Stress-Based High-Throughput Screening Assays to Identify Inhibitors of Cell Envelope Biogenesis


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Materials, Reagents and Enzymes
2.3. Plasmid Construction
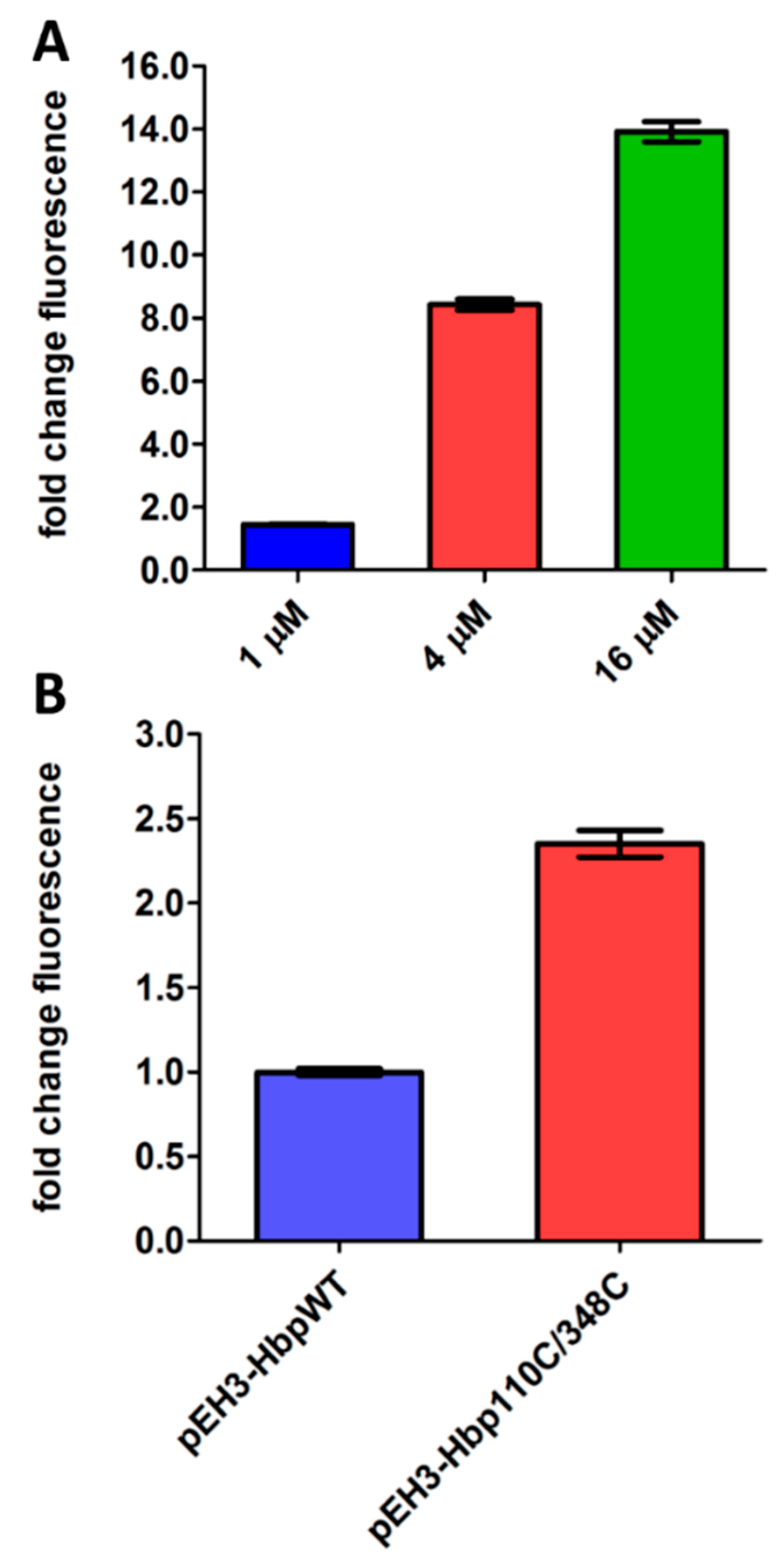
2.4. DjlA and Hbp Expression
2.5. Susceptibility to Antibiotics and Stress Activation
3. Results
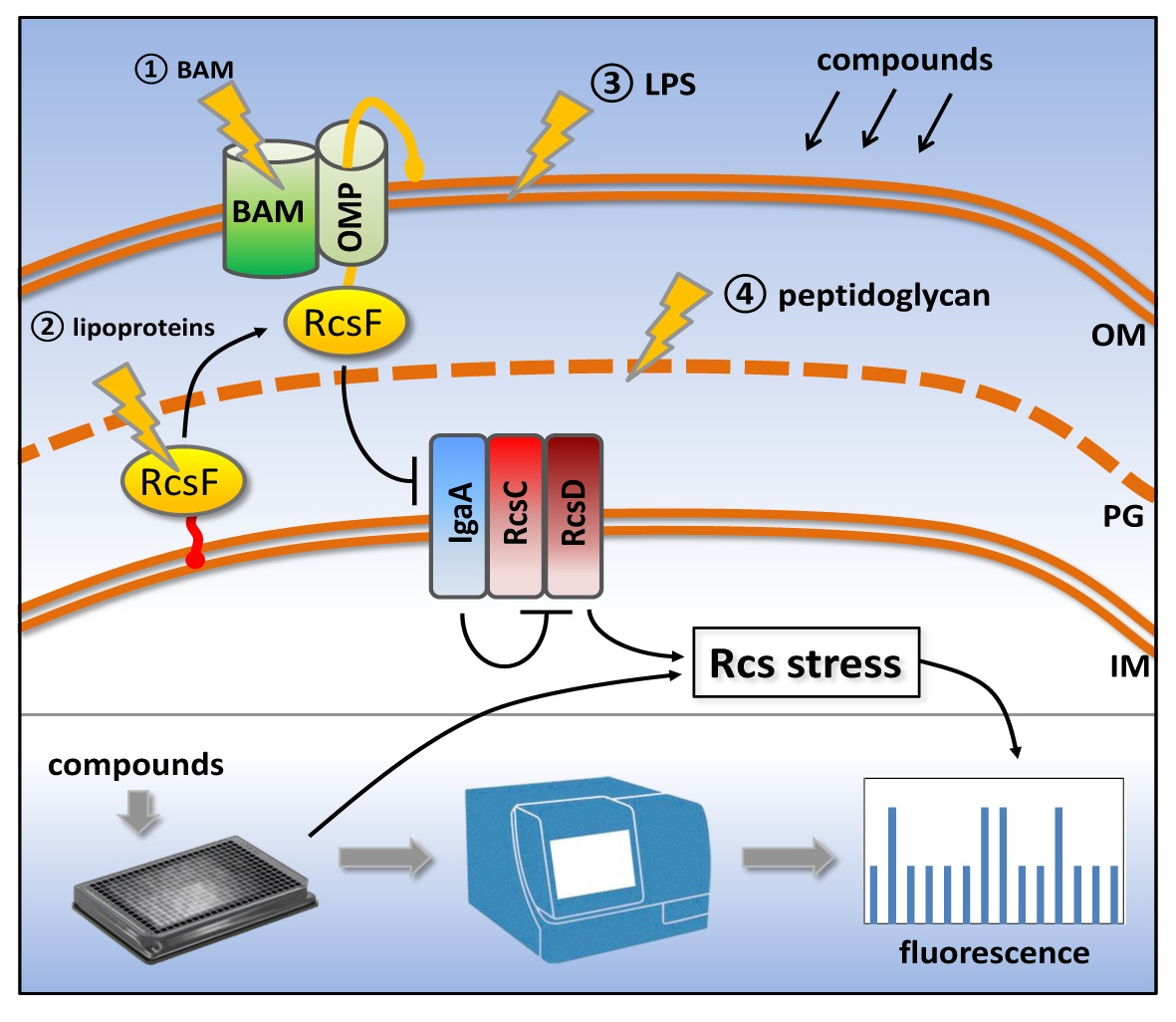
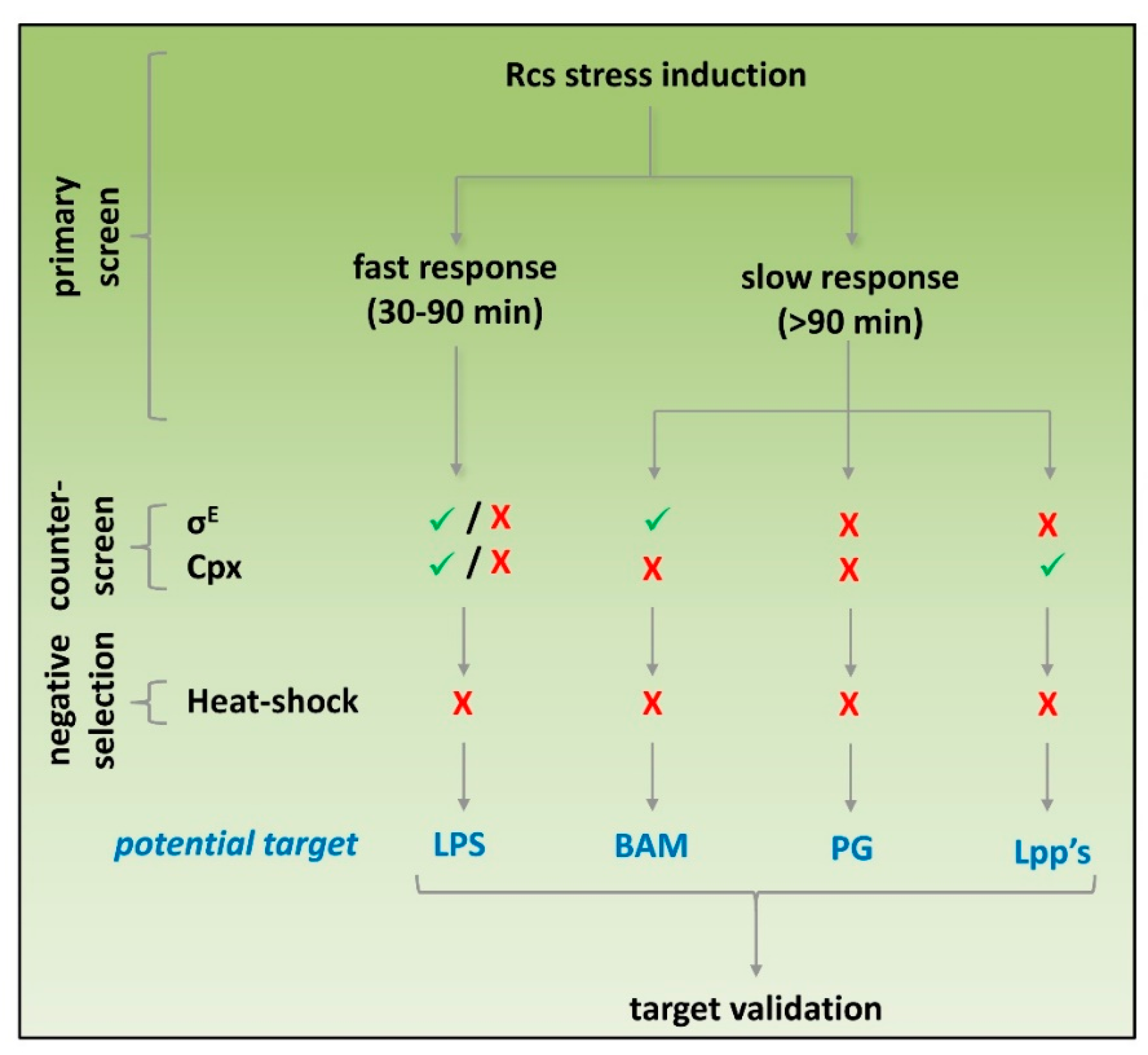
3.1. Development of Rcs and Cpx Stress Reporter Assays
3.2. Compromised LPS Integrity Induces Rcs and Cpx Stress Reporters
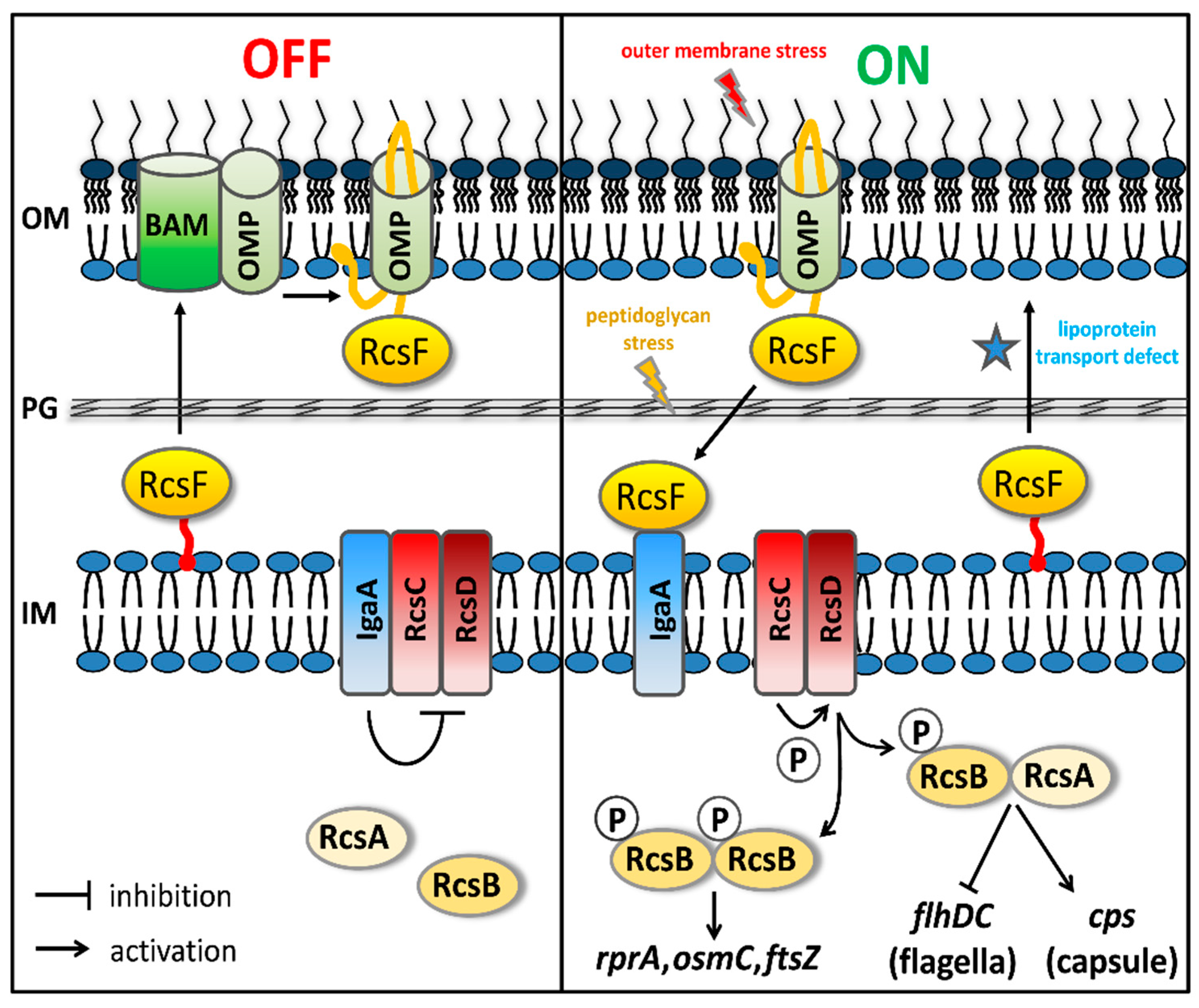
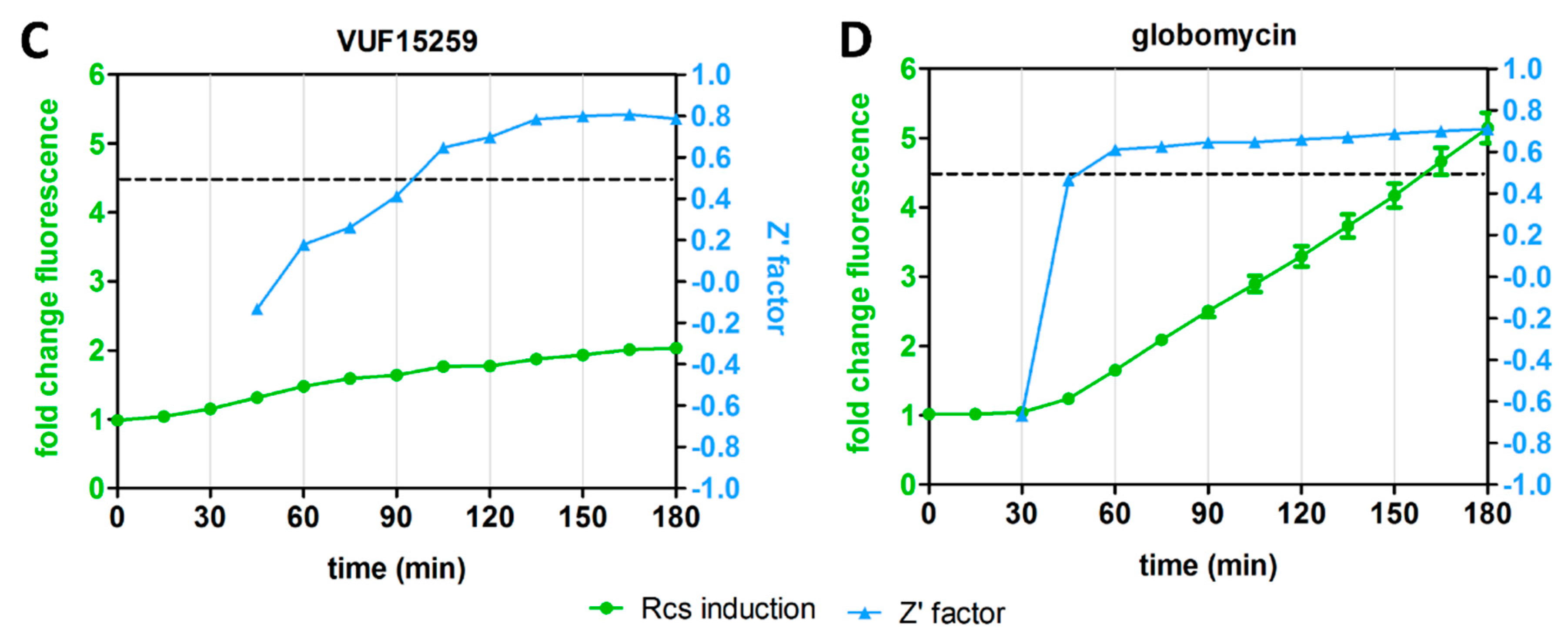
3.3. Inhibition of Biogenesis of PG, Lipoprotein and OMP Predominantly Activates the Rcs Stress Reporter
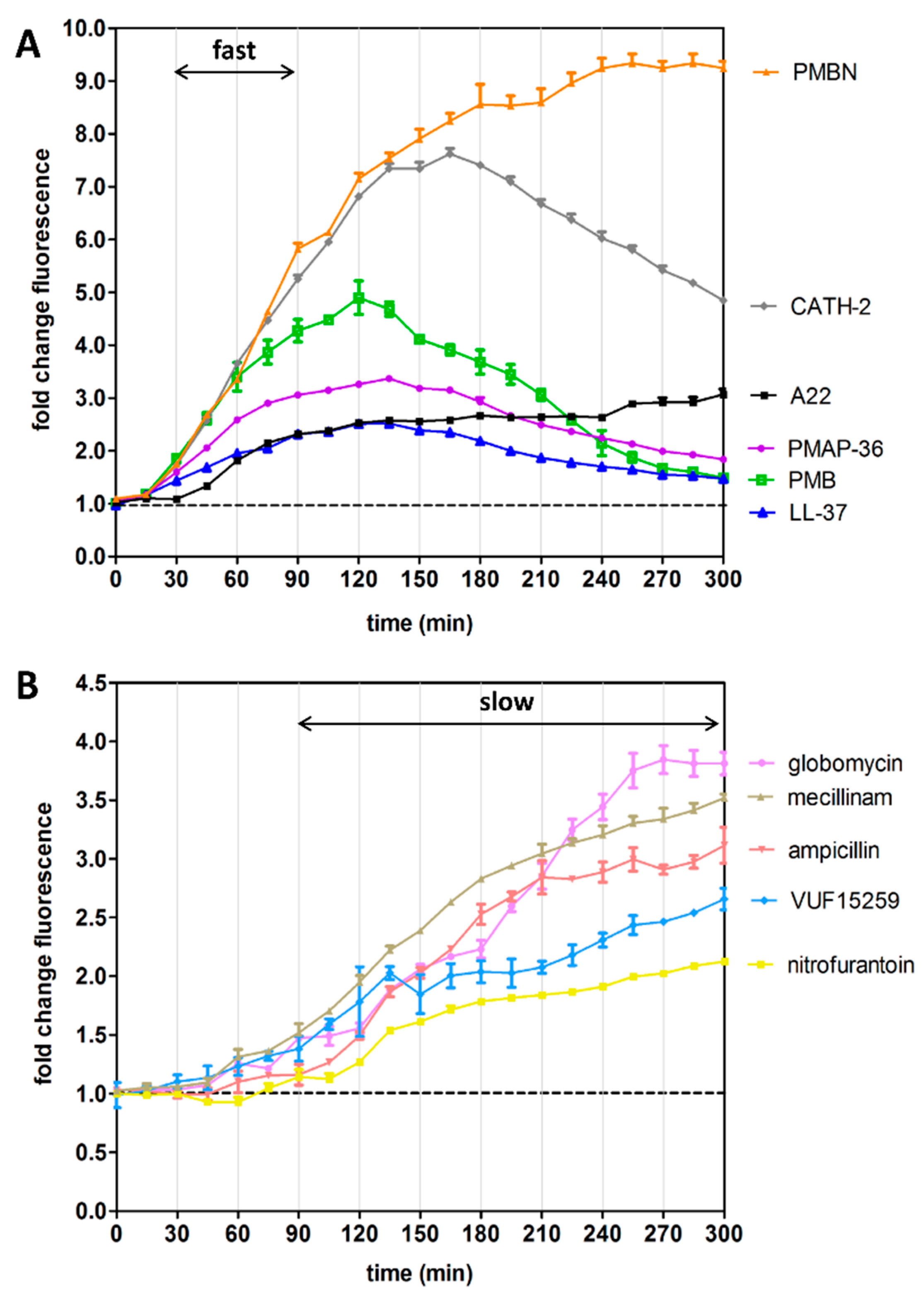
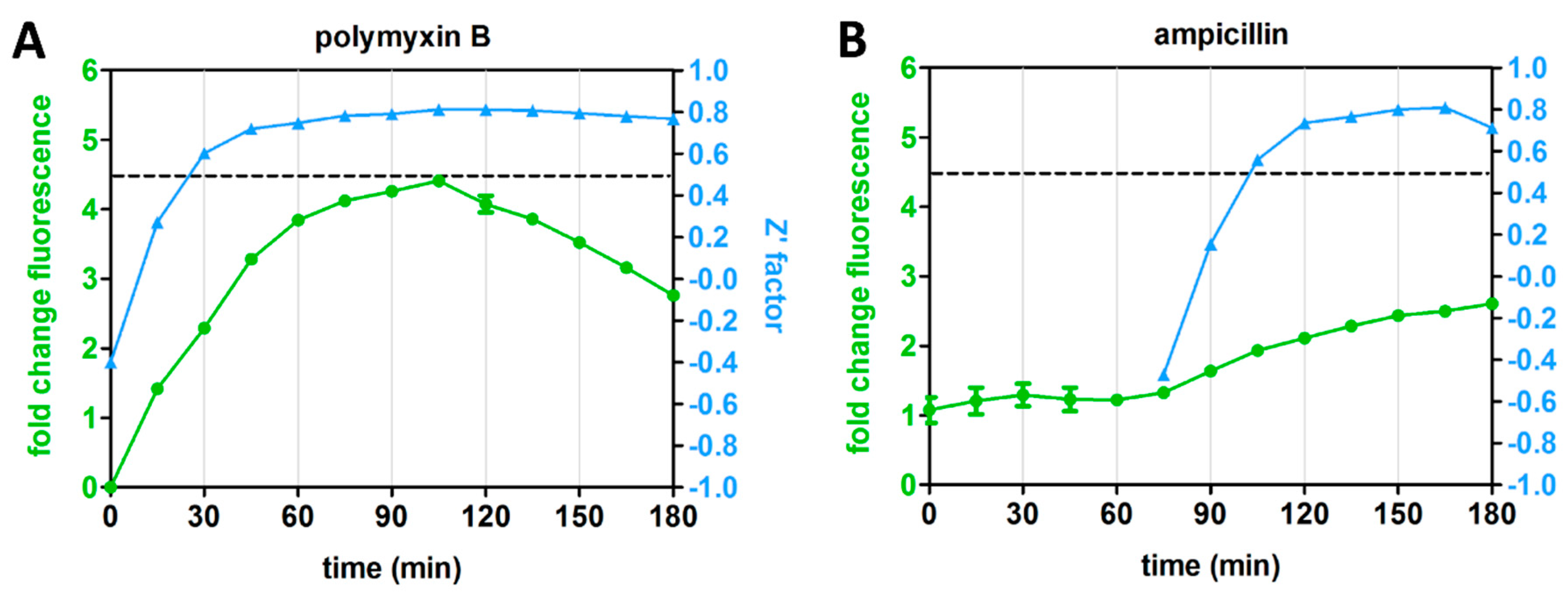
3.4. Kinetics of Rcs Stress Reporter Induction
3.5. Differential Kinetics of Rcs Stress Reporter Induction Can Be Exploited in HTS Format
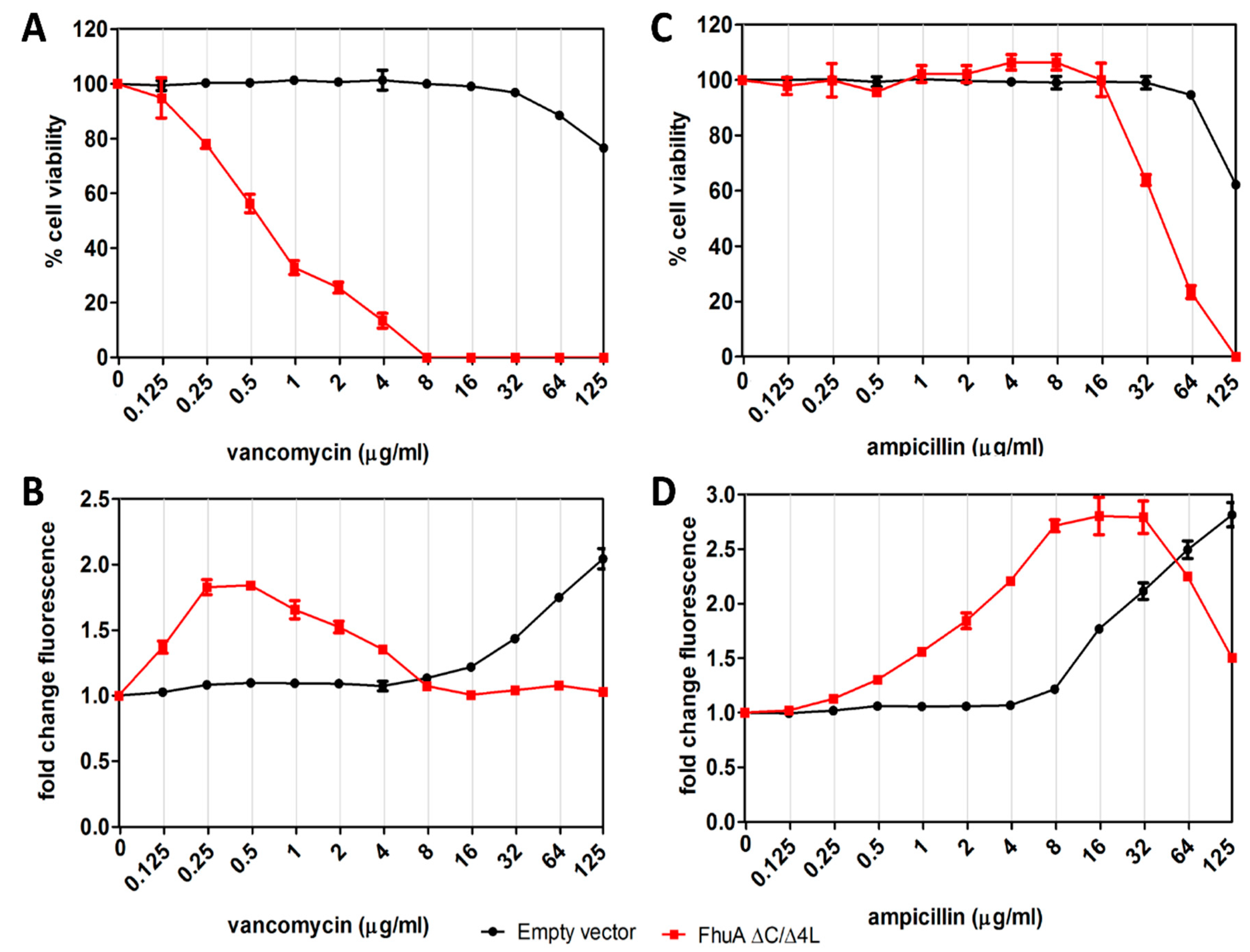
3.6. Expression of FhuA ∆C/∆4L Can Support Identification of Large-Scaffold Inhibitors
4. Discussion and Conclusions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [Green Version]
- Alcalde-Rico, M.; Hernando-Amado, S.; Blanco, P.; Martínez, J.L. Multidrug efflux pumps at the crossroad between antibiotic resistance and bacterial virulence. Front. Microbiol. 2016, 7, 1483. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Bury-Moné, S.; Nomane, Y.; Reymond, N.; Barbet, R.; Jacquet, E.; Imbeaud, S.; Jacq, A.; Bouloc, P. Global analysis of extracytoplasmic stress signaling in Escherichia coli. PLoS Genet. 2009, 5. [Google Scholar] [CrossRef] [Green Version]
