β-Lytic Protease of Lysobacter capsici VKM B-2533T

Abstract
:1. Introduction
2. Results
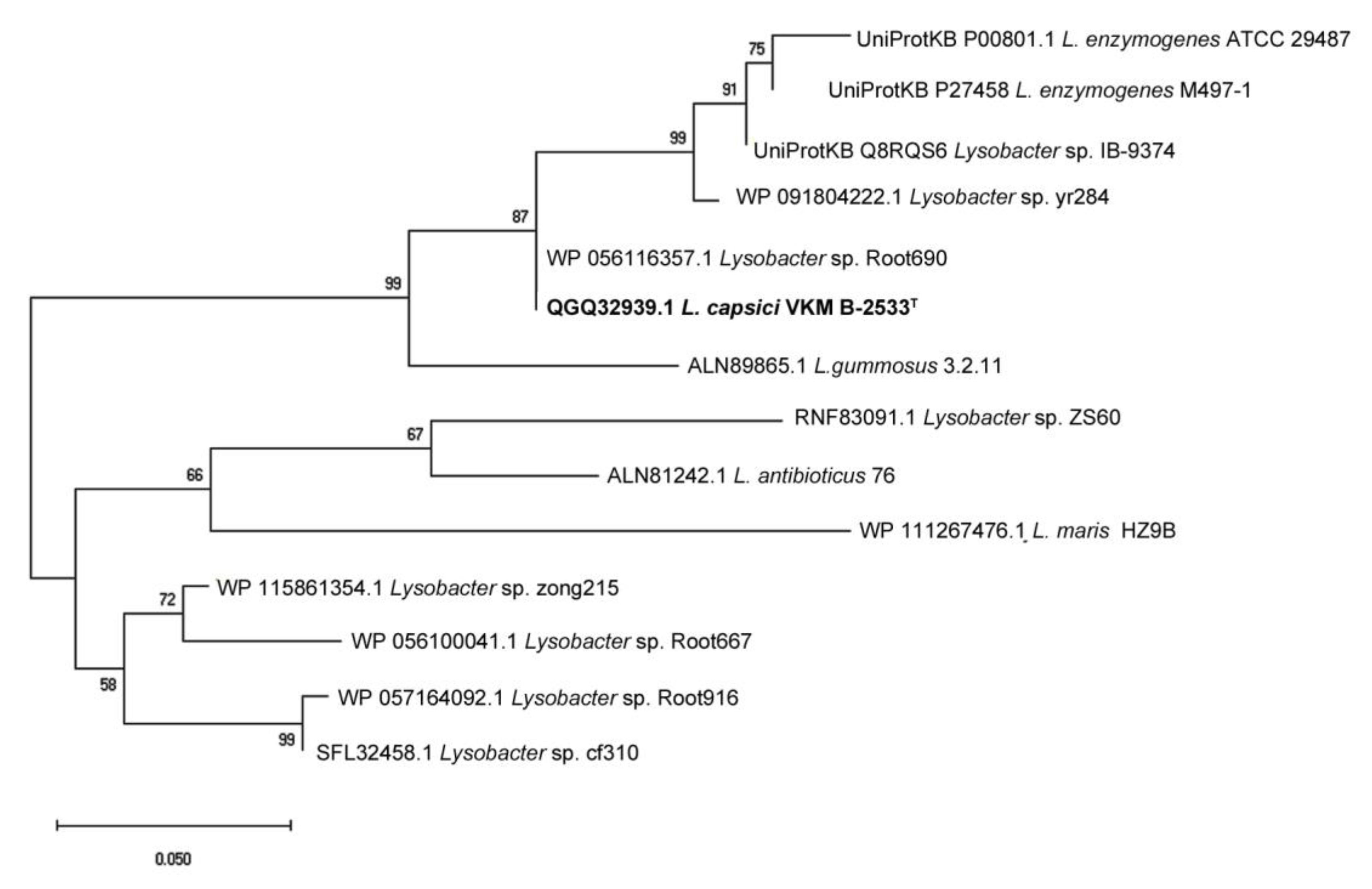
2.1. Phylogenetic Analysis
2.2. Physico-Chemical Properties of BlpLc
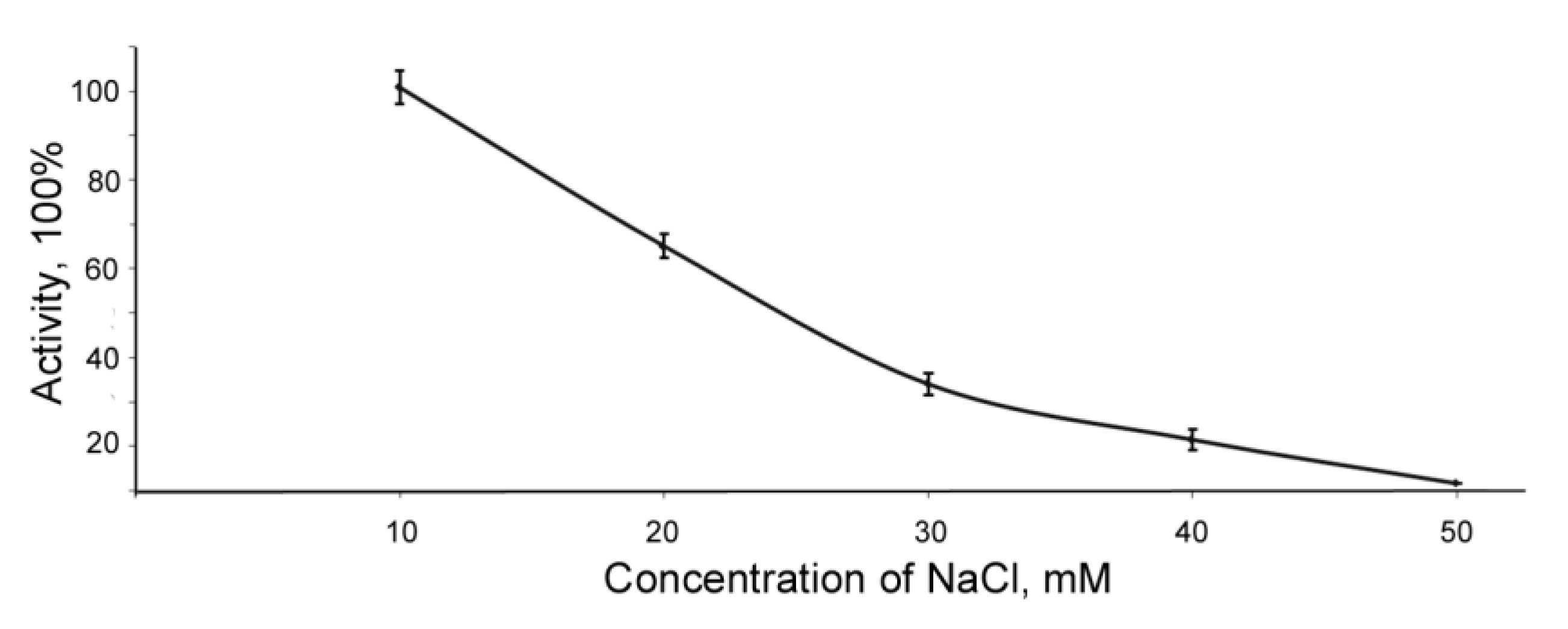
2.3. Effect of Inhibitors on BlpLc Activity
2.4. Determination of BlpLc Specificity of Action and Hydrolyzed Bond Type
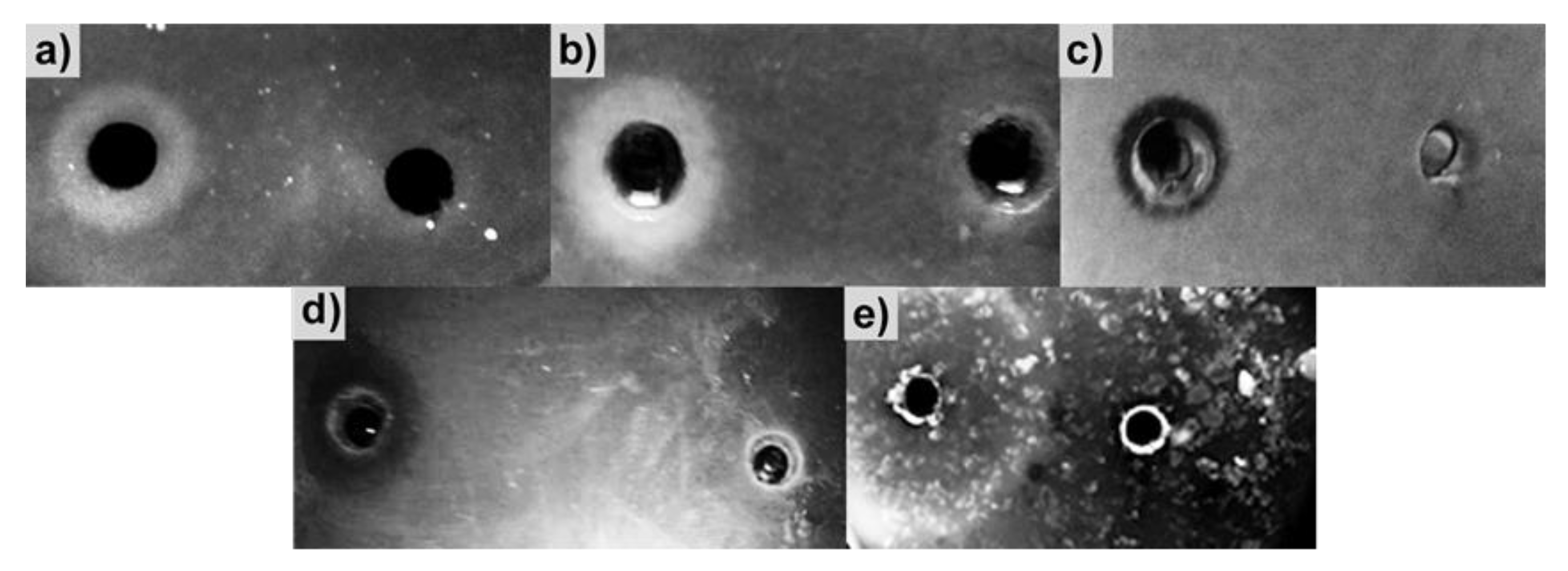
2.5. Antimicrobial Action of BlpLc
3. Discussion
4. Materials and Methods
4.1. Strain and Cultivation Conditions
4.2. Isolation of BlpLc
4.3. Protein Concentration Assay
4.4. Determination of BlpLc Bacteriolytic and Proteolytic Activity
4.5. Electrophoresis of Proteins
4.6. Determination of Conditions for Optimal Manifestation of BlpLc Bacteriolytic Activity
4.7. Effect of Inhibitors on BlpLc Activity
4.8. Determination of the Effect of Metal Substitution in the Enzyme Active Site
4.9. Assay of the Action of Bleacher and Detergents on BlpLc Bacteriolytic Activity
4.10. Determination of BlpLc Substrate Specificity by Spot Test
4.11. Determination of the Type of Hydrolyzed Bond in Protein Substrates by MALDI-TOF
4.12. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fleming, A. On a remarkable bacteriolytic element found in tissues and secretions. Proc. R. Soc. 1922, 93, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Hughey, V.L.; Johnson, E.A. Antimicrobial activity of lysozyme against bacteria involved in food spoilage and food-borne disease. Appl. Environ. Microbiol. 1987, 53, 2165–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callewaert, L.; Michiels, C.W. Lysozymes in the animal kingdom. J. Biosci. 2010, 35, 127–160. [Google Scholar] [CrossRef]
- Aminlari, L.; Hashemi, M.M.; Aminlari, M. Modified lysozymes as novel broad spectrum natural antimicrobial agents in foods. J. Food Sci. 2014, 79, R1077–R1090. [Google Scholar] [CrossRef]
