
Enhancement of β-Alanine Biosynthesis in Escherichia coli Based on Multivariate Modular Metabolic Engineering

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Media and Culture Conditions
2.3. Measurement of Amino Acid Concentration
3. Results and Discussion
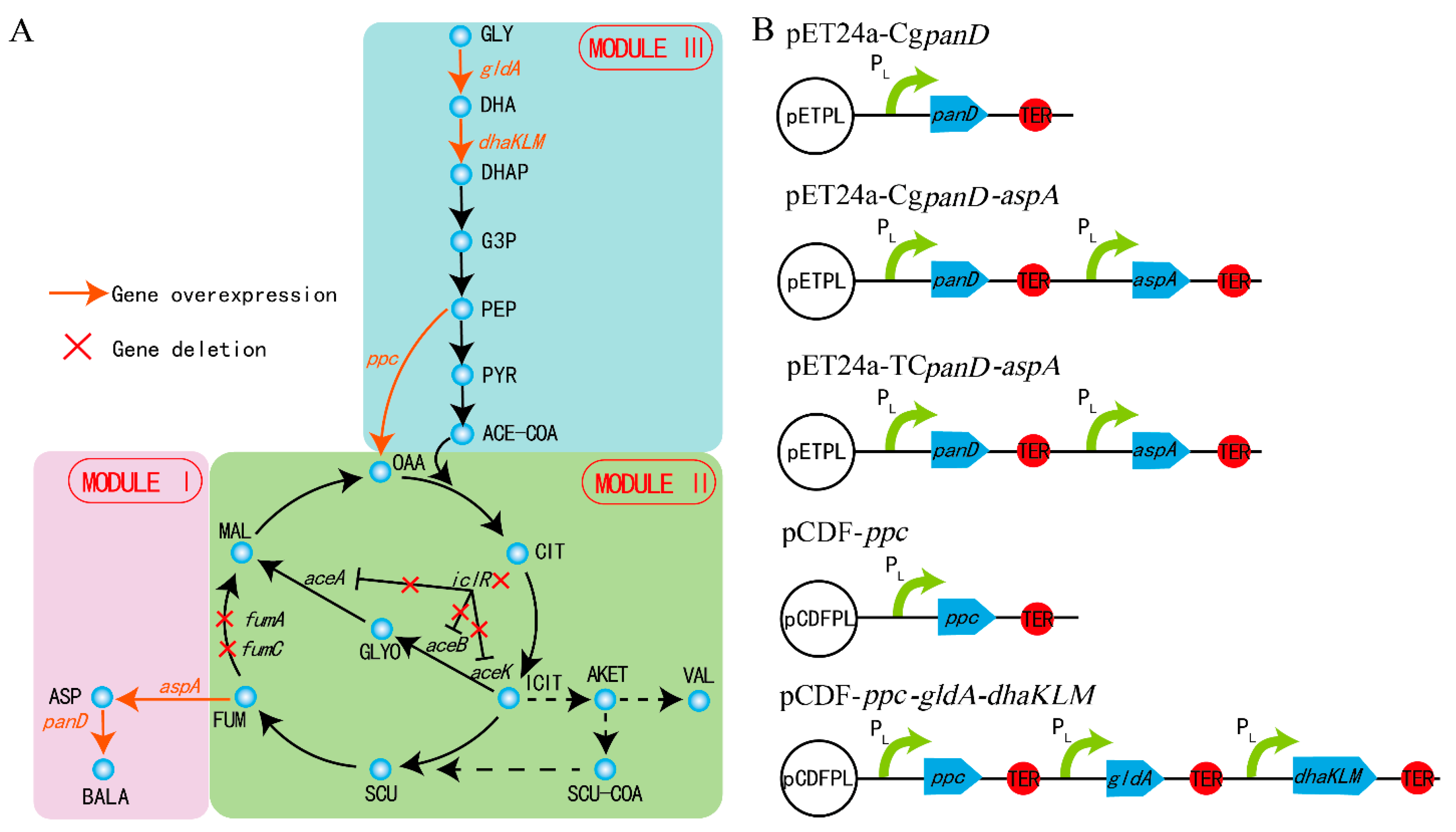
3.1. Module Separation of β-Alanine Biosynthesis
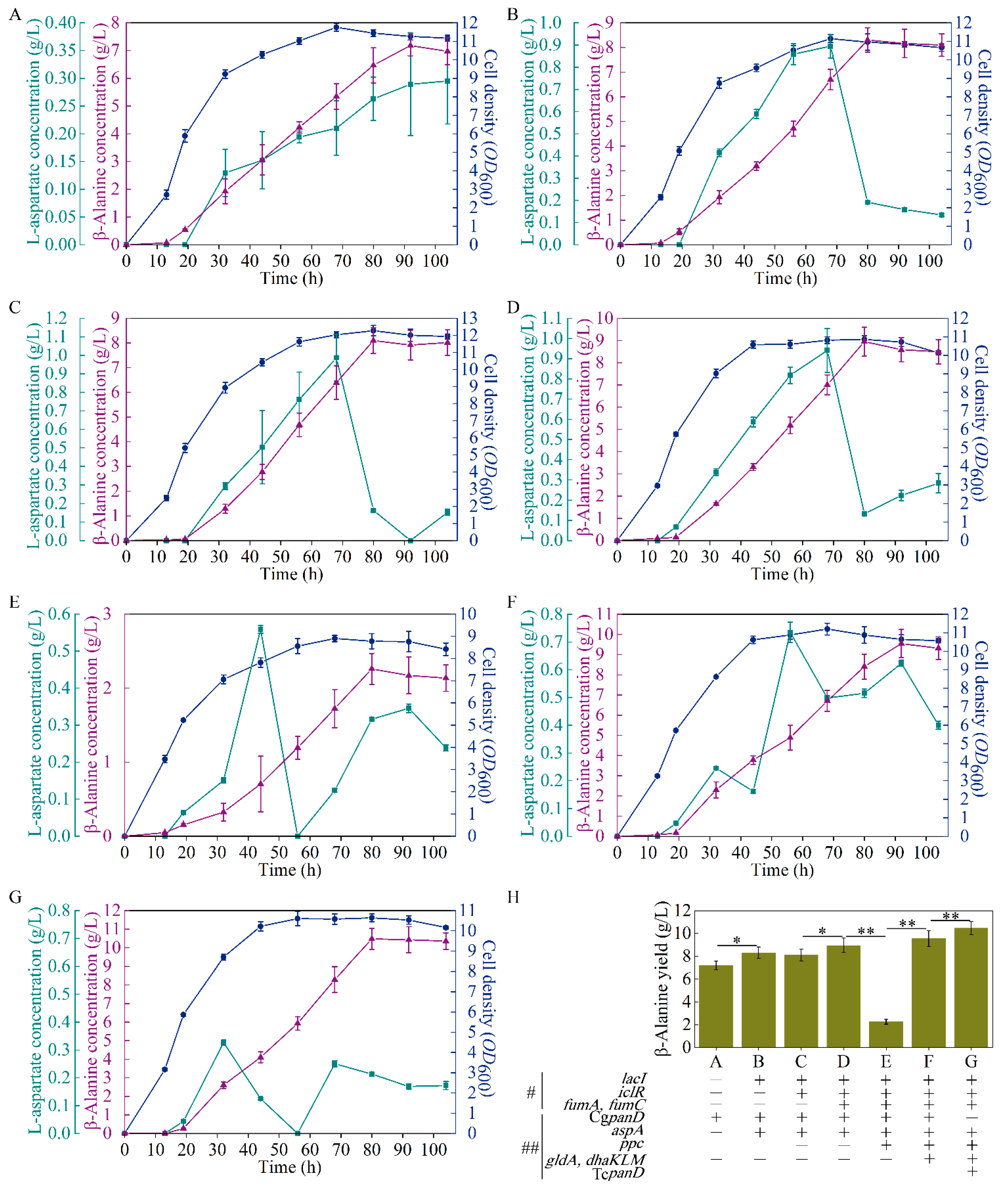
3.2. Regulation in the β-Alanine Biosynthesis Module
3.3. Regulation between the β-Alanine Biosynthesis Module and the TCA Module
3.4. Regulation in the TCA Module
3.5. Regulation between the TCA Module and the Glycolysis Module
3.6. Regulation in the Glycolysis Module
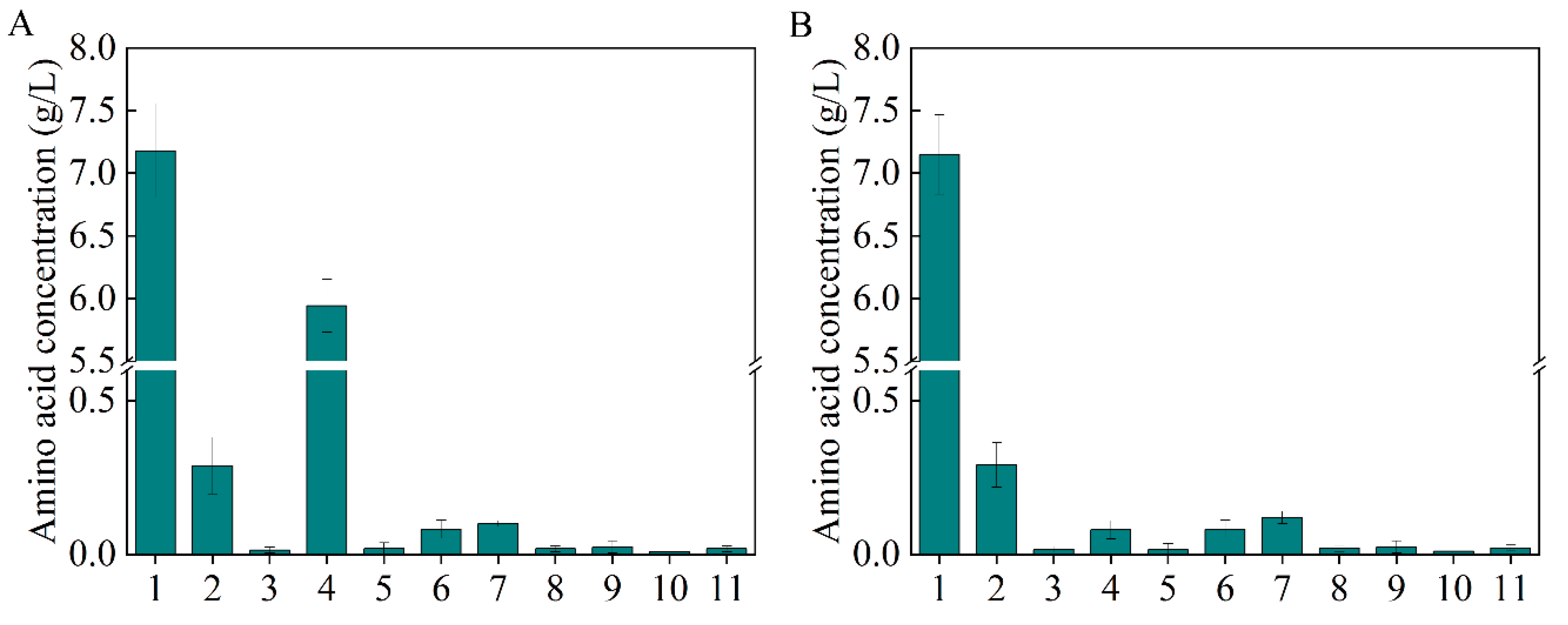
3.7. Intensive Regulation in the β-Alanine Biosynthesis Module
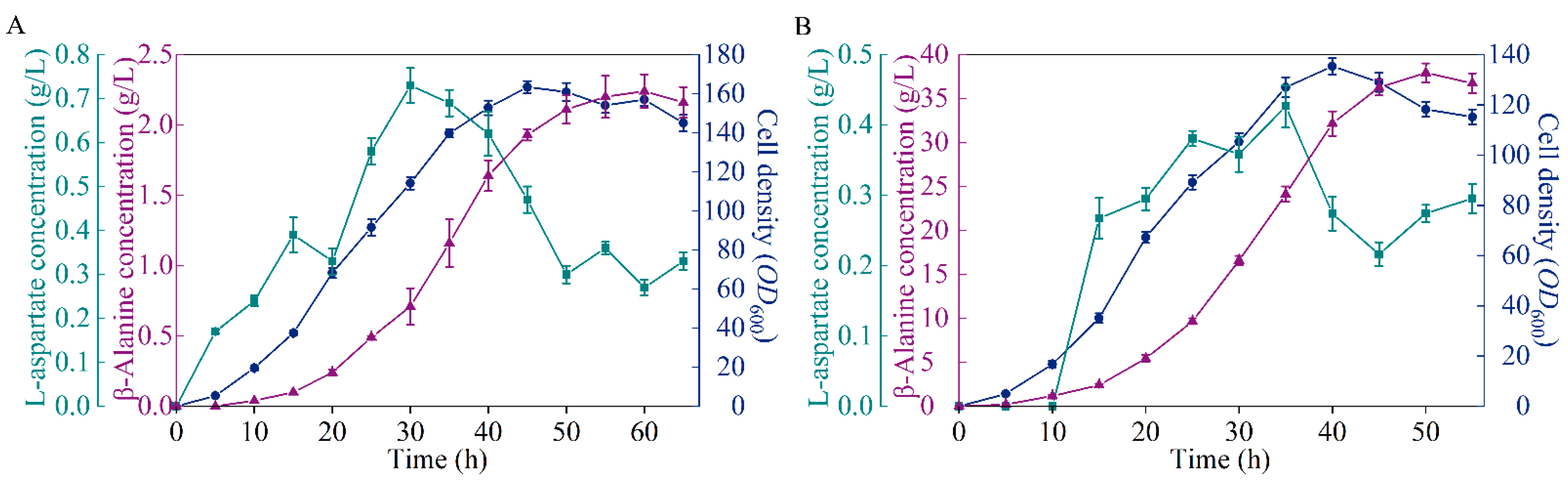
3.8. Performance of the Microbial Cell Factory in Fed-Batch Fermentation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Itagaki, T.; Komori, Y.; Sugahara, Y.; Kuroda, K. Synthesis of a kaolinite–poly(β-alanine) intercalation compound. J. Mater. Chem. 2001, 11, 3291–3295. [Google Scholar] [CrossRef]
- Yang, H.; Zhao, J.; Yan, M.; Pispas, S.; Zhang, G. Nylon 3 synthesized by ring opening polymerization with a metal-free catalyst. Polym. Chem. 2011, 2, 2888–2892. [Google Scholar] [CrossRef]
- Könst, P.M.; Franssen, M.C.R.; Scott, E.L.; Sanders, J.P.M. A study on the applicability of l-aspartate α-decarboxylase in the biobased production of nitrogen containing chemicals. Green Chem. 2009, 11, 1646–1652. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, L.; Li, Y.; Zhang, L.; Shi, G. Synthesis of β-alanine from l-aspartate using l-aspartate-α-decarboxylase from Corynebacterium glutamicum. Biotechnol. Lett. 2014, 36, 1681–1686. [Google Scholar] [CrossRef]
