Effect of Weed Management on the Parasitoid Community in Mediterranean Vineyards

 ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
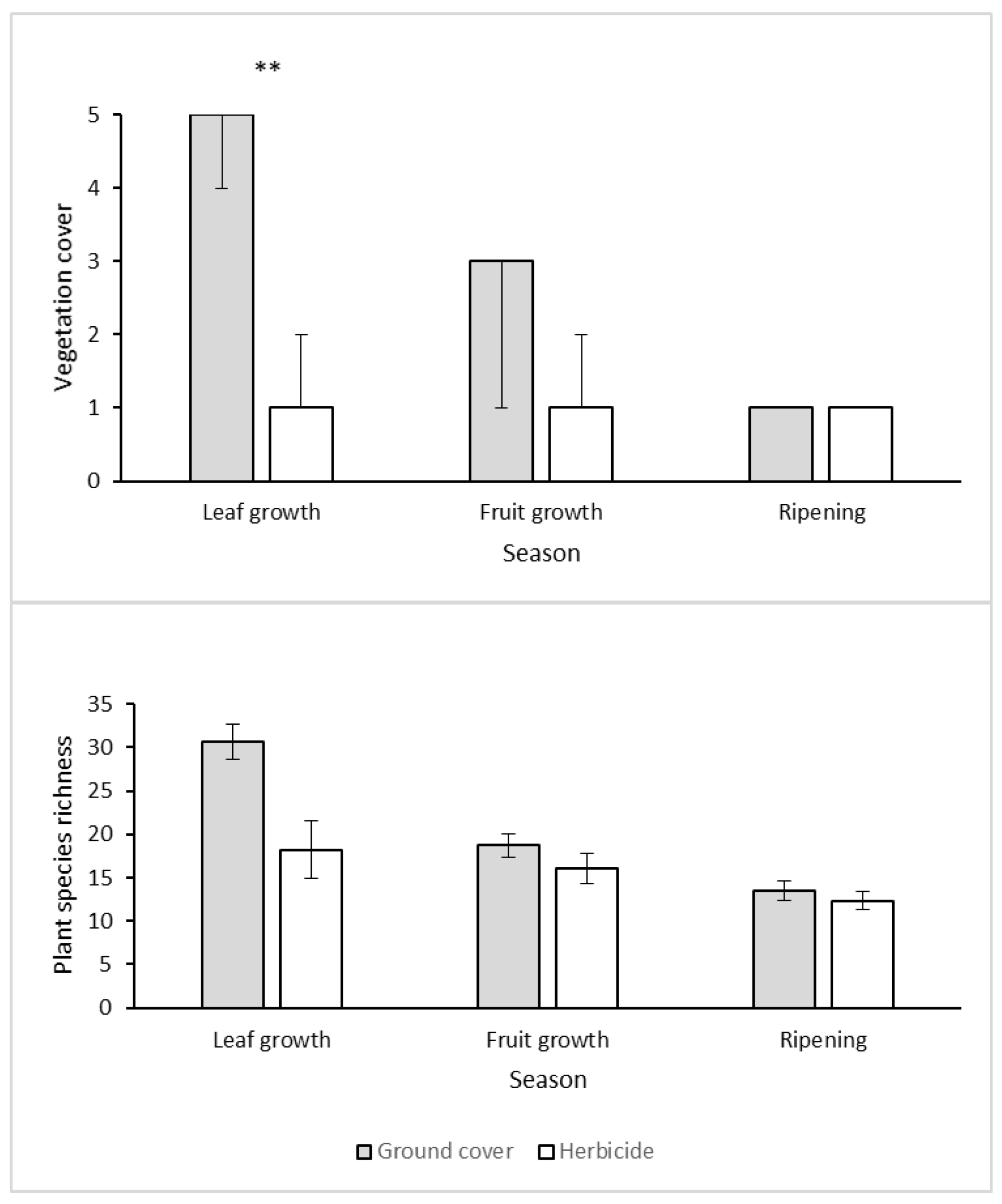
3.1. Vegetation Cover and Richness
3.2. Collected Arthropods and Parasitoids
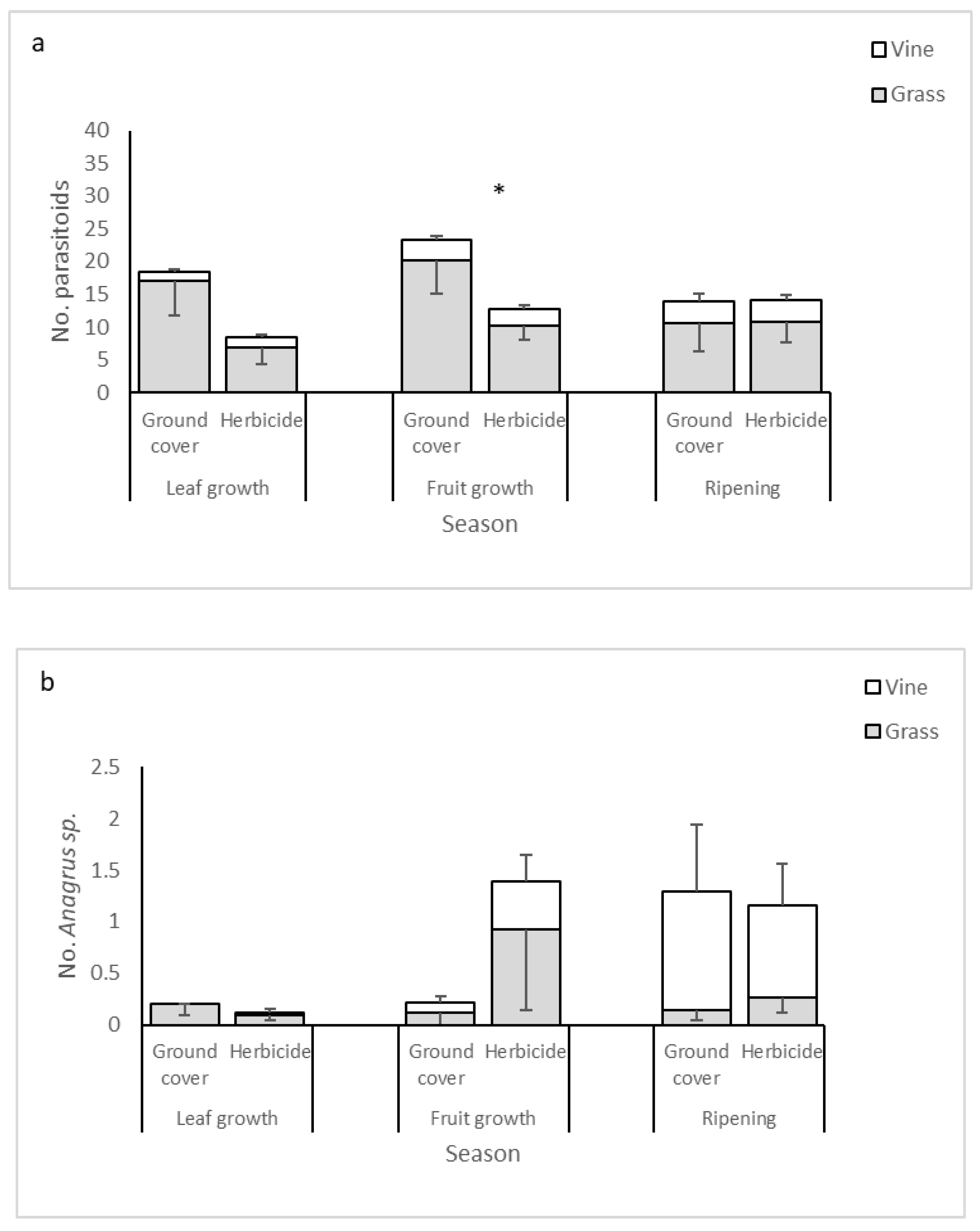
3.3. Parasitoid Abundances
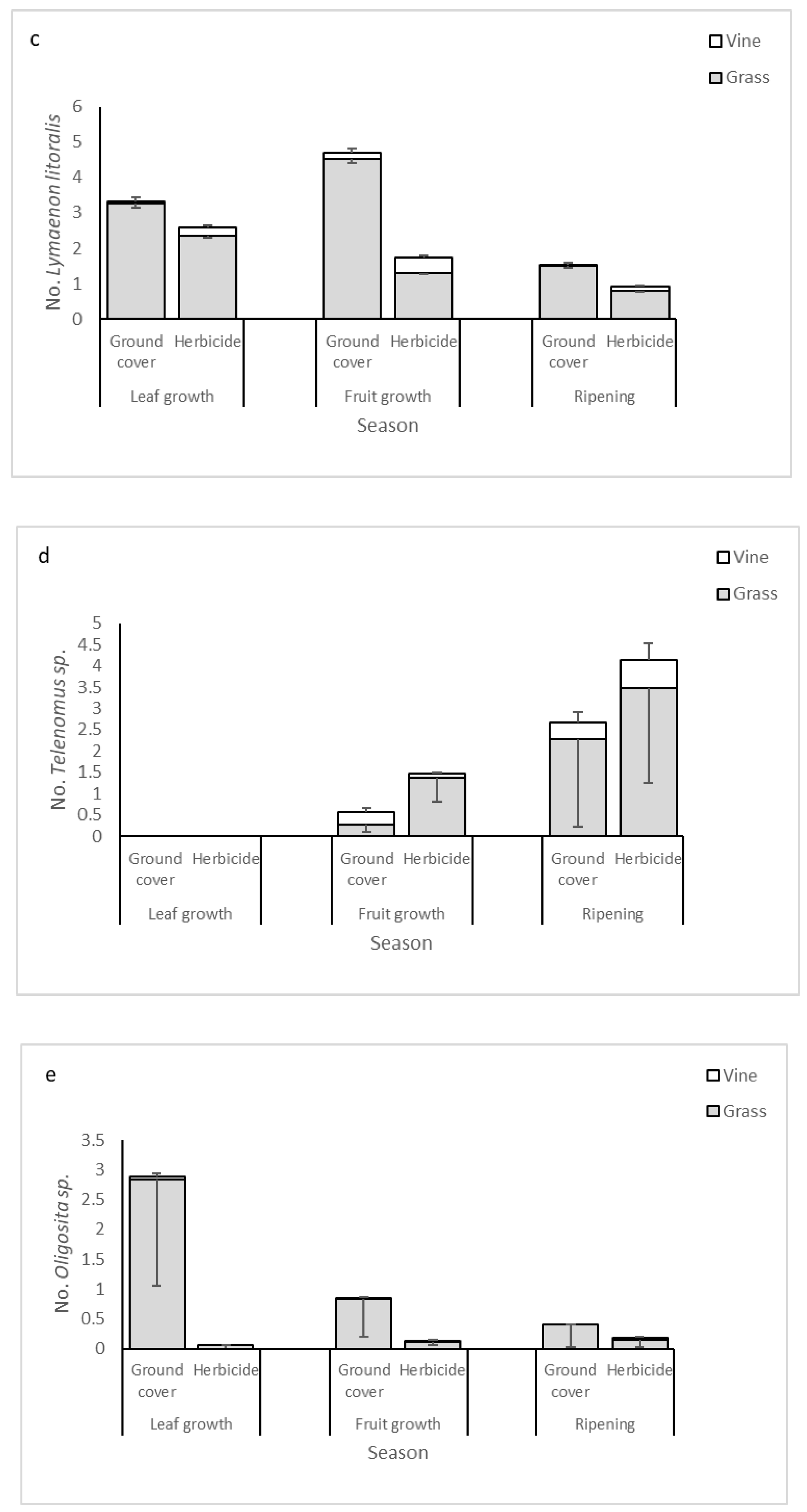
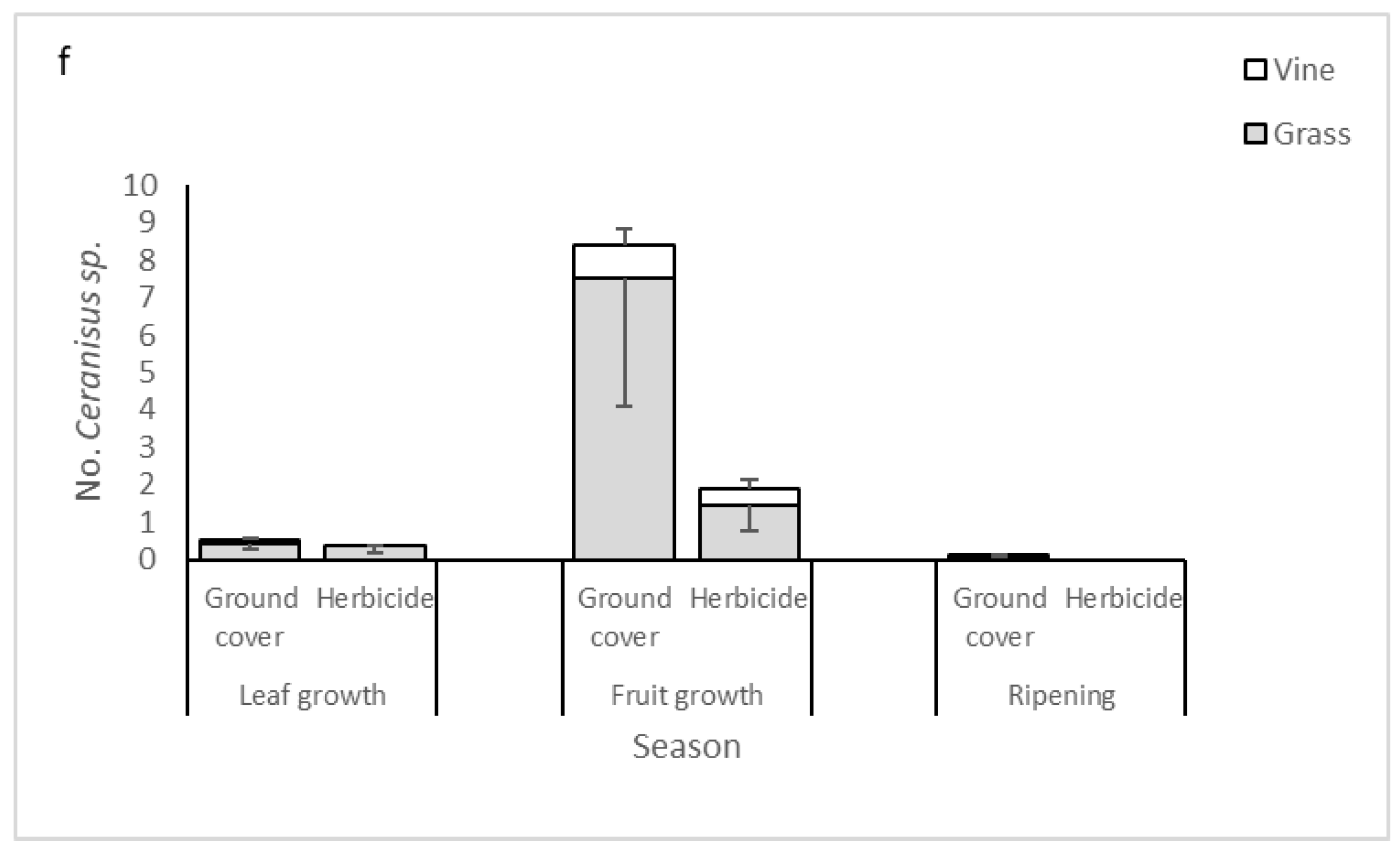
3.4. Dominant Parasitoid Species
3.5. Main Groups of Potential Hosts
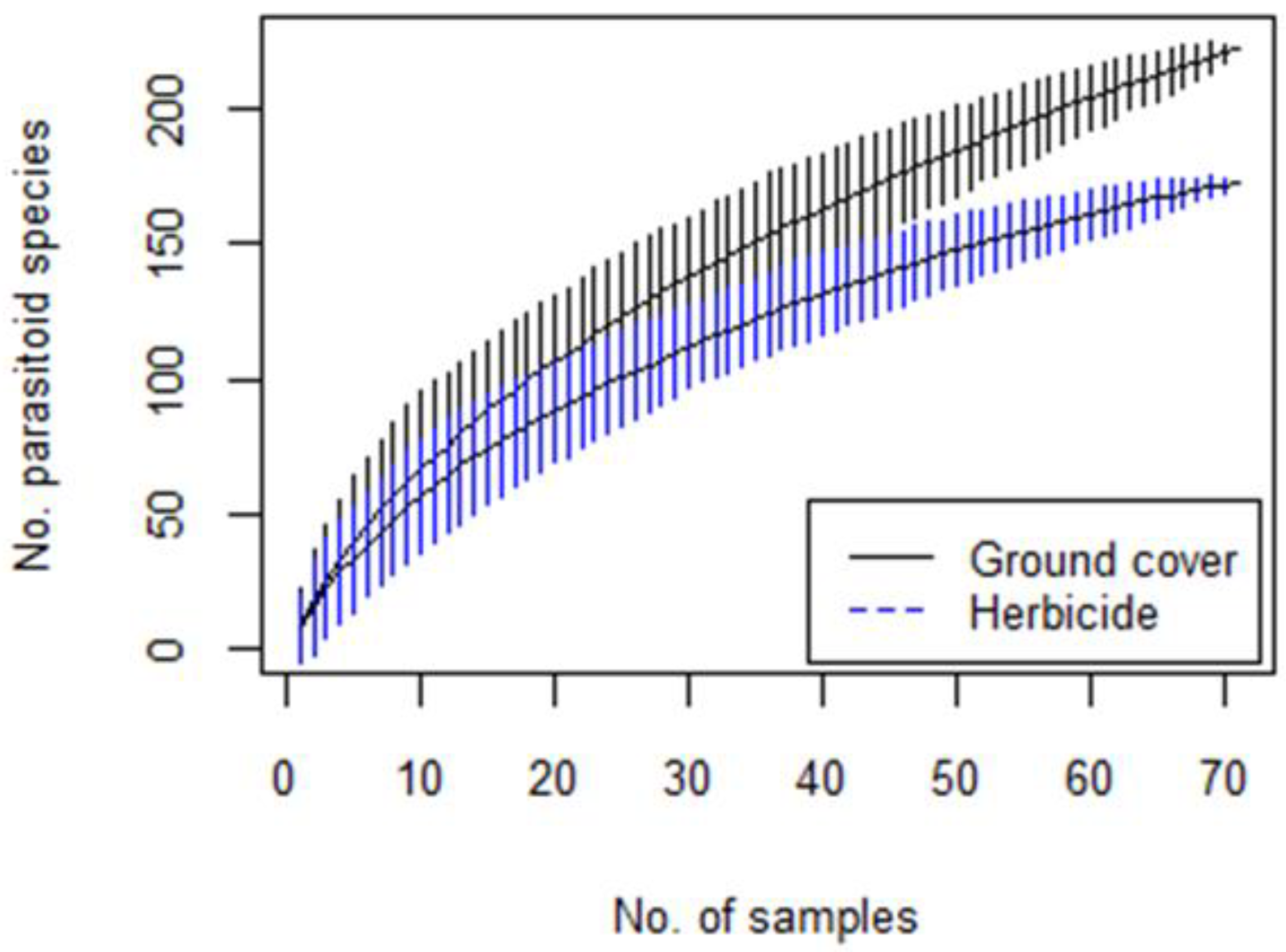
3.6. Parasitoid Species Richness
3.7. Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbosa, P.A. Conservation Biological Control; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Lövei, G.L.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S. A functional overview of conservation biological control. Crop Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Mansion-Vaquié, A.; Ferrante, M.; Cook, S.; Pell, J.; Lövei, G.L. Manipulating field margins to increase predation intensity in fields of winter wheat (Triticum aestivum). J. Appl. Entomol. 2017, 141, 600–611. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, A.K.; Landis, D.A.; Wratten, S.D. Maximizing ecosystem services from conservation biological control: The role of habitat management. Biol. Cont. 2008, 45, 254–271. [Google Scholar] [CrossRef]
- Hassan, K.; Pervin, M.; Mondal, F.; Mala, M. Habitat management: A key option to enhance natural enemies of crop pest. Univ. J. Plant. Sci. 2016, 4, 50–57. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Mailloux, J.; Le Bellec, F.; Kreiter, S.; Tixier, M.; Dubois, P. Influence of ground cover management on diversity and density of phytoseiid mites (Acari: Phytoseiidae) in Guadeloupean citrus orchards. Exp. Appl. Acarol. 2010, 52, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Rand, T.; Bianchi, F. The landscape context of trophic interactions: Insect spillover across the crop-noncrop interface. Ann. Zool. Fenn. 2005, 42, 421–432. [Google Scholar]
- Bianchi, F.J.; Booij, C.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Letourneau, D.K.; Jedlicka, J.A.; Bothwell, S.G.; Moreno, C.R. Effects of natural enemy biodiversity on the suppression of arthropod herbivores in terrestrial ecosystems. Ann. Rev. Ecol. Evol. Syst. 2009, 40, 573–592. [Google Scholar] [CrossRef]
- Dainese, M.; Martin, E.A.; Aizen, M.A.; Albrecht, M.; Bartomeus, I.; Bommarco, R.; Carvalheiro, L.G.; Chaplin-Kramer, R.; Gagic, V.; Garibaldi, L.A. A global synthesis reveals biodiversity-mediated benefits for crop production. Sci. Adv. 2019, 5, eaax0121. [Google Scholar] [CrossRef] [Green Version]
- Meagher, R.; Meyer, J. Influence of ground cover and herbicide treatments on Tetranychus urticae populations in peach orchards. Exp. Appl. Acarol. 1990, 9, 149–158. [Google Scholar] [CrossRef]
- Cahenzli, F.; Sigsgaard, L.; Daniel, C.; Herz, A.; Jamar, L.; Kelderer, M.; Jacobsen, S.K.; Kruczyńska, D.; Matray, S.; Porcel, M. Perennial flower strips for pest control in organic apple orchards—A pan-European study. Agric. Ecosyst. Environ. 2019, 278, 43–53. [Google Scholar] [CrossRef]