- Vollmer, W.; Joris, B.; Charlier, P.; Foster, S. Bacterial peptidoglycan (murein) hydrolases. FEMS Microbiol. Rev. 2008, 32, 259–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, A.; Wolfe, R.S. Extracellular enzyme from Myxobacter AL-1 that exhibits both beta-1,4-glucanase and chitosanase activites. J. Bacteriol. 1974, 120, 844–853. [Google Scholar] [CrossRef] [Green Version]
- Stepnaya, O.A.; Ledova, L.A.; Kulaev, I.S. Bacteriolytic enzymes. In Uspekhi biologicheskoy khimii (Russia); Poglazov, B.F., Ed.; RAS: Moscow, Russia, 1999; Volume 39, pp. 327–354. [Google Scholar]
- Vasilyeva, N.V.; Tsfasman, I.M.; Suzina, N.E.; Stepnaya, O.A.; Kulaev, I.S. Secretion of bacteriolytic endopeptidase L5 of Lysobacter sp. XL1 into the medium by means of outer membrane vesicles. FEBS J. 2008, 275, 3827–3835. [Google Scholar] [CrossRef]
- Xie, Y.; Wright, S.; Shen, Y.; Du, L. Bioactive natural products from Lysobacter. Nat. Prod. Rep. 2012, 29, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Masschelein, J.; Jenner, M.; Challis, G.L. Antibiotics from Gram-negative bacteria: A comprehensive overview and selected biosynthetic highlights. Nat. Prod. Rep. 2017, 34, 712–783. [Google Scholar] [CrossRef]
- Puopolo, G.; Tomada, S.; Pertot, I. The impact of the omics era on the knowledge and use of Lysobacter species to control phytopathogenic micro-organisms. J. Appl. Microbiol. 2018, 124, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Peterson, E.A.; Gillespie, D.C.; Cook, F.D. A wide-spectrum antibiotic produced by a species of Sorangium. Can. J. Microbiol. 1966, 12, 221–230. [Google Scholar] [CrossRef]
- Whitaker, D.R. Lytic enzymes of Sorangium sp. Isolation and enzymatic properties of the alpha- and beta-lytic proteases. Can. J. Biochem. 1965, 43, 1935–1954. [Google Scholar] [CrossRef]