- Hews, C.L.; Cho, T.; Rowley, G.; Raivio, T.L. Maintaining integrity under stress: Envelope stress response regulation of pathogenesis in gram-negative bacteria. Front. Cell. Infect. Microbiol. 2019, 9, 313. [Google Scholar] [CrossRef] [Green Version]
- Wall, E.; Majdalani, N.; Gottesman, S. The complex rcs regulatory cascade. Ann. Rev. Microbiol. 2018, 72, 111–139. [Google Scholar] [CrossRef]
- Laloux, G.G.; Collet, J.F. Major tom to ground control: How lipoproteins communicate extra-cytoplasmic stress to the decision center of the cell. J. Bacteriol. 2017, 199. [Google Scholar] [CrossRef] [Green Version]
- Létoquart, J.; Rodriguez-Alonso, R.; Nguyen, V.S.; Louis, G.; Calabrese, A.N.; Radford, S.E.; Cho, S.-H.; Remaut, H.; Collet, J.F. Structural insight into the formation of lipoprotein-β-barrel complexes by the β-barrel assembly machinery. Nat. Chem. Biol. 2020, 1019–1025. [Google Scholar] [CrossRef] [Green Version]
- Steenhuis, M.; Abdallah, A.M.; de Munnik, S.M.; Kuhne, S.; Westerhausen, S.; Wagner, S.; van der Wel, N.N.; Wijtmans, M. Inhibition of autotransporter biogenesis by small molecules. Mol. Microbiol. 2019, 112, 81–98. [Google Scholar] [CrossRef] [Green Version]
- Shaner, N.C.; Lambert, G.G.; Chammas, A.; Ni, Y.; Cranfill, P.J.; Baird, M.A.; Sell, B.R.; Allen, J.R.; Day, R.N.; Israelsson, M.; et al. A bright monomeric green fluorescent protein derived from branchiostoma lanceolatum. Nat. Methods 2013, 10, 407–409. [Google Scholar] [CrossRef]