- Markó, V.; Jenser, G.; Kondorosy, E.; Ábrahám, L.; Balázs, K. Flowers for better pest control? The effects of apple orchard ground cover management on green apple aphids (Aphis spp.) (Hemiptera: Aphididae), their predators and the canopy insect community. Biocontrol Sci. Technol. 2013, 23, 126–145. [Google Scholar] [CrossRef]
- Paredes, D.; Cayuela, L.; Gurr, G.M.; Campos, M. Is ground cover vegetation an effective biological control enhancement strategy against olive pests? PLoS ONE 2015, 10, e0117265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahl, H.M.; Leslie, A.W.; Hooks, C.R. Effects of red clover living mulch on arthropod herbivores and natural enemies, and cucumber yield. Ann. Entomol. Soc. Am. 2019, 112, 356–364. [Google Scholar] [CrossRef]
- Tschumi, M.; Albrecht, M.; Entling, M.H.; Jacot, K. High effectiveness of tailored flower strips in reducing pests and crop plant damage. Proc. R. Soc. B Biol. Sci. 2015, 282, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Balmer, O.; Pfiffner, L.; Schied, J.; Willareth, M.; Leimgruber, A.; Luka, H.; Traugott, M. Noncrop flowering plants restore top-down herbivore control in agricultural fields. Ecol. Evol. 2013, 3, 2634–2646. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Chang, M.; Tiwari, S.; Sharma, S.; Wratten, S.D. Habitat management for pest management: Limitations and prospects. Ann. Entomol. Soc. Am. 2019, 112, 302–317. [Google Scholar] [CrossRef]
- D’Alberto, C.F.; Hoffmann, A.A.; Thomson, L.J. Limited benefits of non-crop vegetation on spiders in Australian vineyards: Regional or crop differences? Biocontrol 2012, 57, 541–552. [Google Scholar] [CrossRef]
