

Evaluating the Fungal Pathogens’ Inhibition Efficiency of Composite Film Combined with Antagonistic Yeasts and Sodium Alginate on Peach
Abstract
:1. Introduction
2. Materials and Methods
2.1. Peach Sampling and Cultivation of Microorganisms
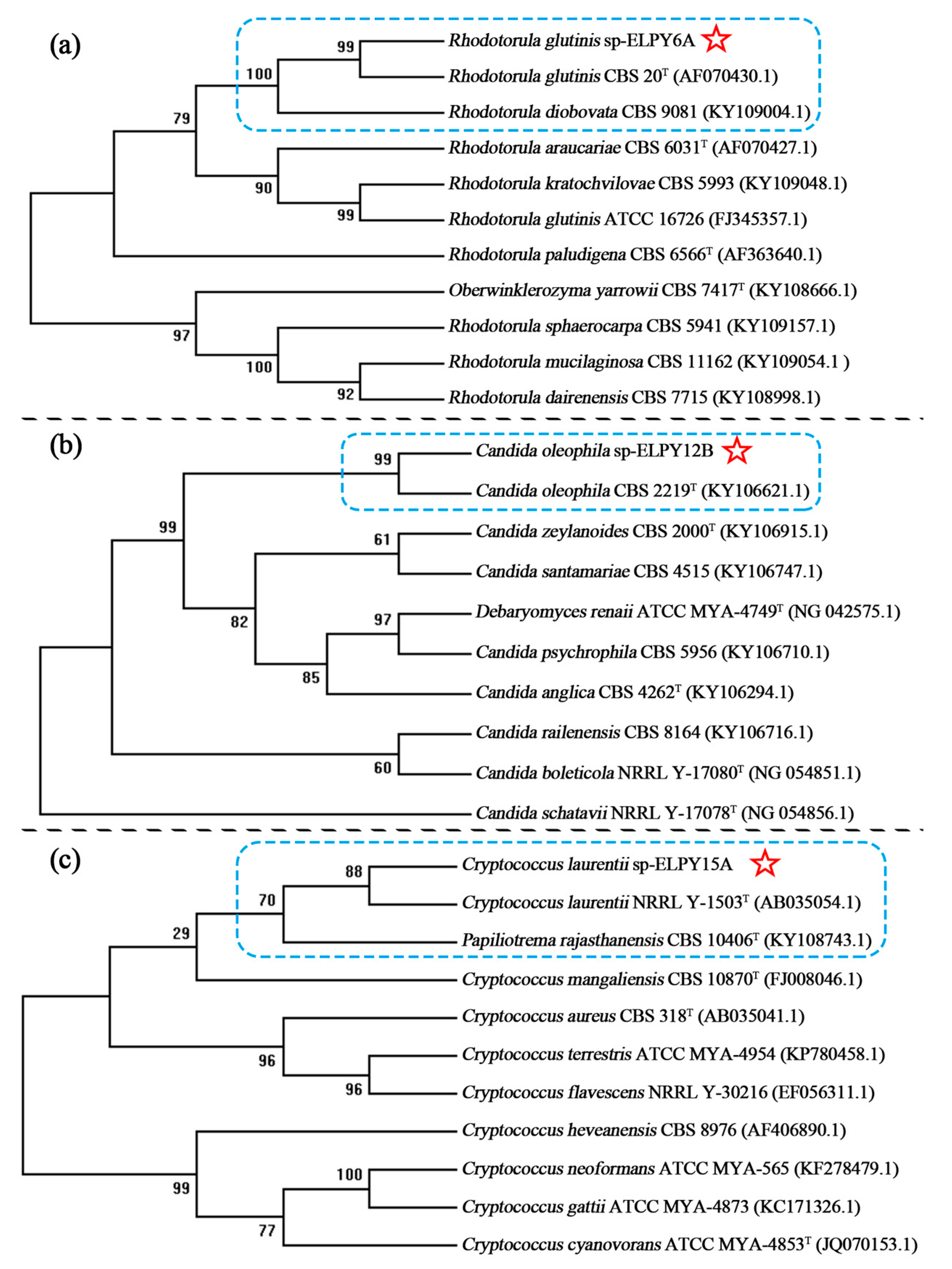
2.2. Molecular Identification of Antagonistic Yeasts
2.3. In Vitro Antifungal Test of Antagonistic Yeasts
2.4. Colonizing Capacity of Candidate Antagonistic Yeasts
2.5. Antagonistic Yeast Preservative Effect Analysis on Fungi-Inoculated Peach
2.6. Evaluating the Fungal Pathogens’ Inhibition Efficiency for Composite Films
2.7. Statistical Data Processing
3. Results
3.1. Isolation and Identification of Yeasts on Peaches
3.2. In Vitro Antifungal Test for Screening the Isolated Yeasts
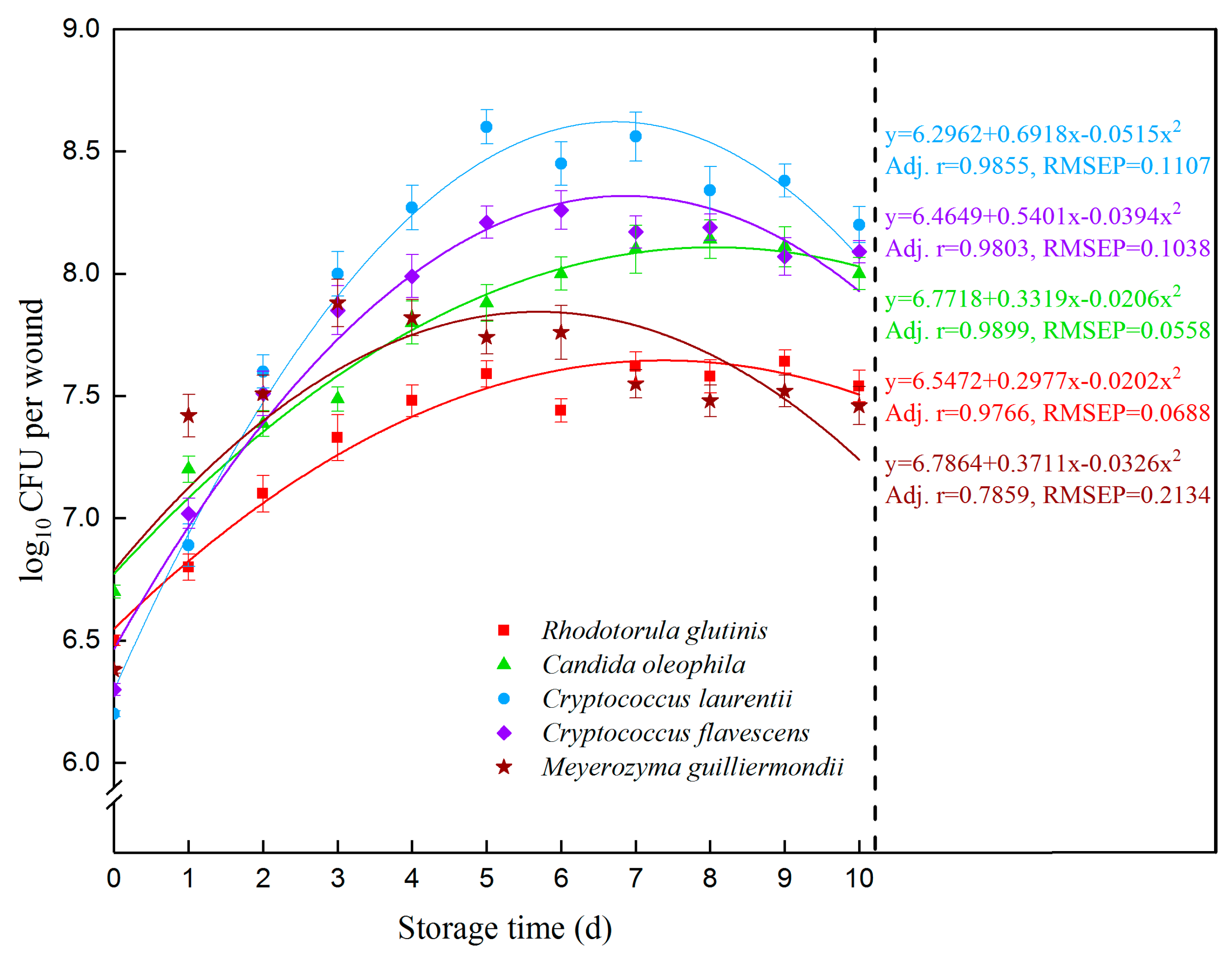
3.3. Colonizing Capacity of Candidate Antagonistic Yeasts on Peaches
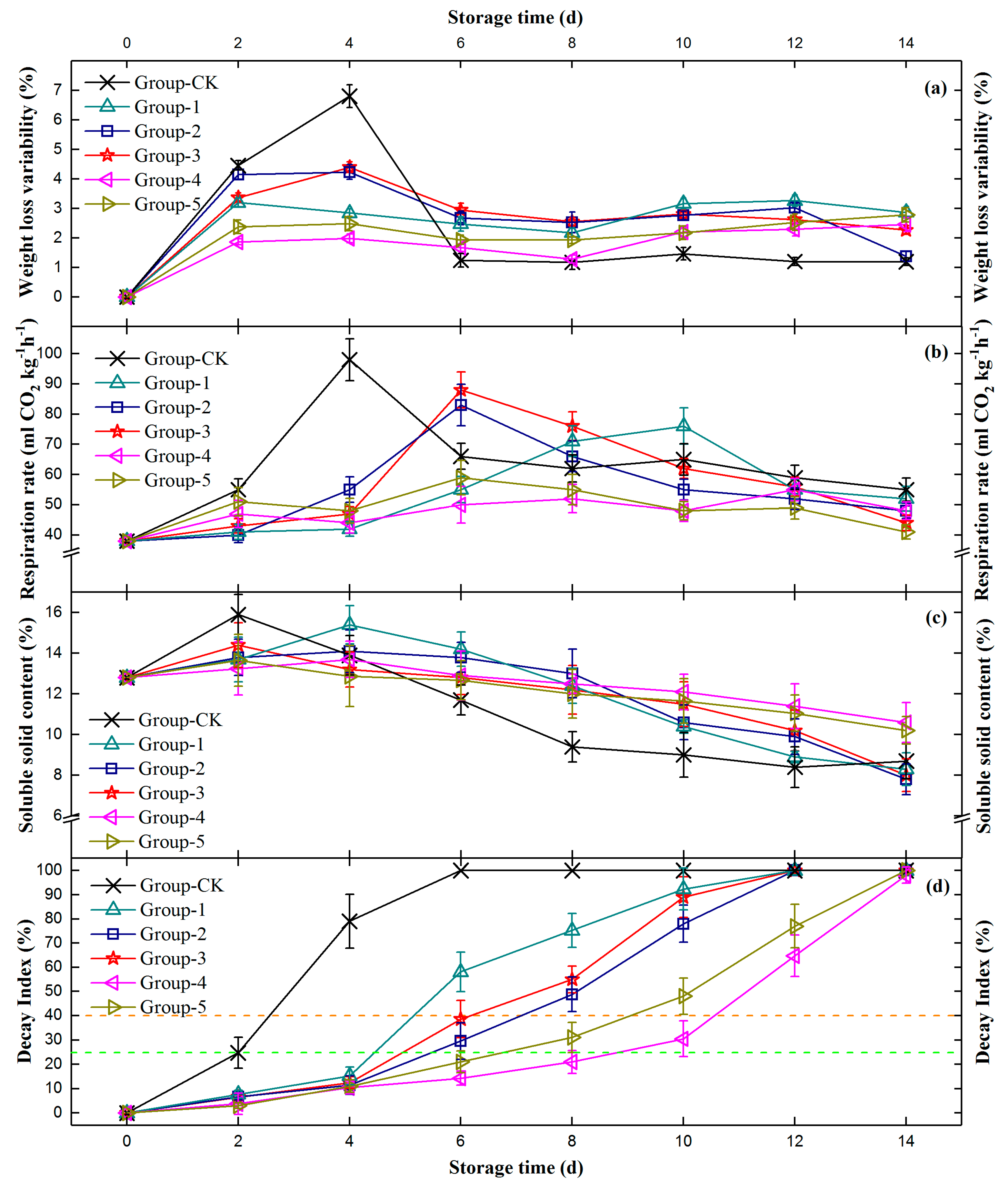
3.4. In Vivo Preservative Effects of Candidate Antagonistic Yeasts on Fungi-Inoculated Peaches
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Usall, J.; Torres, R.; Teixid, N. Biological Control of Postharvest Diseases on Fruit: A Suitable Alternative? Curr. Opin. Food Sci. 2016, 11, 51–55. [Google Scholar] [CrossRef]
- Bhat, R.; Stamminger, R. Preserving Strawberry Quality by Employing Novel Food Preservation and Processing Techniques—Recent Updates and Future Scope—An Overview. J. Food Process Eng. 2015, 38, 536–554. [Google Scholar] [CrossRef]
