
The Curious Case of Earthworms and COVID-19

1
Department of Biosciences, Aarhus University, 8600 Silkeborg, Denmark
2
Department of Biology & CESAM, University of Aveiro, 3810-193 Aveiro, Portugal
*
Author to whom correspondence should be addressed.
Biology 2021, 10(10), 1043; https://doi.org/10.3390/biology10101043
Submission received: 9 September 2021
/
Revised: 8 October 2021
/
Accepted: 12 October 2021
/
Published: 14 October 2021
(This article belongs to the Section Infection Biology)
{kind=link}
Abstract
:Simple Summary
Earthworms have been used for centuries in traditional medicine, and more than a century ago were praised by Charles Darwin as one of the most important organisms in the history of the world. These worms are well-studied with a wealth of information available, for example on the genome, the gene expression, the immune system, the general biology, and ecology. These worms live in many habitats, and they had to find solutions for severe environmental challenges. The common compost worm, Eisenia fetida, has developed a unique mechanism to deal with intruding (nano)materials, bacteria, and viruses. It deals with the intruders by covering these with a defence toxin (lysenin) targeted to kill the intruder. We outline how this mechanism probably can be used as a therapeutic model for human COVID-19 (Severe Acute Respiratory Syndrome Coronavirus 2, SARS-CoV-2) and other corona viruses.
Abstract
Earthworms have been used for centuries in traditional medicine and are used globally as an ecotoxicological standard test species. Studies of the earthworm Eisenia fetida have shown that exposure to nanomaterials activates a primary corona-response, which is covering the nanomaterial with native proteins, the same response as to biological invaders such as a virus. We outline that the earthworm Eisenia fetida is possibly immune to COVID-19 (Severe Acute Respiratory Syndrome Coronavirus 2, SARS-CoV-2), and we describe the likely mechanisms of highly receptor-specific pore-forming proteins (PFPs). A non-toxic version of this protein is available, and we hypothesize that it is possible to use the earthworm’s PFPs based anti-viral mechanism as a therapeutic model for human SARS-CoV-2 and other corona viruses. The proteins can be used as a drug, for example, delivered with a nanoparticle in a similar way to the current COVID-19 vaccines. Obviously, careful consideration should be given to the potential risk of toxicity elicited by lysenin for in vivo usage. We aim to share this view to activate its exploration by the wider scientific community while promoting a potential therapeutic development.
1. Introduction
Earthworms have been used for centuries in traditional medicine [1], and more than a century ago were praised by Darwin [2] as one of the most important organisms in the history of the world. They are widely distributed in many habits, occurring in virtually all soils where the moisture and organic content is adequate, and have tremendous importance for general soil function, and for the growth of agricultural crops [3].
Earthworms such as Eisenia fetida are probably the oldest standardised soil model-organisms in ecotoxicology, i.e., used to assess environmental contamination [4], and they are being tested globally [5,6,7,8]. The knowledge developed within these ecotoxicological test on earthworms is vast, not only regarding the standard information required for chemical substances tested under REACH (Registration, Evaluation, Authorisation of Chemicals) frameworks, but also regarding a wide diversity of additional biological endpoints. For these worms, assays can be conducted both using in vitro (cells) and in vivo (whole organisms) systems [7,9], and the endpoints cover information on, e.g., the genome, all omics (transcriptomics, metabolomics, proteomics), the innate immune system, and all of the organism’s life cycle stages (cocoons, juveniles, adults) [10,11]. Hence, there is an excellent understanding of the physiology of these organisms.
The various earthworm species live in a wide diversity of habitats and have had to find solutions—adaptations—for severe environmental challenges. The species Eisenia fetida has developed a unique mechanism to deal with intruding (nano)materials, bacteria, and viruses.
In our studies on the potential hazards of nanomaterials [10,12,13], searching for the mechanism of how nanomaterials affect organisms, we have found that nanomaterials become covered by protein coronas when exposed to biological media or in an organism. For Eisenia fetida, the dominant protein in the corona is lysenin, which is rapidly induced and highly regulated. Lysenin is likely to have evolved in response to biological invaders, e.g., a virus [14,15]. Hence, the worms treat nanoparticle intruders as it treats biological intruders.
2. Earthworm and COVID-19
This led us to wonder if the humble earthworm could have a mechanism that counters infection by enveloped viruses, such as SARS-CoV-2 (Severe Acute Respiratory Syndrome Coronavirus 2, COVID-19), Severe Acute Respiratory Syndrome (SARS), and Middle East Respiratory Syndrome (MERS).
COVID-19 research covers a wide range of specific internal infection (and infection countering) mechanisms, and/or corona viral entry points [16,17,18,19,20,21,22,23]. However, it has long been realized that a weak point of enveloped viruses may be the lipid layer, e.g., of the SARS-CoV-2 envelope [24]. The targeting of viral lipids or host lipids required for viral assembly may represent an effective virucidal approach to develop new anti-viral drugs [25,26,27]. Such lipids, which include sphingolipids, may also interfere with SARS-CoV-2 by binding to ACE-2 (Angiotensin-Converting Enzyme 2) [25,28].