- Baggen, L.; Gurr, G.; Meats, A. Flowers in tri-trophic systems: Mechanisms allowing selective exploitation by insect natural enemies for conservation biological control. Entomol. Exp. Appl. 1999, 91, 155–161. [Google Scholar] [CrossRef]
- Paiola, A.; Assandri, G.; Brambilla, M.; Zottini, M.; Pedrini, P.; Nascimbene, J. Exploring the potential of vineyards for biodiversity conservation and delivery of biodiversity-mediated ecosystem services: A global-scale systematic review. Sci. Total Environ. 2020, 706, 135839. [Google Scholar] [CrossRef] [PubMed]
- Altieri, M.A.; Ponti, L.; Nicholls, C.I. Manipulating vineyard biodiversity for improved insect pest management: Case studies from northern California. Int. J. Biodivers. Sci. Manag. 2005, 1, 191–203. [Google Scholar] [CrossRef]
- Thomson, L.J.; Hoffmann, A.A. Vegetation increases the abundance of natural enemies in vineyards. Biol. Cont. 2009, 49, 259–269. [Google Scholar] [CrossRef]
- Danne, A.; Thomson, L.; Sharley, D.; Penfold, C.; Hoffmann, A. Effects of native grass cover crops on beneficial and pest invertebrates in Australian vineyards. Environ. Entomol. 2010, 39, 970–978. [Google Scholar] [CrossRef] [PubMed]
- English-Loeb, G.; Rhainds, M.; Martinson, T.; Ugine, T. Influence of flowering cover crops on Anagrus parasitoids (Hymenoptera: Mymaridae) and Erythroneura leafhoppers (Homoptera: Cicadellidae) in New York vineyards. Agric. For. Entomol. 