- Oliveira Filho, J.G.d.; Silva, G.d.C.; Oldoni, F.C.A.; Miranda, M.; Florencio, C.; Oliveira, R.M.D.d.; Gomes, M.d.P.; Ferreira, M.D. Edible Coating Based on Carnauba Wax Nanoemulsion and Cymbopogon martinii Essential Oil on Papaya Postharvest Preservation. Coatings 2022, 12, 1700. [Google Scholar] [CrossRef]
- Embuscado, M.E.; Huber, K.C. Edible Films and Coatings for Food Applications; Springer: New York, NY, USA, 2008. [Google Scholar]
- Xiao, Q.; Lu, K.; Tong, Q.; Liu, C. Barrier Properties and Microstructure of Pullulan–Alginate-Based Films. J. Food Process Eng. 2015, 38, 155–161. [Google Scholar] [CrossRef]
- Du, X.L.; Li, H.; Zhou, W.H.; Liu, Y.; Li, J.L. Determination of Quality Changes in Peaches Wrapped in Active Paper and Stored at Ambient Temperature in Summer. Sci. Rep. 2017, 7, 11830. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. PCR Protocols; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions Through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Atsamnia, D.; Hamadache, M.; Hanini, S.; Benkortbi, O.; Oukrif, D. Prediction of the Antibacterial Activity of Garlic Extract on E. coli, S. aureus and B. subtilis by Determining the Diameter of the Inhibition Zones Using Artificial Neural Networks. LWT-Food Sci. Technol. 2017, 82, 287–295. [Google Scholar] [CrossRef]
- Zhang, D.P.; Lu, C.G.; Zhang, T.T.; Spadaro, D.; Liu, D.W.; Liu, W.C. Candida pruni sp. nov. is a New Yeast Species with Antagonistic Potential Against Brown Rot of Peaches. Arch. Microbiol. 