Eisenia fetida has a well-known extracellular target defence mechanism involving pore-forming proteins (PFPs) that target lipids [14,15]. PFPs are produced by chloragogen and coelomic cells as soluble proteins that migrate toward the target membrane where they bind with high affinity. These proteins are a group of closely related proteins: lysenin (41 Kda), fetidin (40/45 Kda), and eiseniapore (38 Kda), which respond immediately to “foreign material” invading the worms [10,12,13,14,15,29]. Lysenin and fetidin bind strongly to sphingomyelin (SM, a sphingolipid), destabilize the cell membranes, and cause lysis [30,31]. The process consists of PFPs, e.g., lysenin, forming three (inner)–10 (outer) nm wide pores in the invading “materials/organisms’” membrane [32,33,34]. Sphingomyelin is present in most cell membranes, including the SARS-CoV-2 envelope, in which it may have a higher abundance [35,36,37,38]. In particular, lysenin and fetidin target SM, whereas the eiseniapore is not solely dependent on SM but also binds to galactosylceramide [32,39]. Although the mechanism and structure are well-understood, the exact biological function of lysenin and fetidin is not fully understood, since invading bacteria or fungi rarely have SM in the membrane [40]; however, invading eukaryotes and many enveloped viruses will contain SM. Hence, lysenin (and fetidin) are excellent for defending against enveloped viruses due to their SM specificity and should be further explored.
The virus acts by hijacking the target cells’ metabolic system, altering their lipid metabolism so that it produces the lipid rafts needed by the virus for formation and for replication, which, in some cases, contain substantial enrichment of the sphingolipids [37,38,39,41,42,43,44,45,46]. There are indications that both cellular and viral SM is required for virus infections of Influenza type A virus [35], for HIV [42], for West Nile virus [46], and SARS-CoV [47,48,49]. Moreover, SM depletion in influenza A viruses’ envelopes reduces their ability to infect cells [50], and crude extracts of E. fetida tissues can inhibit their cytotoxicity by 69–100% [51]. Hence, worms are unlikely to be infected by SARS-CoV-2 or other envelope (corona) embedded virus, and may be better at handling other viruses, such as Ebola virus and HIV, which require SM and sphingomyelinase activity for infectivity [52].
The earthworm avoids internal damage (self-destruction) by the PFPs (e.g., lysenin) because they are virtually absent of SM [53,54]. Sadly, of course, we humans are not similar to worms and do have SM in our cell membranes, with some areas enriched in SM, e.g., in the myelin layer of nerve axons or exosomes [55]. Hence, to directly inject native E. fetida PFPs, e.g., lysenin, into humans will not provide an effective approach to prevent or treat human viral infections.
3. Exploration
However, explorations of traditional and new potential applications of E. fetida in medicine [1,56] could be extended to include a possible exploitation of earthworms’ virucidal and anti-viral mechanisms to fight infections by SARS-CoV-2 and related corona viruses. For example, by modifying the N-terminal [30,31,34], it is possible to synthesize a modified version of lysenin that is not toxic to human cells (NT-Lysenin) but that still selectively binds to SM clusters (5–6 SM) in membranes [57]. It is also possible to tag lysenin, e.g., with polyhistidine or fluorescence tags [58], and this is often used to identify SM in membranes. Further, it is possible to module the action of the lysenin-induced channels by interactions with multivalent inorganic and organic cations [59].
As mentioned above, the virus membrane differs from that of the host and the virus highjacks the cells’ metabolic system to alter the lipid metabolism so that it produces the lipid rafts needed by the virus for replication [37,38,44,60]. Thus, it may be possible to target this binding process therapeutically (Figure 1) if the SM distribution differs sufficiently between human and viral membranes, as shown for HIV and West Nile virus where the SM level is (e.g., 2–3 times) higher than the target cell [46,61,62,63]. Further, for SARS-CoV (and HIV), the virus binding is both dependent on the lipid rafts (sphingomyelin and cholesterol) and the proteins (e.g., ACE2) in the cell membrane, hence NT-lysenin may be used to study whether there is a reorganisation of SM in the cell surface of the target cell and of the virus [47,48,49,64]. It may be noted that SM plays a particular role in determining the cholesterol accessibility (due to high SM affinity to cholesterol [65]) and that higher cholesterol availability in membrane coincides with SARS-CoV-2 entry [66]. Other alternatives, to ensure that lysenin-derivatives specifically bind to viral membranes, may also emerge if researchers explore their therapeutic potential, e.g., developing SARS-CoV-2-related molecular traps [18]. Since it is possible to tag lysenin, lysenin can also deliver drugs attached to the N-terminal [58,67]. In addition, the delivery of lysenin can be targeted to specific tissues if they are carried with nanocarriers (inside or outside), e.g., gold nanoparticles, lipid- or polyethylene glycol (PEG)-carriers, as used for vaccines [68,69]. The delivery carrier should obviously be chosen for this specific purpose, and it should be considered that lysenin may interact with the nanocarrier itself, e.g., if a sphingomyelin-based liposome is used [70,71]. Hence, various model carriers should be tested, e.g., some of the carriers that are already approved for vaccines. Further, earthworms contain a number of other interesting anti-infection compounds, e.g., lumbricins [72]. Obviously, similar considerations and investigations should be given to the two other earthworm PFPs, fetidin and eiseniapore, perhaps as a combined approach. To support the approach, it is well-known that pore-forming proteins have important biological functions and that these can be therapeutically effective, e.g., against cancer, inflammatory diseases, and immunomodulation [68,73,74,75]. Given this, it seems prudent to take an extra look at this humble earthworm to help us develop new treatments, also in the light of the mutation in the spike protein.