- Christensen, P.; Cook, F.D. Lysobacter, a new genus of nonfruiting, gliding bacteria with a high base ratio. Int. J. Syst. Evol. Microbiol. 1978, 28, 367–393. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Norioka, S.; Sakiyama, F. Purification, Characterization, and Primary Structure of a Novel Cell Wall Hydrolytic Amidase, CwhA, from Achromobacter lyticus. J. Biochem. 2000, 127, 1033–1039. [Google Scholar] [CrossRef]
- Stepnaya, O.A.; Begunova, E.A.; Tsfasman, I.M.; Kulaev, I.S. Lysoamidase: A bacteriolytic enzyme preparation. Isolation and identification of some physico-chemical properties of extracellular muramidase of the bacteria Xanthomonas sp. Biokhimiia 1996, 61, 648–655. [Google Scholar]
- Stepnaia, O.A.; Begunova, E.A.; Tsfasman, I.M.; Kulaev, I.S. [Bacteriolytic enzyme preparation lysoamidase. Purification and some properties of bacteriolytic peptidase L1]. Biokhimiia 1996, 61, 656–663. [Google Scholar]
- Stepnaya, O.A.; Tsfasman, I.M.; Logvina, I.A.; Ryazanova, L.P.; Muranova, T.A.; Kulaev, I.S. Isolation and characterization of a new extracellular bacteriolytic endopeptidase of Lysobacter sp. XL1. Biochemistry 2005, 70, 1031–1037. [Google Scholar] [CrossRef]
- Begunova, E.A.; Stepnaya, O.A.; Lysanskaya, V.Y.; Kulaev, I.S. Specificity of the action of lysoamidase on Staphylococcus aureus 209P cell walls. Biochemistry 2003, 68, 735–739. [Google Scholar] [CrossRef]
- Vasilyeva, N.V.; Shishkova, N.A.; Marinin, L.I.; Ledova, L.A.; Tsfasman, I.M.; Muranova, T.A.; Stepnaya, O.A.; Kulaev, I.S. Lytic peptidase L5 of Lysobacter sp. XL1 with broad antimicrobial spectrum. J. Mol. Microbiol. Biotechnol. 2014, 24, 59–66. [Google Scholar] [CrossRef]
- Fuhrmann, C.N.; Kelch, B.A.; Ota, N.; Agard, D.A. The 0.83 Å resolution crystal structure of alpha-lytic protease reveals the detailed structure of the active site and identifies a source of conformational strain. J. Mol. Biol. 2004, 338, 999–1013. [Google Scholar] [CrossRef]