2003, 5, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Burgio, G.; Marchesini, E.; Reggiani, N.; Montepaone, G.; Schiatti, P.; Sommaggio, D. Habitat management of organic vineyard in Northern Italy: The role of cover plants management on arthropod functional biodiversity. Bull. Entomol. Res. 2016, 106, 759–768. [Google Scholar] [CrossRef]
- Shapira, I.; Rosenfeld, A.; Rothschild, A.; Ackerman, M.; Eshel, G.; Keasar, T. Herbaceous vegetation enhancement increases biodiversity in a wine-producing vineyard in Israel, promoting shifts in agricultural practices in other vineyards. Conserv. Evid. 2017, 14, 10–15. [Google Scholar]
- Daane, K.M.; Hogg, B.N.; Wilson, H.; Yokota, G.Y. Native grass ground covers provide multiple ecosystem services in Californian vineyards. J. Appl. Ecol. 2018, 55, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Perez, R.; Gloria Saenz-Romo, M.; Vicente-Diez, I.; Ibanez-Pascual, S.; Martinez-Villar, E.; Santiago Marco-Mancebon, V.; Perez-Moreno, I.; Campos-Herrera, R. Impact of vineyard ground cover management on the occurrence and activity of entomopathogenic nematodes and associated soil organisms. Agric. Ecosyst. Environ. 2020, 301, 107028. [Google Scholar] [CrossRef]
- Olmstead, M.; James, D. Ecosystem approaches to managing insect and mite populations effectively in inland desert vineyards utilizing sustainable production practices—A review. J. Am. Pomolog. Soc. 2014, 68, 97–110. [Google Scholar]
- Kidd, N.A.; Jervis, M.A. Population dynamics. In Insects as Natural Enemies; Springer: Dordrecht, The Netherlands, 2007; pp. 435–523. [Google Scholar]