4. Conclusions
More studies should be conducted to explore the kinetic processes of PFPs formation, topological accumulation, and dynamic alterations of the sphingomyelin pool in human airway epithelial cells. Careful consideration must also be given to the potential risk of toxicity elicited by lysenin for in vivo usage. It is worth noticing that there is a good understanding of lysenin’s binding to sphingomyelin clusters in cell membranes, cells producing these proteins are easily extracted from the worms, and that the species (E. fetida) is present in many laboratories across the world, since they have been OECD standard test species for environmental toxicology for more than 25 years. Hence, organizations such as the WHO (World Health Organization) can quickly mobilize many scientists to explore the possibility of using the pore-forming proteins to target viruses and verify the efficiency of the presented hypothesis.
Author Contributions
J.J.S.-F. and M.J.B.A. contributed equally to the idea and writing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the European Commission Project H2020-NMBP-2017 BIORIMA: BIOmaterial Risk MAnagement (GA No. 760928) and CESAM (UIDB/50017/2020 and UIDP/50017/2020), to FCT/MEC through national funds, and the co-funding by the FEDER, within the PT2020 Partnership Agreement and Compete 2020.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data are available in the main text.
Acknowledgments
We thank the editor and the anonymous reviewers for their constructive comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shen, Y. Earthworms in Traditional Chinese Medicine. Zool. Middle East 2010, 51, 171–173. [Google Scholar] [CrossRef]
- Darwin, C. The Formation of Vegetable Mould through the Action of Worms: With Observations on Their Habits; John Murray: London, UK, 1881. [Google Scholar]
- Phillips, H.R.P.; Bach, E.M.; Bartz, M.L.C.; Bennett, J.M.; Beugnon, R.; Briones, M.J.I.; Brown, G.G.; Ferlian, O.; Gongalsky, K.B.; Guerra, C.A.; et al. Global data on earthworm abundance, biomass, diversity and corresponding environmental properties. Sci. Data 2021, 8, 1–12. [Google Scholar] [CrossRef]
- OECD. OECD Guideline for Testing of Chemicals; No. 207: Earthworm, Acute Toxicity Tests; Organisation for Economic Co-operation and Development; OECD: Paris, France, 1984. [Google Scholar]
- Scott-Fordsmand, J.J.; Krogh, P.H.; Schaefer, M.; Johansen, A. The toxicity testing of double-walled nanotubes-contaminated food to Eisenia veneta earthwormes. Ecotoxicol. Environ. Saf. 2008, 71, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Scott-Fordsmand, J.J.; Weeks, J.M. Biomarkers in earthworms. Rev. Environ. Contam. Toxicol. 2000, 165, 117–159. [Google Scholar] [PubMed]
- Ribeiro, M.J.; Amorim, M.J.B.; Scott-Fordsmand, J.J. Cell In Vitro Testing with Soil Invertebrates-Challenges and Opportunities toward Modeling the Effect of Nanomaterials: A Surface-Modified CuO Case Study. Nanomaterials 2019, 9, 1087. [Google Scholar] [CrossRef] [Green Version]
- Gomes, S.I.L.; Hansen, D.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Effects of silver nanoparticles to soil invertebrates. Oxidative Stress Biomark. Eisenia Fetida 2015, 199, 49–55. [Google Scholar] [CrossRef]
- Mariyadas, J.; Amorim, M.J.B.; Jensen, J.; Scott-Fordsmand, J.J. Earthworm avoidance of silver nanomaterials over time. Environ. Pollut. 2018, 239, 751–756. [Google Scholar] [CrossRef]