- Zhang, J.; Chung, T.D.Y.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef]
- Laubacher, M.E.; Ades, S.E. The Rcs phosphorelay is a cell envelope stress response activated by peptidoglycan stress and contributes to intrinsic antibiotic resistance. J. Bacteriol. 2008, 190, 2065–2074. [Google Scholar] [CrossRef] [Green Version]
- Farris, C.; Sanowar, S.; Bader, M.W.; Pfuetzner, R.; Miller, S.I. Antimicrobial peptides activate the Rcs regulon through the outer membrane lipoprotein RcsF. J. Bacteriol. 2010, 192, 4894–4903. [Google Scholar] [CrossRef] [Green Version]
- DiGiuseppe, P.A.; Silhavy, T.J. Signal detection and target gene induction by the CpxRA two-component system. J. Bacteriol. 2003, 185, 2432–2440. [Google Scholar] [CrossRef] [Green Version]
- Konovalova, A.; Mitchell, A.M.; Silhavy, T.J. A lipoprotein/b-barrel complex monitors lipopolysaccharide integrity transducing information across the outer membrane. Elife 2016, 5, 1–17. [Google Scholar] [CrossRef]
- Shiba, Y.; Matsumoto, K.; Hara, H. DjlA negatively regulates the rcs signal transduction system in Escherichia coli. Genes Genet. Syst. 2006, 81, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Delhaye, A.; Laloux, G.; Collet, J.F. The lipoprotein NlpE is a Cpx sensor that serves as a sentinel for protein sorting and folding defects in the Escherichia coli envelope. J. Bacteriol. 2019, 201, e00611–e00618. [Google Scholar] [CrossRef] [Green Version]