- Tishchenko, S.; Gabdulkhakov, A.; Melnik, B.; Kudryakova, I.; Latypov, O.; Vasilyeva, N.; Leontievsky, A. Structural studies of component of lysoamidase bacteriolytic complex from Lysobacter sp. XL1. Protein J. 2016, 35, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Kudryakova, I.V.; Gabdulkhakov, A.G.; Tishchenko, S.V.; Lysanskaya, V.Y.; Suzina, N.E.; Tsfasman, I.M.; Afoshin, A.S.; Vasilyeva, N.V. Structural and functional properties of antimicrobial protein L5 of Lysobacter sp. XL1. Appl. Microbiol. Biotechnol. 2018, 102, 10043–10053. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Norioka, S.; Sakiyama, F. Bacteriolytic activity and specificity of Achromobacter beta-lytic protease. J. Biochem. 1998, 124, 332–339. [Google Scholar] [CrossRef]
- Ahmed, K.; Chohnan, S.; Ohashi, H.; Hirata, T.; Masaki, T.; Sakiyama, F. Purification, bacteriolytic activity, and specificity of beta-lytic protease from Lysobacter sp. IB-9374. J. Biosci. Bioeng. 2003, 95, 27–34. [Google Scholar] [CrossRef]
- Afoshin, A.S.; Kudryakova, I.V.; Borovikova, A.O.; Suzina, N.E.; Toropygin, I.Y.; Shishkova, N.A.; Vasilyeva, N.V. Lytic potential of Lysobacter capsici VKM B-2533T: Bacteriolytic enzymes and outer membrane vesicles. Sci. Rep. 2020, 10, 9944. [Google Scholar] [CrossRef] [PubMed]
- Takami, H.; Toyoda, A.; Uchiyama, I.; Itoh, T.; Takaki, Y.; Arai, W.; Nishi, S.H.; Kawai, M.; Shin-Ya, K.; Ikeda, H. Complete genome sequence and expression profile of the commercial lytic enzyme producer Lysobacter enzymogenes M497-1. DNA Res. 2017, 24, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Blobel, G.; Dobberstein, B. Transfer of proteins across membranes. I. Presence of proteolytically processed and unprocessed nascent immunoglobulin light chains on membrane-bound ribosomes of murine myeloma. J. Cell Biol. 1975, 67, 835–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujishige, A.; Smith, K.R.; Silen, J.L.; Agard, D.A. Correct folding of alpha-lytic protease is required for its extracellular secretion from Escherichia coli. J. Cell Biol. 1992, 118, 33–42. [Google Scholar] [CrossRef]