- Hayashi, Y.; Heckmann, L.-H.; Simonsen, V.; Scott-Fordsmand, J.J. Time-course profiling of molecular stress responses to silver nanoparticles in the earthworm Eisenia fetida. Ecotoxicol. Environ. Saf. 2013, 98, 219–226. [Google Scholar] [CrossRef]
- Boraschi, D.; Alijagic, A.; Auguste, M.; Barbero, F.; Ferrari, E.; Hernadi, S.; Mayall, C.; Michelini, S.; Navarro Pacheco, N.I.; Prinelli, A.; et al. Addressing Nanomaterial Immunosafety by Evaluating Innate Immunity across Living Species. Small 2020, 16, e2000598. [Google Scholar] [CrossRef]
- Hayashi, Y.; Engelmann, P.; Foldbjerg, R.; Szabo, M.; Somogyi, I.; Pollak, E.; Molnar, L.; Autrup, H.; Sutherland, D.S.; Scott-Fordsmand, J.; et al. Earthworms and humans in vitro: Characterizing evolutionarily conserved stress and immune responses to silver nanoparticles. Environ. Sci. Technol. 2012, 46, 4166–4173. [Google Scholar] [CrossRef]
- Hayashi, Y.; Miclaus, T.; Engelmann, P.; Autrup, H.; Sutherland, D.S.; Scott-Fordsmand, J.J. Nanosilver pathophysiology in earthworms: Transcriptional profiling of secretory proteins and the implication for the protein corona. Nanotoxicology 2016, 10, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Swiderska, B.; Kedracka-Krok, S.; Plytycz, B. Data on proteins of lysenin family in coelomocytes of Eisenia andrei and E. fetida obtained by tandem mass spectrometry coupled with liquid chromatography. Data Brief 2016, 9, 629–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swiderska, B.; Kedracka-Krok, S.; Panz, T.; Morgan, A.J.; Falniowski, A.; Grzmil, P.; Plytycz, B. Lysenin family proteins in earthworm coelomocytes—Comparative approach. Dev. Comp. Immunol. 2017, 67, 404–412. [Google Scholar] [CrossRef]
- Abu-Farha, M.; Thanaraj, T.A.; Qaddoumi, M.G.; Hashem, A.; Abubaker, J.; Al-Mulla, F. The Role of Lipid Metabolism in COVID-19 Virus Infection and as a Drug Target. Int. J. Mol. Sci. 2020, 21, 3544. [Google Scholar] [CrossRef] [PubMed]
- Arvin, A.M.; Fink, K.; Schmid, M.A.; Cathcart, A.; Spreafico, R.; Havenar-Daughton, C.; Lanzavecchia, A.; Corti, D.; Virgin, H.W. A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nature 2020, 584, 353–363. [Google Scholar] [CrossRef]
- DeKosky, B.J. A molecular trap against COVID-19. Science 2020, 369, 1167–1168. [Google Scholar] [CrossRef]
- Hussain, A.; Kaler, J.; Tabrez, E.; Tabrez, S.; Tabrez, S.S.M. Novel COVID-19: A Comprehensive Review of Transmission, Manifestation, and Pathogenesis. Cureus 2020, 12, e8184. [Google Scholar] [CrossRef]
- Jones, G.W.; Monopoli, M.P.; Campagnolo, L.; Pietroiusti, A.; Tran, L.; Fadeel, B. No small matter: A perspective on nanotechnology-enabled solutions to fight COVID-19. Nanomedicine 2020, 17, 2411–2427. [Google Scholar] [CrossRef]
- Kinaret, P.A.S.; Del Giudice, G.; Greco, D. COVID-19 acute responses and possible long term consequences: What nanotoxicology can teach us. Nano Today 2020, 35, 100945. [Google Scholar] [CrossRef]
- Kuri-Cervantes, L.; Pampena, M.B.; Meng, W.; Rosenfeld, A.M.; Ittner, C.A.G.; Weisman, A.R.; Agyekum, R.S.; Mathew, D.; Baxter, A.E.; Vella, L.A.; et al. Comprehensive mapping of immune perturbations associated with severe COVID-19. Sci. Immunol. 2020, 5, eabd7114. [Google Scholar] [CrossRef]
- Yan, V.C.; Muller, F.L. Advantages of the Parent Nucleoside GS-441524 over Remdesivir for COVID-19 Treatment. ACS Med. Chem. Lett. 2020, 11, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Schaulies, S.; Schumacher, F.; Wigger, D.; Schöl, M.; Waghmare, T.; Schlegel, J.; Seibel, J.; Kleuser, B. Sphingolipids: Effectors and Achilles Heals in Viral Infections? Cells 2021, 10, 2175. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.J.; Becker, K.A.; Gripp, B.; Hoffmann, M.; Keitsch, S.; Wilker, B.; Soddemann, M.; Gulbins, A.; Carpinteiro, E.; Patel, S.H.; et al. Sphingosine prevents binding of SARS-CoV-2 spike to its cellular receptor ACE2. J. Biol. Chem. 2020, 295, 15174–15182. [Google Scholar] [CrossRef]