- Steenhuis, M.; Koningstein, G.M.; Oswald, J.; Pick, T.; Koch, G.; Whitehead, R.C.; Swanton, E.; High, S.; Luirink, J. Eeyarestatin 24 impairs SecYEG-dependent protein trafficking and inhibits growth of clinically relevant pathogens. Mol. Microbiol. 2020, 115, 1–17. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Tomoyasu, T.; Takaya, A.; Morioka, M.; Yamamoto, T. Effects of disruption of heat shock genes on susceptibility of Escherichia coli to fluoroquinolones. BMC Microbiol. 2003, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Vanbogelen, R.A.; Kelley, P.M.; Neidhardt, F.C. Differential induction of Heat Shock, SOS, and Oxidation Stress Regulons and Accumulation of Nucleotides in Escherichia coli. J. Bacteriol. 1987, 169, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.-H.; Szewczyk, J.; Pesavento, C.; Zietek, M.; Banzhaf, M.; Roszczenko, P.; Asmar, A.; Laloux, G.; Hov, A.-K.; Leverrier, P.; et al. Detecting envelope stress by monitoring β-barrel assembly. Cell 2014, 159, 1652–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, R.; Peterson, M.S.; Bentley, W.E. Stochastic Kinetic Analysis of the Escherichia coli Stress Circuit Using Sigma32 -Targeted Antisense. Biotechnol. Bioeng. 2001, 75. [Google Scholar] [CrossRef] [PubMed]
- Soufi, B.; Krug, K.; Harst, A.; Macek, B. Characterization of the E. coli proteome and its modifications during growth and ethanol stress. Front. Microbiol. 2015, 6, 103. [Google Scholar] [CrossRef] [PubMed]
- Genevaux, P.; Wawrzynow, A.; Zylicz, M.; Georgopoulos, C.; Kelley, W.L. DjlA is a third DnaK co-chaperone of Escherichia coli, and DjlA-mediated induction of colanic acid capsule requires DjlA-DnaK interaction. J. Biol. 2001, 276, 7906–7912. [Google Scholar] [CrossRef] [Green Version]