- Wang, L.; Piao, X.; Cui, S.; Hu, M.; Tao, Y. Enhanced production of β-alanine through co-expressing two different subtypes of l-aspartate-α-decarboxylase. J. Ind. Microbiol. Biotechnol. 2020, 47, 465–474. [Google Scholar] [CrossRef]
- Zou, X.; Guo, L.; Huang, L.; Li, M.; Zhang, S.; Yang, A.; Zhang, Y.; Zhu, L.; Zhang, H.; Zhang, J.; et al. Pathway construction and metabolic engineering for fermentative production of β-alanine in Escherichia coli. Appl. Microbiol. Biotechnol. 2020, 104, 2545–2559. [Google Scholar] [CrossRef]
- Cheng, K.-K.; Wang, G.-Y.; Zeng, J.; Zhang, J.-A. Improved Succinate Production by Metabolic Engineering. BioMed Res. Int. 2013, 2013, 538790. [Google Scholar] [CrossRef] [Green Version]
- Pontrelli, S.; Chiu, T.-Y.; Lan, E.I.; Chen, F.Y.H.; Chang, P.; Liao, J.C. Escherichia coli as a host for metabolic engineering. Metab. Eng. 2018, 50, 16–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, K.V.; Sanders, T.M.; Prather, K.L.J. A dynamic metabolite valve for the control of central carbon metabolism. Metab. Eng. 2012, 14, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, P.; Fan, Y.; Bao, H.; Du, G.; Zhou, J.; Chen, J. Multivariate modular metabolic engineering of Escherichia coli to produce resveratrol from l-tyrosine. J. Biotechnol. 2013, 167, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Biggs, B.W.; De Paepe, B.; Santos, C.N.S.; De Mey, M.; Kumaran Ajikumar, P. Multivariate modular metabolic engineering for pathway and strain optimization. Curr. Opin. Biotechnol. 2014, 29, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Chen, J.; Zhang, X. Systematic engineering of pentose phosphate pathway improves Escherichia coli succinate production. Biotechnol. Biofuels 2016, 9, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.; Li, D.; Kang, J.; Jiang, P.; Sun, J.; Zhang, D. Metabolic engineering of Escherichia coli for de novo biosynthesis of vitamin B12. Nat. Commun. 