- Makino, A.; Abe, M.; Ishitsuka, R.; Murate, M.; Kishimoto, T.; Sakai, S.; Hullin-Matsuda, F.; Shimada, Y.; Inaba, T.; Miyatake, H.; et al. A novel sphingomyelin/cholesterol domain-specific probe reveals the dynamics of the membrane domains during virus release and in Niemann- Pick type C. FASEB J. 2017, 31, 1301–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makino, A.; Abe, M.; Murate, M.; Inaba, T.; Yilmaz, N.; Hullin-Matsuda, F.; Kishimoto, T.; Schieber, N.L.; Taguchi, T.; Arai, H.; et al. Visualization of the heterogeneous membrane distribution of sphingomyelin associated with cytokinesis, cell polarity, and sphingolipidosis. FASEB J. 2015, 29, 477–493. [Google Scholar] [CrossRef] [PubMed]
- Meacci, E.; Garcia-Gil, M.; Pierucci, F. SARS-CoV-2 Infection: A Role for S1P/S1P Receptor Signaling in the Nervous System? Int. J. Mol. Sci. 2020, 21, 6773. [Google Scholar] [CrossRef]
- Bokori-Brown, M.; Martin, T.G.; Naylor, C.E.; Basak, A.K.; Titball, R.W.; Savva, C.G. Cryo-EM structure of lysenin pore elucidates membrane insertion by an aerolysin family protein. Nat. Commun. 2016, 7, 11293. [Google Scholar] [CrossRef] [Green Version]
- Kulma, M.; Herec, M.; Grudzinski, W.; Anderluh, G.; Gruszecki, W.I.; Kwiatkowska, K.; Sobota, A. Sphingomyelin-rich domains are sites of lysenin oligomerization: Implications for raft studies. Biochim. Biophys. Acta-Biomembr. 2010, 1798, 471–481. [Google Scholar] [CrossRef]
- Ando, J.; Kinoshita, M.; Cui, J.; Yamakoshi, H.; Dodo, K.; Fujita, K.; Murata, M.; Sodeoka, M. Sphingomyelin distribution in lipid rafts of artificial monolayer membranes visualized by Raman microscopy. Proc. Natl. Acad. Sci. USA 2015, 112, 4558–4563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, S.; Nüssler, F.; Kauschke, E.; Lutsch, G.; Cooper, E.L.; Herrmann, A. Interaction of earthworm hemolysin with lipid membranes requires sphingolipids. J. Biol. Chem. 1997, 272, 20884–20892. [Google Scholar] [CrossRef] [Green Version]
- Lange, S.; Kauschke, E.; Mohrig, W.; Cooper, E.L. Biochemical characteristics of Eiseniapore, a pore-forming protein in the coelomic fluid of earthworms. Eur. J. Biochem. 1999, 262, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Shogomori, H.; Kobayashi, T. Lysenin: A sphingomyelin specific pore-forming toxin. Biochim. Biophys. Acta 2008, 1780, 612–618. [Google Scholar] [CrossRef]
- Audi, A.; Soudani, N.; Dbaibo, G.; Zaraket, H. Depletion of Host and Viral Sphingomyelin Impairs Influenza Virus Infection. Front. Microbiol. 2020, 11, 612. [Google Scholar] [CrossRef]
- Callens, N.; Brugger, B.; Bonnafous, P.; Drobecq, H.; Gerl, M.J.; Krey, T.; Roman-Sosa, G.; Rumenapf, T.; Lambert, O.; Dubuisson, J.; et al. Morphology and Molecular Composition of Purified Bovine Viral Diarrhea Virus Envelope. PLoS Path. 2016, 12, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Oliva, A.; Ortega-Gonzalez, P.; Risco, C. Targeting host lipid flows: Exploring new antiviral and antibiotic strategies. Cell. Microbiol. 2019, 21, e12996. [Google Scholar] [CrossRef] [Green Version]