- Shiba, Y.; Miyagawa, H.; Nagahama, H.; Matsumoto, K.; Kondo, D.; Matsuoka, S.; Matsumoto, K. Exploring the relationship between lipoprotein mislocalization and activation of the Rcs signal transduction system in Escherichia coli. Microbiology 2012, 158, 1238–1248. [Google Scholar] [CrossRef] [Green Version]
- Jong, W.S.P.P.; ten Hagen-Jongman, C.M.; den Blaauwen, T.; Slotboom, D.J.; Tame, J.R.H.H.; Wickström, D.; de Gier, J.-W.; Otto, B.R.; Luirink, J.; Jan Slotboom, D.; et al. Limited tolerance towards folded elements during secretion of the autotransporter Hbp. Mol. Microbiol. 2007, 63, 1524–1536. [Google Scholar] [CrossRef] [Green Version]
- Scheenstra, M.R.; van den Belt, M.; Tjeerdsma-van Bokhoven, J.L.M.; Schneider, V.A.F.; Ordonez, S.R.; van Dijk, A.; Veldhuizen, E.J.A.; Haagsman, H.P. Cathelicidins PMAP-36, LL-37 and CATH-2 are similar peptides with different modes of action. Sci. Rep. 2019, 9, 4780. [Google Scholar] [CrossRef]
- Tsubery, H.; Ofek, I.; Cohen, S.; Fridkin, M. Structure-function studies of polymyxin B nonapeptide: Implications to sensitization of gram-negative bacteria. J. Med. Chem. 2000, 43, 3085–3092. [Google Scholar] [CrossRef]
- McMurry, L.M.; Oethinger, M.; Levy, S.B. Triclosan targets lipid synthesis. Nature 1998, 394, 531–532. [Google Scholar] [CrossRef]
- El Arnaout, T.; Soulimane, T. Targeting lipoprotein biogenesis: Considerations towards antimicrobials. Trends Biochem. Sci. 2019, 44, 701–715. [Google Scholar] [CrossRef]