2018, 9, 4917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, X.; Zhang, H. Balancing the non-linear rosmarinic acid biosynthetic pathway by modular co-culture engineering. Metab. Eng. 2019, 54, 1–11. [Google Scholar] [CrossRef]
- Piao, X.; Wang, L.; Lin, B.; Chen, H.; Liu, W.; Tao, Y. Metabolic engineering of Escherichia coli for production of L-aspartate and its derivative β-alanine with high stoichiometric yield. Metab. Eng. 2019, 54, 244–254. [Google Scholar] [CrossRef]
- Song, C.W.; Lee, J.; Ko, Y.-S.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of 3-aminopropionic acid. Metab. Eng. 2015, 30, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhu, Y.; Zhou, Z. Systematic engineering of the rate-limiting step of β-alanine biosynthesis in Escherichia coli. Electron. J. Biotechnol. 2021, 51, 88–94. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, L.; Yin, M.; Zhou, Z. Novel Mode Engineering for β-Alanine Production in Escherichia coli with the Guide of Adaptive Laboratory Evolution. Microorganisms 2021, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, Z.; Cui, W.; Zhou, Z. Establishment of Bioprocess for Synthesis of Nicotinamide by Recombinant Escherichia coli Expressing High-Molecular-Mass Nitrile Hydratase. Appl. Biochem. Biotechnol. 2017, 182, 1458–1466. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, S.Y. Enhanced production of poly(3-hydroxybutyrate) by filamentation-suppressed recombinantEscherichia coli in a defined medium. J. Environ. Polym. Degrad. 1996, 4, 131–134. [Google Scholar] [CrossRef]