2014, 196, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Pavli, F.; Kovaiou, I.; Apostolakopoulou, G.; Kapetanakou, A.; Skandamis, P.; Nychas, G.J.; Tassou, C.; Chorianopoulos, N. Alginate-Based Edible Films Delivering Probiotic Bacteria to Sliced Ham Pretreated with High Pressure Processing. Int. J. Mol. Sci. 2017, 18, 1867. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Identification and Phylogeny of Ascomycetous Yeasts from Analysis of Nuclear Large Subunit (26S) Ribosomal DNA Partial Sequences. Anton. Leeuw. Int. J. G. 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Buzzini, P.; Lachance, M.A.; Yurkov, A. (Eds.) Yeasts in Natural Ecosystems: Diversity; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Aguilar-Uscanga, B.; Francois, J.M. A Study of the Yeast Cell Wall Composition and Structure in response to growth Conditions and Mode of Cultivation. Lett. Appl. Microbiol. 2003, 37, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.H. Stress Biology of Yeasts and Fungi: Applications for Industrial Brewing and Fermentation; Springer: Tokyo, Japan, 2015. [Google Scholar]
- Picha, D.H. Weight Loss in Sweet Potatoes During Curing and Storage: Contribution of Transpiration and Respiration. J. Am. Soc. Hortic. 1986, 111, 889–892. [Google Scholar] [CrossRef]
- Bautista-Banos, S. Postharvest Decay: Control Strategies; Elsevier Inc.: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Watkins*, C.; Miller, W.B. A Summary of Physiological Processes or Disorders in Fruits, Vegetables and Ornamental Products that are Delayed or Decreased, Increased, or Unaffected by Application of 1-Methylcyclopropene (1-MCP). HortScience 2004, 39, 816. [Google Scholar] [CrossRef]