- Konovalova, A.; Kahne, D.E.; Silhavy, T.J. Outer membrane biogenesis. Ann. Rev. Microbiol. 2017, 71, 539–556. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, N.; Silhavy, T.J. Sensing external stress: Watchdogs of the Escherichia coli cell envelope. Curr. Opin. Microbiol. 2005, 8, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Meyer, K.J.; Iinishi, A.; Favre-Godal, Q.; Green, R.; Manuse, S.; Caboni, M.; Mori, M.; Niles, S.; Ghiglieri, M.; et al. A new antibiotic selectively kills gram-negative pathogens. Nature 2019, 576, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Kotova, V.Y.; Mironov, A.S.; Zavilgelsky, G.B. Role of reactive oxygen species in the bactericidal action of quinolones as inhibitors of DNA gyrase. Mol. Biol. 2014, 48, 870–877. [Google Scholar] [CrossRef]
- Huttner, A.; Verhaegh, E.M.; Harbarth, S.; Muller, A.E.; Theuretzbacher, U.; Mouton, J.W. Nitrofurantoin revisited: A systematic review and meta-analysis of controlled trials. J. Antimicrob. Chemother. 2015, 70, 2456–2464. [Google Scholar] [CrossRef] [Green Version]
- Krishnamoorthy, G.; Wolloscheck, D.; Weeks, J.W.; Croft, C.; Rybenkov, V.V.; Zgurskaya, H.I. Breaking the permeability barrier of Escherichia coli by controlled hyperporination of the outer membrane. Antimicrob. Agents Chemother. 2016, 60, 7372–7381. [Google Scholar] [CrossRef] [Green Version]
- Niedzwiecki, D.J.; Mohammad, M.M.; Movileanu, L. Inspection of the engineered FhuA Δc/Δ4L protein nanopore by polymer exclusion. Biophys. J. 2012, 103, 2115–2124. [Google Scholar] [CrossRef] [Green Version]