- Rea, K.; Cremers, T.I.F.H.; Westerink, B.H.C. HPLC conditions are critical for the detection of GABA by microdialysis. J. Neurochem. 2005, 94, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Ren, C.; Xu, Y. Deciphering the crucial roles of transcriptional regulator GadR on gamma-aminobutyric acid production and acid resistance in Lactobacillus brevis. Microb. Cell Factories 2019, 18, 108. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, Y.; Li, L.; Linhardt, R.J.; Yan, Y. Regulating malonyl-CoA metabolism via synthetic antisense RNAs for enhanced biosynthesis of natural products. Metab. Eng. 2015, 29, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yu, L.; Zhou, L.; Zhou, Z. One-Pot Biosynthesis of L-Aspartate from Maleate via an Engineered Strain Containing a Dual-Enzyme System. Appl. Environ. Microbiol. 2019, 85, e01327-19. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wei, H.; Wang, T.; Xu, Q.; Zhang, C.; Fan, X.; Ma, Q.; Chen, N.; Xie, X. Current status on metabolic engineering for the production of l-aspartate family amino acids and derivatives. Bioresour. Technol. 2017, 245, 1588–1602. [Google Scholar] [CrossRef]
- Kumar, P.; Dubey, K.K. Modulation of fatty acid metabolism and tricarboxylic acid cycle to enhance the lipstatin production through medium engineering in Streptomyces toxytricini. Bioresour. Technol. 2016, 213, 64–68. [Google Scholar] [CrossRef]
- Chen, S.; Li, S.; Huang, T.; Yang, S.; Liu, K.; Ma, B.; Shi, Y.; Miao, Y. Nitrate reduction by Paracoccus thiophilus strain LSL 251 under aerobic condition: Performance and intracellular central carbon flux pathways. Bioresour. Technol. 2020, 308, 123301. [Google Scholar] [CrossRef]
- Lama, S.; Kim, Y.; Nguyen, D.T.; Im, C.H.; Sankaranarayanan, M.; Park, S. Production of 3-hydroxypropionic acid from acetate using metabolically-engineered and glucose-grown Escherichia coli. Bioresour. Technol. 2021, 320, 124362. [Google Scholar] [CrossRef]