- Yang, Z.; Cao, S.; Su, X.; Jiang, Y. Respiratory Activity and Mitochondrial Membrane Associated with Fruit Senescence in Postharvest Peaches in Response to UV-C Treatment. Food Chem. 2014, 161, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Zhang, B.; Ye, Z.; Chen, K.; Guo, J.; Gu, X.; Shen, J. Pulp volatiles measured by An Electronic Nose Are Related to Harvest Season, T.S.S. Concentration and TSS/TA Ratio Among 39 Peaches and Nectarines. Sci. Hortic. 2013, 150, 146–153. [Google Scholar] [CrossRef]
- Gatto, M.A.; Ippolito, A.; Linsalata, V.; Cascarano, N.A.; Nigro, F.; Vanadia, S.; Di Venere, D. Activity of Extracts From Wild Edible Herbs Against Postharvest Fungal Diseases of Fruit and Vegetables. Postharvest Biol. Technol. 2011, 61, 72–82. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group Name | Treatments | Instructions |
|---|---|---|
| Control Check (CK) | Sterile Water |
|
| Group 1 | Na-alginate film |
|
| Group 2 | Cryptococcus laurentii |
|
| Group 3 | Candida oleophila | |
| Group 4 | Na-alginate film composite by Cryptococcus laurentii |
|
| Group 5 | Na-alginate film composite by Candida oleophila |
| No. | The Tested Items | The Diameter of the Inhibition Zone (mm) | Total Relative Inhibitory Rate (%) | |||||
|---|---|---|---|---|---|---|---|---|
| A. tubingensis | P. expansum | B. elliptica | R. stolonifer | A. alternate | M. fructicola | |||
| 1 | Sterile Water (dnc) | 6.00 ± 0.00 g | 6.00 ± 0.00 f | 6.00 ± 0.00 f | 6.00 ± 0.00 g | 6.00 ± 0.00 e | 6.00 ± 0.00 g | N/A+ |