- Hajek, A.E.; Eilenberg, J. Natural Enemies: An. Introduction to Biological Control; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Kishinevsky, M.; Keasar, T.; Bar-Massada, A. Parasitoid abundance on plants: Effects of host abundance, plant species, and plant flowering state. Arthropod Plant Interact. 2017, 11, 155–161. [Google Scholar] [CrossRef]
- Kishinevsky, M.; Cohen, N.; Chiel, E.; Wajnberg, E.; Keasar, T. Sugar feeding of parasitoids in an agroecosystem: Effects of community composition, habitat and vegetation. Insect Conserv. Divers. 2018, 11, 50–57. [Google Scholar] [CrossRef]
- Masner, L. Key to genera of Scelionidae of the Holarctic region, with descriptions of new genera and species (Hymenoptera: Proctotrupoidea). Mem. Entomol. Soc. Can. 1980, 112, 54. [Google Scholar] [CrossRef] [Green Version]
- Goulet, H.; Huber, J.T. Hymenoptera of the World: An Identification Guide to Families; Agriculture Canada Publication: Ottawa, ON, Canada, 1993. [Google Scholar]
- Gibson, G.A.; Huber, J.T.; Woolley, J.B. Annotated Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera); NRC Research Press: Ottawa, ON, Canada, 1997. [Google Scholar]
- Pinto, J.D. A review of the New World genera of Trichogrammatidae (Hymenoptera). J. Hymenopt. Res. 2006, 15, 38–163. [Google Scholar]
- Huber, J.T.; Viggiani, G.; Jesu, R. Order Hymenoptera, family Mymaridae. Arthropod Fauna UAE 2009, 2, 270–297. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Martinez Arbizu, P. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis; R Package Version 0.0; Volume 1. 2017. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 18 December 2020).