- Hart, E.M.; Mitchell, A.M.; Konovalova, A.; Grabowicz, M.; Sheng, J.; Han, X.; Rodriguez-Rivera, F.P.; Schwaid, A.G.; Malinverni, J.C.; Balibar, C.J.; et al. A small-molecule inhibitor of BamA impervious to efflux and the outer membrane permeability barrier. Proc. Natl. Acad. Sci. USA 2019, 116, 21748–21757. [Google Scholar] [CrossRef] [Green Version]
- Lehman, K.M.; Grabowicz, M. Countering gram-negative antibiotic resistance: Recent progress in disrupting the outer membrane with novel therapeutics. Antibiotics 2019, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Zabawa, T.P.; Pucci, M.J.; Parr, T.R.; Lister, T. Treatment of gram-negative bacterial infections by potentiation of antibiotics. Curr. Opin. Microbiol. 2016, 33, 7–12. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stress Reporters a | |||||||
|---|---|---|---|---|---|---|---|
| Antibacterial a | Concentration | Mechanism of Action | Rcs | σE | Cpx | GroES | |
| ampicillin | 80 µM | CELL ENVELOPE | inhibit PG synthesis | + [12] | − | − | − |
| mecillinam | 3 µM | + [12] | − | − | − | ||
| SDS | n.a. | solubilizes lipid bilayer | − [13] | − | − | − | |
| triclosan | n.a. | inhibits fatty acid biogenesis | − | − | − | − | |
| EDTA | 40 µM | affects LPS integrity | − [13] | − | + [14] | − | |
| PMB | 0.6 µM | affect LPS integrity/ membrane destabilization | + [13,15] | − | − | − | |
| PMBN | 26 µM | + [13] | + | + | − | ||
| LL-37 | 10 µM | + [13] | − | + | − | ||
| CATH-2 | 1.2 µM | + | − | − | − | ||
| PMAP-36 | 1.2 µM | + | − | + | − | ||