| 2 | Amphotericin B (dpc) | 20.53 ± 2.12 a | 18.76 ± 1.56 a | 19.88 ± 1.44 a | 18.94 ± 1.76 a | 19.07 ± 2.11 a | 19.85 ± 2.69 a | N/A |
| 3 | Cryptococcus laurentii | 12.73 ± 1.25 bcd | 16.38 ± 1.37 ab | 15.04 ± 1.66 b | 11.15 ± 1.16 def | 14.42 ± 1.58 b | 14.93 ± 1.36 b | 60.04 |
| 4 | Candida oleophila | 13.88 ± 1.43 b | 14.62 ± 2.59 c | 12.97 ± 1.79 bcde | 12.64 ± 1.52 bc | 12.58 ± 2.39 bc | 13.35 ± 1.48 b | 54.35 |
| 5 | Meyerozyma guilliermondii | 12.48 ± 0.85 bcd | 13.39 ± 1.93 c | 13.25 ± 1.64 bcd | 14.23 ± 0.68 b | 12.36 ± 2.85 bc | 10.70 ± 1.14 bcd | 49.87 |
| 6 | Sporobolomyces roseus | 11.52 ± 1.29 cd | 13.83 ± 2.04 bc | 13.75 ± 2.15 bc | 14.22 ± 1.77 b | 13.11 ± 2.53 bc | 9.54 ± 1.02 def | 49.32 |
| 7 | Metschnikowia citriensis | 13.65 ± 2.01 b | 13.96 ± 1.83 bc | 10.63 ± 1.00 de | 12.94 ± 2.21 bc | 13.75 ± 2.22 bc | 8.58 ± 1.31 defg | 46.29 |
| 8 | Rhodotorula glutinis | 10.63 ± 1.13 de | 11.96 ± 1.38 cd | 13.67 ± 1.68 bc | 13.56 ± 1.67 bc | 12.18 ± 1.37 bc | 10.97 ± 2.01 bcd | 45.62 |
| 9 | Cryptococcus flavescens | 10.59 ± 0.96 cd | 12.77 ± 1.76 c | 11.86 ± 1.47 cde | 12.02 ± 1.73 bcd | 11.35 ± 1.33 bc | 12.54 ± 2.37 bc | 43.35 |
| 10 | Metschnikowia zizyphicola | 12.74 ± 1.48 bc | 14.11 ± 1.65 bc | 11.42 ± 1.59 cde | 13.07 ± 0.91 bc | 12.24 ± 2.55 bc | 7.35 ± 0.82 fg | 43.11 |
| 11 | Pichia anomala | 8.24 ± 0.76 f | 12.42 ± 1.79 c | 10.27 ± 1.06 e | 12.51 ± 1.16 bc | 7.63 ± 1.19 de | 13.46 ± 1.39 b | 35.21 |