- Koendjbiharie, J.G.; van Kranenburg, R.; Kengen, S.W.M. The PEP-pyruvate-oxaloacetate node: Variation at the heart of metabolism. FEMS Microbiol. Rev. 2020, 45, 1–19. [Google Scholar] [CrossRef]
- Zhang, B.; Zhu, Y.; Zhang, J.; Wang, D.; Sun, L.; Hong, J. Engineered Kluyveromyces marxianus for pyruvate production at elevated temperature with simultaneous consumption of xylose and glucose. Bioresour. Technol. 2017, 224, 553–562. [Google Scholar] [CrossRef]
- Durnin, G.; Clomburg, J.; Yeates, Z.; Alvarez, P.J.J.; Zygourakis, K.; Campbell, P.; Gonzalez, R. Understanding and harnessing the microaerobic metabolism of glycerol in Escherichia coli. Biotechnol. Bioeng. 2009, 103, 148–161. [Google Scholar] [CrossRef]
- Williamson, J.M.; Brown, G.M. Purification and properties of L-Aspartate-alpha-decarboxylase, an enzyme that catalyzes the formation of beta-alanine in Escherichia coli. J. Biol. Chem. 1979, 254, 8074–8082. [Google Scholar] [CrossRef]
- Cui, W.; Shi, Z.; Fang, Y.; Zhou, L.; Ding, N.; Zhou, Z. Significance of Arg3, Arg54, and Tyr58 of l-aspartate α-decarboxylase from Corynebacterium glutamicum in the process of self-cleavage. Biotechnol. Lett. 2014, 36, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.I.; Suh, S.W. Crystal Structure of the Schiff Base Intermediate Prior to Decarboxylation in the Catalytic Cycle of Aspartate α-Decarboxylase. J. Mol. Biol. 2004, 340, 1–7. [Google Scholar] [CrossRef]
- Gopalan, G.; Chopra, S.; Ranganathan, A.; Swaminathan, K. Crystal structure of uncleaved L-aspartate-alpha-decarboxylase from Mycobacterium tuberculosis. Proteins 2006, 65, 796–802. [Google Scholar] [CrossRef]