| VUF15259 [9] | 100 µM | inhibits BAM complex | + | + [9] | − | − | |
| globomycin | 38 µM | inhibits lipoprotein maturation | + [15,16] | − | + [17] | − | |
| nitrofurantoin | 0.6 µM | CYTOSOL | general oxidative damage | + | − | − | + [18] |
| levofloxacin | 2.7 nM | inhibit DNA synthesis | − | − | − | + [19] | |
| nalidixic acid | 4.3 µM | − | − | − | + [20] | ||
| chloramphenicol | n.a. | inhibit protein synthesis | − | − | − | − | |
| tetracycline | n.a. | − | − | − | − | ||
| A22 | 11 µM | disrupts morphology and chromosome segregation | + [15,21] | + | − | + | |
| sulfamethoxazole | 3.9 µM | inhibits folic acid synthesis | − | − | − | − | |
| ethanol | 2.1 nM | GENERAL | generic protein and membrane damage | + | + [22,23] | + | + [22,23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steenhuis, M.; ten Hagen-Jongman, C.M.; van Ulsen, P.; Luirink, J. Stress-Based High-Throughput Screening Assays to Identify Inhibitors of Cell Envelope Biogenesis. Antibiotics 2020, 9, 808. https://doi.org/10.3390/antibiotics9110808
Steenhuis M, ten Hagen-Jongman CM, van Ulsen P, Luirink J. Stress-Based High-Throughput Screening Assays to Identify Inhibitors of Cell Envelope Biogenesis. Antibiotics. 2020; 9(11):808. https://doi.org/10.3390/antibiotics9110808
Chicago/Turabian StyleSteenhuis, Maurice, Corinne M. ten Hagen-Jongman, Peter van Ulsen, and Joen Luirink. 2020. "Stress-Based High-Throughput Screening Assays to Identify Inhibitors of Cell Envelope Biogenesis" Antibiotics 9, no. 11: 808. https://doi.org/10.3390/antibiotics9110808