| 12 | Candida glabrata | 9.11 ± 0.47 ef | 9.20 ± 1.06 e | 11.89 ± 1.79 cde | 11.34 ± 0.88 cde | 10.61 ± 0.95 cd | 8.34 ± 0.73 efg | 30.22 |
| 13 | Candida inconspicua | 7.86 ± 0.43 fg | 8.78 ± 0.95 e | 10.56 ± 2.35 de | 8.29 ± 1.26 f | 11.57 ± 1.35 bc | 7.43 ± 0.66 fg | 22.82 |
| 14 | Pichia fermentans | 6.00 ± 0.00 g | 9.47 ± 0.84 de | 10.56 ± 1.75 de | 9.68 ± 1.14 ef | 7.93 ± 0.82 de | 10.34 ± 1.39 cde | 22.19 |
| 15 | Torulaspora delbrueckii | 6.00 ± 0.00 g | 6.00 ± 0.00 f | 7.49 ± 0.59 f | 6.00 ± 0.00 g | 12.43 ± 2.77 bc | 8.82 ± 0.98 def | 13.25 |
| 16 | Clavispora lusitaniae | 6.00 ± 0.00 g | 7.36 ± 0.99 ef | 6.00 ± 0.00 f | 9.81 ± 0.77 def | 6.00 ± 0.00 e | 7.59 ± 1.17 fg | 8.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.; Li, S.; Luo, A.; Yin, X.; Fan, K.; Mou, L.; Li, J. Evaluating the Fungal Pathogens’ Inhibition Efficiency of Composite Film Combined with Antagonistic Yeasts and Sodium Alginate on Peach. Coatings 2023, 13, 417. https://doi.org/10.3390/coatings13020417
Du X, Li S, Luo A, Yin X, Fan K, Mou L, Li J. Evaluating the Fungal Pathogens’ Inhibition Efficiency of Composite Film Combined with Antagonistic Yeasts and Sodium Alginate on Peach. Coatings. 2023; 13(2):417. https://doi.org/10.3390/coatings13020417
Chicago/Turabian StyleDu, Xiaolong, Shaobin Li, An Luo, Xiaoli Yin, Kai Fan, Linyun Mou, and Jianlong Li. 2023. "Evaluating the Fungal Pathogens’ Inhibition Efficiency of Composite Film Combined with Antagonistic Yeasts and Sodium Alginate on Peach" Coatings 13, no. 2: 417. https://doi.org/10.3390/coatings13020417