleneden Beah, OR, USA, 2002; 304p. [Google Scholar]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Wiser, S.K.; Brotons, L. Using species combinations in indicator analyses. Methods Ecol. Evol. 2012, 3, 973–982. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Comita, L.S.; Muller-Landau, H.C.; Aguilar, S.; Hubbell, S.P. Asymmetric Density Dependence Shapes Tropical Tree Species Abundances. Science 2010, 329, 330–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peay, K.G.; Baraloto, C.; Fine, P.V. Strong coupling of plant and fungal community structure across western Amazonian rainforests. ISME J. 2013, 7, 1852–1861. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, R.; Swinfield, T.; Gallery, R.E.; Lewis, O.T.; Gripenberg, S.; Narayan, L.; Freckleton, R.P. Testing the Janzen-Connell mechanism: Pathogens cause overcompensating density dependence in a tropical tree. Ecol. Lett. 2010, 13, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Castro-Luna, A.A.; Sosa, V.J.; Castillo-Campos, G. Bat diversity and abundance associated with the degree of secondary succession in a tropical forest mosaic in south-eastern Mexico. Anim. Conserv. 2007, 10, 219–228. [Google Scholar] [CrossRef]
- Barlow, J.; Gardner, T.A.; Araujo, I.S.; Avila-Pires, T.C.; Bonaldo, A.B.; Costa, J.E.; Esposito, M.C.; Ferreira, L.V.; Hawes, J.; Hernandez, M.I.M.; et al. Quantifying the biodiversity value of tropical primary, secondary, and plantation forests. Proc. Natl. Acad. Sci. USA 2007, 104, 18555–18560. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.L.; Herriott, I.C.; Stone, K.E.; McFarland, J.W.; Booth, M.G.; Leigh, M.B. Structure and resilience of fungal communities in Alaskan boreal forest soils. Can. J. For. Res. 2010, 40, 1288–1301. [Google Scholar] [CrossRef] [Green Version]
- Mueller, R.C.; Paula, F.S.; Mirza, B.S.; Rodrigues, J.L.; Nüsslein, K.; Bohannan, B.J. Links between plant and fungal communities across a deforestation chronosequence in the Amazon rainforest. ISME J. 2014, 8, 1548–1550. [Google Scholar] [CrossRef]
- Chaverri, P.; Vilchez, B. Hypocrealean (Hypocreales, Ascomycota) Fungal Diversity in Different Stages of Tropical Forest Succession in Costa Rica1. Biotropica 2006, 38, 531–543. [Google Scholar] [CrossRef]
- Pinho, B.X.; Pimentel, F.; Melo LDe Tabarelli, M.; Pierce, S.; Lohbeck, M. Soil-mediated filtering organizes tree assemblages in regenerating tropical forests. J. Ecol. 2018, 106, 137–147. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Öpik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, Ü.; Zobel, M. The online database MaarjAM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef]
- Erlandson, S.R.; Savage, J.A.; Cavender-Bares, J.M.; Peay, K.G. Soil moisture and chemistry influence diversity of ectomycorrhizal fungal communities associating with willow along an hydrologic gradient. FEMS Microbiol. Ecol. 2016, 92, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J. Plant diversity in tropical forests: A review of mechanisms of species coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef]
- Gilbert, G.S. Dimensions of Plant Disease in Tropical Forests; Biotic Interactions in the Tropics: Their Role in the Maintenance of Species Diversity; Cambridge University Press: Cambridge, UK, 2005; pp. 141–164. [Google Scholar] [CrossRef]
- Bell, T.; Freckleton, R.P.; Lewis, O.T. Plant pathogens drive density-dependent seedling mortality in a tropical tree. Ecol. Lett. 2006, 9, 569–574. [Google Scholar] [CrossRef] [PubMed]
- FAO. Global Forest Resources Assessment 2015; FAO Forestry: Rome, Italy, 2016. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Community | All Sites | Primary Forest | Secondary Forest | |||
|---|---|---|---|---|---|---|
| Var. (%) | p | Var. (%) | p | Var. (%) | p | |
| Successional stage (cat.) | 26.96 | 0.007 | - | - | - | - |
| Region (cat.) | 14.40 | 0.004 | 25.59 | 0.3 | 19.79 | 0.008 |
| Edaphic index | 15.33a | 0.003 | 17.92 | 0.659 | 21.90 | 0.002 |
| Age | 15.20 | 0.003 | - | - | - | - |
| MAP | 14.40 | 0.005 | 25.91 | 0.217 | 19.82 | 0.005 |
| PS | 13.42 | 0.01 | 32.38 | 0.03 | 18.52 | 0.008 |
| MDRT | 10.5 | 0.003 | 30.39 | 0.05 | 15.11 | 0.056 |
| Tree Community | All Sites | Primary Forest | Secondary Forest | |||
| Var. (%) | p | Var. (%) | p | Var. (%) | p | |
| Successional stage (cat.) | 31.72 | 0.021 | - | - | - | - |
| Region (cat.) | 25.98 | 0.001 | 78.06 | 0.300 | 30.25 | 0.001 |
| Edaphic index | 22.63 | 0.001 | 14.36 | 0.535 | 32.07 | 0.001 |
| Age | 20.85 | 0.001 | - | - | - | - |
| MAP | 26.22 | 0.001 | 78.34 | 0.017 | 31.36 | 0.001 |
| PS | 25.47 | 0.001 | 66.4 | 0.017 | 30.59 | 0.001 |
| MDRT | 12.59 | 0.036 | 23.58 | 0.350 | 18.27 | 0.042 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamo, I.; Ortiz-Malavasi, E.; Chazdon, R.; Chaverri, P.; ter Steege, H.; Geml, J. Soil Fungal Community Composition Correlates with Site-Specific Abiotic Factors, Tree Community Structure, and Forest Age in Regenerating Tropical Rainforests. Biology 2021, 10, 1120. https://doi.org/10.3390/biology10111120
Adamo I, Ortiz-Malavasi E, Chazdon R, Chaverri P, ter Steege H, Geml J. Soil Fungal Community Composition Correlates with Site-Specific Abiotic Factors, Tree Community Structure, and Forest Age in Regenerating Tropical Rainforests. Biology. 2021; 10(11):1120. https://doi.org/10.3390/biology10111120
Chicago/Turabian StyleAdamo, Irene, Edgar Ortiz-Malavasi, Robin Chazdon, Priscila Chaverri, Hans ter Steege, and József Geml. 2021. "Soil Fungal Community Composition Correlates with Site-Specific Abiotic Factors, Tree Community Structure, and Forest Age in Regenerating Tropical Rainforests" Biology 10, no. 11: 1120. https://doi.org/10.3390/biology10111120