- Dai, F.; Qiao, L.; Cao, C.; Liu, X.; Tong, X.; He, S.; Hu, H.; Zhang, L.; Wu, S.; Tan, D.; et al. Aspartate Decarboxylase is Required for a Normal Pupa Pigmentation Pattern in the Silkworm, Bombyx mori. Sci. Rep. 2015, 5, 10885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borodina, I.; Kildegaard, K.R.; Jensen, N.B.; Blicher, T.H.; Maury, J.; Sherstyk, S.; Schneider, K.; Lamosa, P.; Herrgård, M.J.; Rosenstand, I.; et al. Establishing a synthetic pathway for high-level production of 3-hydroxypropionic acid in Saccharomyces cerevisiae via β-alanine. Metab. Eng. 2015, 27, 57–64. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Description | Source |
|---|---|---|
| B0016-082BB | B0016 ΔackA-pta ΔpflB ΔadhE ΔfrdA ΔldhA ΔlysC ΔpanC Δpts, panD: CgpanD | [18] |
| B0016-01 | B0016-082BB, pETpL-CgpanD | This study |
| B0016-02 | B0016-01, ΔlacI, pETpL-CgpanD-aspA | This study |
| B0016-03 | B0016-02, ΔiclR, pETpL-CgpanD-aspA | This study |
| B0016-04 | B0016-03, ΔfumA, ΔfumC, pETpL-CgpanD-aspA | This study |
| B0016-05 | B0016-04, pETpL-CgpanD-aspA, pCDFPL-ppc | This study |
| B0016-06 | B0016-04, pETpL-CgpanD-aspA, pCDFPL-ppc-gldA-dhaKLM | This study |
| B0016-07 | B0016-06, pETpL-TcpanD-aspA, pCDFPL-ppc-gldA-dhaKLM | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Zhou, L.; Zhou, Z. Enhancement of β-Alanine Biosynthesis in Escherichia coli Based on Multivariate Modular Metabolic Engineering. Biology 2021, 10, 1017. https://doi.org/10.3390/biology10101017
Xu J, Zhou L, Zhou Z. Enhancement of β-Alanine Biosynthesis in Escherichia coli Based on Multivariate Modular Metabolic Engineering. Biology. 2021; 10(10):1017. https://doi.org/10.3390/biology10101017
Chicago/Turabian StyleXu, Jian, Li Zhou, and Zhemin Zhou. 2021. "Enhancement of β-Alanine Biosynthesis in Escherichia coli Based on Multivariate Modular Metabolic Engineering" Biology 10, no. 10: 1017. https://doi.org/10.3390/biology10101017