- Lemon, J. Plotrix: A package in the red light district of R. R-News 2006, 6, 8–12. [Google Scholar]
- Meyer, D.; Zeileis, A.; Hornik, K. Vcd: Visualizing Categorical Data. Available online: https://cran.r-project.org/web/packages/vcd/ (accessed on 18 December 2020).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package. R Package Version 2.4-3; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Yee, T.W.; Stoklosa, J.; Huggins, R.M. The VGAM package for capture-recapture data using the conditional likelihood. J. Stat. Softw. 2015, 65, 3475. [Google Scholar] [CrossRef] [Green Version]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Stephens, C.; Schellhorn, N.; Wood, G.; Austin, A. Parasitic wasp assemblages associated with native and weedy plant species in an agricultural landscape. Aust. J. Entomol. 2006, 45, 176–184. [Google Scholar] [CrossRef]
- Dainese, M.; Schneider, G.; Krauss, J.; Steffan-Dewenter, I. Complementarity among natural enemies enhances pest suppression. Sci. Rep. 2017, 7, 8172. [Google Scholar] [CrossRef] [PubMed]
- Petanidou, T.; Kallimanis, A.S.; Tzanopoulos, J.; Sgardelis, S.P.; Pantis, J.D. Long-term observation of a pollination network: Fluctuation in species and interactions, relative invariance of network structure and implications for estimates of specialization. Ecol. Lett. 2008, 11, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.M. Complex long-term dynamics of pollinator abundance in undisturbed Mediterranean montane habitats over two decades. Ecol. Monogr. 2019, 89, e01338. [Google Scholar] [CrossRef] [Green Version]
- Segoli, M.; Kishinevsky, M.; Rozenberg, T.; Hoffmann, I. Parasitoid abundance and community composition in desert vineyards and their adjacent natural habitats. Insects 2020, 11, 580. [Google Scholar] [CrossRef]
- Shapira, I.; Gavish-Regev, E.; Sharon, R.; Harari, A.R.; Kishinevsky, M.; Keasar, T. Habitat use by crop pests and natural enemies in a Mediterranean vineyard agroecosystem. Agric. Ecosyst. Environ. 2018, 267, 109–118. [Google Scholar] [CrossRef]
- Rand, T.; Tylianakis, J.; Tscharntke, T. Spillover edge effects: The dispersal of agriculturally subsidized insect natural enemies into adjacent natural habitats. Ecol. Lett. 2006, 9, 603–614. [Google Scholar] [CrossRef]
- Teschner, N.; Orenstein, D.E.; Shapira, I.; Keasar, T. Socio-ecological research and the transition toward sustainable agriculture. Int. J. Agric. Sustain. 2017, 15, 99–101. [Google Scholar] [CrossRef]
- Rodrigo-Comino, J. Five decades of soil erosion research in “terroir”. The State-of-the-Art. Earth Sci. Rev. 2018, 179, 436–447. [Google Scholar] [CrossRef]
- López-Piñeiro, A.; Muñoz, A.; Zamora, E.; Ramírez, M. Influence of the management regime and phenological state of the vines on the physicochemical properties and the seasonal fluctuations of the microorganisms in a vineyard soil under semi-arid conditions. Soil Tillage Res. 2013, 126, 119–126. [Google Scholar] [CrossRef]
- Peregrina, F.; Pilar Perez-Alvarez, E.; Colina, M.; Garcia-Escudero, E. Cover crops and tillage influence soil organic matter and nitrogen availability in a semi-arid vineyard. Arch. Agron. Soil Sci. 2012, 58, S95–S102. [Google Scholar] [CrossRef]