- Ketter, E.; Randall, G. Virus Impact on Lipids and Membranes. Annu. Rev. Virol. 2019, 6, 319–340. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, A.; Sekizawa, Y.; Emoto, K.; Sakuraba, H.; Inoue, K.; Kobayashi, H.; Umeda, M. Lysenin, a novel sphingomyelin-specific binding protein. J. Biol. Chem. 1998, 273, 5300–5306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, I.; Jantzen, E. Sphingolipids in Bacteria and Fungi. Anaerobe 2001, 7, 103–112. [Google Scholar] [CrossRef]
- Bagam, P.; Singh, D.P.; Inda, M.E.; Batra, S. Unraveling the role of membrane microdomains during microbial infections. Cell Biol. Toxicol. 2017, 33, 429–455. [Google Scholar] [CrossRef]
- Brügger, B.; Glass, B.; Haberkant, P.; Leibrecht, I.; Wieland, F.T.; Kräusslich, H.G. The HIV lipidome: A raft with an unusual composition. Proc. Natl. Acad. Sci. USA 2006, 103, 2641–2646. [Google Scholar] [CrossRef] [Green Version]
- Martin-Acebes, M.A.; Vazquez-Calvo, A.; Saiz, J.C. Lipids and flaviviruses, present and future perspectives for the control of dengue, Zika, and West Nile viruses. Prog. Lipid Res. 2016, 64, 123–137. [Google Scholar] [CrossRef]
- Zhang, Z.; He, G.; Filipowicz, N.A.; Randall, G.; Belov, G.A.; Kopek, B.G.; Wang, X. Host Lipids in Positive-Strand RNA Virus Genome Replication. Front. Microbiol. 2019, 10, 286. [Google Scholar] [CrossRef] [PubMed]
- Millet, J.K.; Whittaker, G.R. Physiological and molecular triggers for SARS-CoV membrane fusion and entry into host cells. Virology 2018, 517, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Martín-Acebes, M.A.; Merino-Ramos, T.; Blázquez, A.B.; Casas, J.; Escribano-Romero, E.; Sobrino, F.; Saiz, J.C. The composition of West Nile virus lipid envelope unveils a role of sphingolipid metabolism in flavivirus biogenesis. J. Virol. 2014, 88, 12041–12054. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yang, P.; Liu, K.; Guo, F.; Zhang, Y.; Zhang, G.; Jiang, C. SARS coronavirus entry into host cells through a novel clathrin- and caveolae-independent endocytic pathway. Cell Res. 2008, 18, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghunathan, K.; Kenworthy, A.K. Dynamic pattern generation in cell membranes: Current insights into membrane organization. Biochim. Biophys. Acta Biomembr. 2018, 1860, 2018–2031. [Google Scholar] [CrossRef] [PubMed]
- Prakash, H.; Upadhyay, D.; Bandapalli, O.R.; Jain, A.; Kleuser, B. Host sphingolipids: Perspective immune adjuvant for controlling SARS-CoV-2 infection for managing COVID-19 disease. Prostaglandins Other Lipid Mediat. 2020, 152, 106504. [Google Scholar] [CrossRef]
- Ikenouchi, J.; Suzuki, M.; Umeda, K.; Ikeda, K.; Taguchi, R.; Kobayashi, T.; Sato, S.B.; Kobayashi, T.; Stolz, D.B.; Umeda, M. Lipid polarity is maintained in absence of tight junctions. J. Biol. Chem. 2012, 287, 9525–9533. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Wang, J.; Zhang, J.; Yu, B.; Niu, B. An extract from the earthworm Eisenia fetida non-specifically inhib-its the activity of influenza and adenoviruses. J. Tradit. Chin. Med. 2012, 32, 657–663. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.E.; Adhikary, S.; Kolokoltsov, A.A.; Davey, R.A. Ebolavirus requires acid sphingomyelinase activity and plasma membrane sphingomyelin for infection. J. Virol. 2012, 86, 7473–7483. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.; Sugita, M. Sphingolipids in lower animals. Prog. Lipid Res. 1993, 32, 25–45. [Google Scholar] [CrossRef]