- Monteiro, A.; Lopes, C.M. Influence of cover crop on water use and performance of vineyard in Mediterranean Portugal. Agric. Ecosyst. Environ. 2007, 121, 336–342. [Google Scholar] [CrossRef]
- Foley, J.A.; Defries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Wanger, T.C.; DeClerck, F.; Garibaldi, L.A.; Ghazoul, J.; Kleijn, D.; Klein, A.; Kremen, C.; Mooney, H.; Perfecto, I.; Powell, L.L. Integrating agroecological production in a robust post-2020 Global Biodiversity Framework. Nat. Ecol. Evol. 2020, 4, 1150–1152. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.; Collen, B. Defaunation in the anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explanatory Variables | df | LRT | p |
|---|---|---|---|
| Parasitoids | |||
| Management | 1 | 4.04 | 0.044 |
| Habitat | 1 | 36.23 | <0.001 |
| Year | 2 | 7.43 | 0.024 |
| Lymaenon litoralis | |||
| Management | 1 | 4.73 | 0.030 |
| Season | 1 | 7.95 | 0.019 |
| Habitat | 1 | 34.62 | <0.001 |
| Host: Auchenorrhyncha | 1 | 12.54 | <0.001 |
| Management × Habitat | 1 | 4.87 | 0.027 |
| Ceranisus sp. | |||
| Management | 1 | 3.72 | 0.054 |
| Season | 2 | 24.93 | <0.001 |
| Habitat | 1 | 4.66 | 0.031 |
| Host: Thysanoptera | 1 | 47.93 | <0.001 |
| Telenomus sp. | |||
| Management | 1 | 3.47 | 0.063 |
| Season | 2 | 21.30 | <0.001 |
| Habitat | 1 | 3.36 | 0.067 |
| Management × Year | 2 | 5.94 | 0.051 |
| Anagrus sp. | |||
| Season | 2 | 7.67 | 0.021 |
| Year | 2 | 11.23 | 0.003 |
| Host: Auchenorrhyncha | 1 | 33.13 | <0.001 |
| Oligosita sp. | |||
| Management | 1 | 3.72 | 0.054 |
| Habitat | 1 | 13.68 | <0.001 |
| Year | 2 | 5.30 | 0.071 |
| Host: Auchenorrhyncha | 1 | 8.44 | 0.004 |
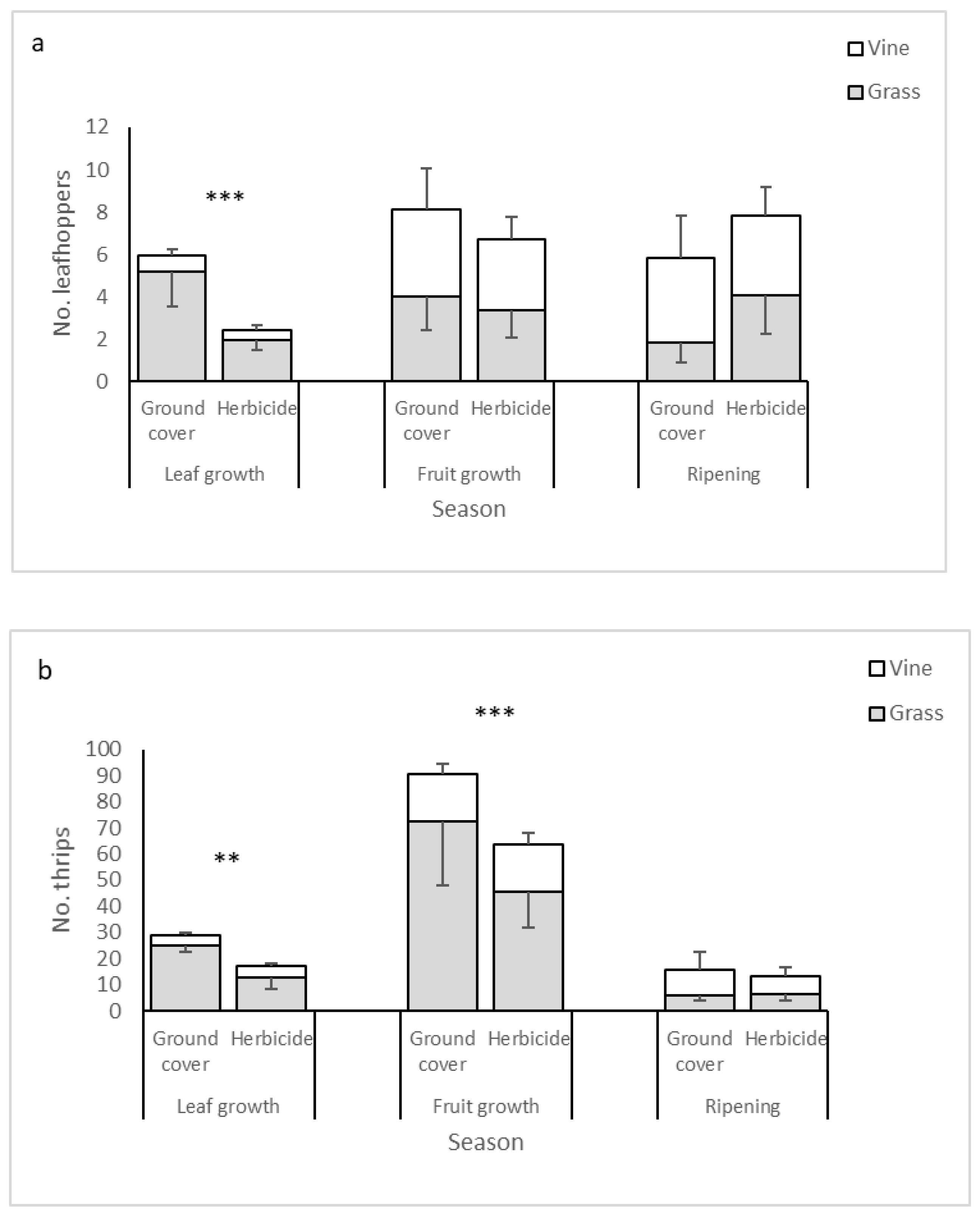
| Leafhoppers | |||
| Management | 1 | 9.31 | 0.002 |
| Year | 2 | 88.76 | <0.001 |
| Management × Season | 2 | 6.78 | 0.03 |
| Thrips | |||
| Management | 1 | 59.33 | <0.001 |
| Season | 2 | 1182.80 | <0.001 |
| Habitat | 1 | 4430.10 | <0.001 |
| Year | 2 | 391.46 | <0.001 |
| Management × Habitat | 1 | 25.77 | <0.001 |
| Management × Season | 2 | 10.30 | 0.006 |
| Df | R2 | F | p | |
|---|---|---|---|---|
| Sampling season | 2 | 0.091 | 6.334 | 0.001 |
| Year | 2 | 0.056 | 3.925 | 0.001 |
| Habitat | 1 | 0.052 | 7.221 | 0.001 |
| Management | 1 | 0.017 | 2.386 | 0.015 |
| Management × Habitat | 1 | 0.018 | 2.528 | 0.016 |
| Residuals | 77 | 0.552 | ||
| Total | 109 | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Möller, G.; Keasar, T.; Shapira, I.; Möller, D.; Ferrante, M.; Segoli, M. Effect of Weed Management on the Parasitoid Community in Mediterranean Vineyards. Biology 2021, 10, 7. https://doi.org/10.3390/biology10010007
Möller G, Keasar T, Shapira I, Möller D, Ferrante M, Segoli M. Effect of Weed Management on the Parasitoid Community in Mediterranean Vineyards. Biology. 2021; 10(1):7. https://doi.org/10.3390/biology10010007
Chicago/Turabian StyleMöller, Gabriella, Tamar Keasar, Idan Shapira, Daniella Möller, Marco Ferrante, and Michal Segoli. 2021. "Effect of Weed Management on the Parasitoid Community in Mediterranean Vineyards" Biology 10, no. 1: 7. https://doi.org/10.3390/biology10010007