- Kobayashi, H.; Ohtomi, M.; Sekizawa, Y.; Ohta, N. Toxicity of coelomic fluid of the earthworm Eisenia foetida to vertebrates but not invertebrates: Probable role of sphingomyelin. Comp. Biochem. Physiol. C Toxicol. Pharm. 2001, 128, 401–411. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Baixauli, F.; Gutiérrez-Vázquez, C.; Sánchez-Madrid, F.; Mittelbrunn, M. Sorting it out: Regulation of exosome loading. Semin. Cancer Biol. 2014, 28, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Augustine, D.; Rao, R.S.; Anbu, J.; Chidambara Murthy, K.N. In vitro cytotoxic and apoptotic induction effect of earthworm coelomic fluid of Eudrilus eugeniae, Eisenia foetida, and Perionyx excavatus on human oral squamous cell carcinoma-9 cell line. Toxicol. Rep. 2019, 6, 347–357. [Google Scholar] [CrossRef]
- Hullin-Matsuda, F.; Murate, M.; Kobayashi, T. Protein probes to visualize sphingomyelin and ceramide phosphoethanolamine. Chem. Phys. Lipids 2018, 216, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, K.; Hordejuk, R.; Szymczyk, P.; Kulma, M.; Abdel-Shakor, A.B.; Płucienniczak, A.; Dołowy, K.; Szewczyk, A.; Sobota, A. Lysenin-His, a sphingomyelin-recognizing toxin, requires tryptophan 20 for cation-selective channel assembly but not for membrane binding. Mol. Membr. Biol. 2007, 24, 121–134. [Google Scholar] [CrossRef]
- Bogard, A.; Abatchev, G.; Hutchinson, Z.; Ward, J.; Finn, P.W.; McKinney, F.; Fologea, D. Lysenin Channels as Sensors for Ions and Molecules. Sensors 2020, 20, 6099. [Google Scholar] [CrossRef] [PubMed]
- Martin-Acebes, M.A.; de Oya, N.J.; Saiz, J.C. Lipid Metabolism as a Source of Druggable Targets for Antiviral Discovery against Zika and Other Flaviviruses. Pharmaceuticals 2019, 12, 97. [Google Scholar] [CrossRef] [Green Version]
- Cordier, T.; Alonso-Sáez, L.; Apothéloz-Perret-Gentil, L.; Aylagas, E.; Bohan, D.A.; Bouchez, A.; Chariton, A.; Creer, S.; Frühe, L.; Keck, F.; et al. Ecosystems monitoring powered by environmental genomics: A review of current strategies with an implementation roadmap. Mol. Ecol. 2020, 30, 2937–2958. [Google Scholar] [CrossRef] [PubMed]
- Molinos-Albert, L.M.; Bilbao, E.; Agulló, L.; Marfil, S.; García, E.; Rodríguez de la Concepción, M.L.; Izquierdo-Useros, N.; Vilaplana, C.; Nieto-Garai, J.A.; Contreras, F.X.; et al. Proteoliposomal formulations of an HIV-1 gp41-based miniprotein elicit a lipid-dependent immunodominant response overlapping the 2F5 binding motif. Sci. Rep. 2017, 7, 40800. [Google Scholar] [CrossRef]
- Molinos-Albert, L.M.; Clotet, B.; Blanco, J.; Carrillo, J. Immunologic Insights on the Membrane Proximal External Region: A Major Human Immunodeficiency Virus Type-1 Vaccine Target. Front. Immunol. 2017, 8, 1154. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Yang, C.; Xu, X.-f.; Xu, W.; Liu, S.-w. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, A.; Rohatgi, R.; Siebold, C. Cholesterol access in cellular membranes controls Hedgehog signaling. Nat. Chem. Biol. 2020, 16, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Kocar, E.; Rezen, T.; Rozman, D. Cholesterol, lipoproteins, and COVID-19: Basic concepts and clinical applications. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158849. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, S.-I.; Zhang, J.; Zhang, S.; Shoemaker, C.B.; Dong, M. Delivery of single-domain antibodies into neurons using a chimeric toxin–based platform is therapeutic in mouse models of botulism. Sci. Transl. Med. 2021, 13, eaaz4197. [Google Scholar] [CrossRef] [PubMed]
- Terán-Navarro, H.; Calderon-Gonzalez, R.; Salcines-Cuevas, D.; García, I.; Marradi, M.; Freire, J.; Salmon, E.; Portillo-Gonzalez, M.; Frande-Cabanes, E.; García-Castaño, A.; et al. Pre-clinical development of Listeria-based nanovaccines as immunotherapies for solid tumours: Insights from melanoma. Oncoimmunology 2018, 8, e1541534. [Google Scholar] [CrossRef] [Green Version]
- Pindiprolu, S.K.S.; Kumar, C.S.P.; Kumar Golla, V.S.K.; Likitha, P.; Chandra, S.; Chandra, R.K. Pulmonary delivery of nanostructured lipid carriers for effective repurposing of salinomycin as an antiviral agent. Med. Hypotheses 2020, 143, 109858. [Google Scholar] [CrossRef]
- Bulbake, U.; Doppalapudi, S.; Kommineni, N.; Khan, W. Liposomal Formulations in Clinical Use: An Updated Review. Pharmaceutics 2017, 9, 12. [Google Scholar] [CrossRef]
- Abatchev, G.; Bogard, A.; Hutchinson, Z.; Ward, J.; Fologea, D. Rapid Production and Purification of Dye-Loaded Liposomes by Electrodialysis-Driven Depletion. Membranes 2021, 11, 417. [Google Scholar] [CrossRef]
- Bodó, K.; Boros, Á.; Rumpler, É.; Molnár, L.; Böröcz, K.; Németh, P.; Engelmann, P. Identification of novel lumbricin homologues in Eisenia andrei earthworms. Dev. Comp. Immunol. 2019, 90, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Xia, S. Biological mechanisms and therapeutic relevance of the gasdermin family. Mol. Asp. Med. 2020, 76, 100890. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Gandhi, S.; Lata, K.; Chattopadhyay, K. Pore-forming toxins in infection and immunity. Biochem. Soc. Trans. 2021, 49, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Al Mamun, A.; Wu, Y.; Monalisa, I.; Jia, C.; Zhou, K.; Munir, F.; Xiao, J. Role of pyroptosis in spinal cord injury and its therapeutic implications. J. Adv. Res. 2020, 28, 97–109. [Google Scholar] [CrossRef] [PubMed]
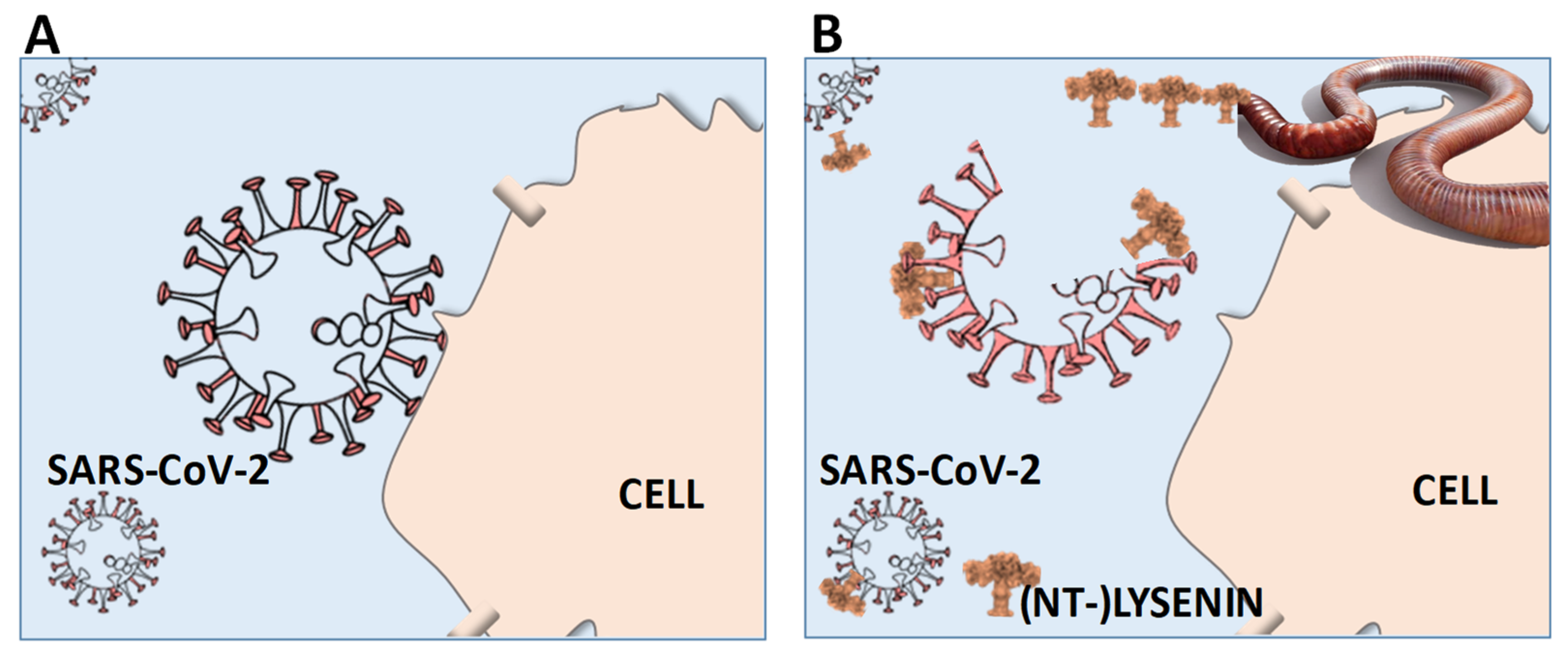
Figure 1.
Schematic (hypothetical) outline of the therapeutic potential of lysenin (-derivatives) to prevent or treat human SARS-CoV-2 (COVID-19) infections. (A) No action: SARS-CoV-2 enters the organism and attaches to the cell membranes, allowing the RNA injection and infection. (B) (NT-)lysenin administration: SARS-CoV-2 enters the organism and encounters lysenin, which binds to sphingomyelin in SARS-CoV-2 envelope (corona), forming pores and disrupting it, hence no cell membrane attachment and, thus, no infection. (NT-)LYSENIN: Non-Toxic-Lysenin: representation of a modified lysenin derivative.
Figure 1.
Schematic (hypothetical) outline of the therapeutic potential of lysenin (-derivatives) to prevent or treat human SARS-CoV-2 (COVID-19) infections. (A) No action: SARS-CoV-2 enters the organism and attaches to the cell membranes, allowing the RNA injection and infection. (B) (NT-)lysenin administration: SARS-CoV-2 enters the organism and encounters lysenin, which binds to sphingomyelin in SARS-CoV-2 envelope (corona), forming pores and disrupting it, hence no cell membrane attachment and, thus, no infection. (NT-)LYSENIN: Non-Toxic-Lysenin: representation of a modified lysenin derivative.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Scott-Fordsmand, J.J.; Amorim, M.J.B. The Curious Case of Earthworms and COVID-19. Biology 2021, 10, 1043. https://doi.org/10.3390/biology10101043
AMA Style
Scott-Fordsmand JJ, Amorim MJB. The Curious Case of Earthworms and COVID-19. Biology. 2021; 10(10):1043. https://doi.org/10.3390/biology10101043
Chicago/Turabian StyleScott-Fordsmand, Janeck J., and Monica J. B. Amorim. 2021. "The Curious Case of Earthworms and COVID-19" Biology 10, no. 10: 1043. https://doi.org/10.3390/biology10101043
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.