- Oza, N.B. Beta-lytic protease, a neutral sorangiopeptidase. Int. J. Pept. Protein Res. 1973, 5, 365–369. [Google Scholar] [CrossRef]
- Antonov, V.K. Chemistry of Proteolysis, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1993; p. 495. ISBN 978-3-662-00981-9. [Google Scholar]
- Szeci, P.B.; Harboe, M. Chymosin. In Handbook of Proteolytic Enzymes, 3rd ed.; Rawlings, N.D., Salvesen, G.S., Eds.; Academic Press: London, UK, 2013; Volume 1, pp. 37–42. ISBN 978-0-12-407744-7. [Google Scholar]
- Whitaker, D.R.; Roy, C.; Tsai, C.S.; Jurásek, L. Lytic enzymes of Sorangium sp. A comparison of the proteolytic properties of the alpha- and beta-lytic proteases. Can. J. Biochem. 1965, 43, 1961–1970. [Google Scholar] [CrossRef]
- Brito, N.; Falcón, M.A.; Carnicero, A.; Gutiérrez-Navarro, A.M.; Mansito, T.B. Purification and peptidase activity of a bacteriolytic extracellular enzyme from Pseudomonas aeruginosa. Res. Microbiol. 1989, 140, 125–137. [Google Scholar] [CrossRef]
- Kessler, E.; Safrin, M.; Abrams, W.R.; Rosenbloom, J.; Ohman, D.E. Inhibitors and specificity of Pseudomonas aeruginosa LasA. J. Biol. Chem. 1997, 272, 9884–9889. [Google Scholar] [CrossRef] [Green Version]
- Barequet, I.S.; Ben Simon, G.J.; Safrin, M.; Ohman, D.E.; Kessler, E. Pseudomonas aeruginosa LasA protease in treatment of experimental staphylococcal keratitis. Antimicrob. Agents. Chemother. 2004, 48, 1681–1687. [Google Scholar] [CrossRef] [Green Version]
- Schleifer, K.H.; Kandler, O. Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol. Rev. 1972, 36, 407–477. [Google Scholar] [CrossRef] [Green Version]
- Lillo, A.; Quirós, L.M.; Fierro, J.F. Relationship between antibacterial activity and cell surface binding of lactoferrin in species of genus Micrococcus. FEMS Microbiol. Lett. 1997, 150, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Stepnaia, O.A.; Begunova, E.A.; Tsfasman, I.M.; Tul’skaia, E.M.; Streshinskaia, G.M.; Naumova, I.B.; Kulaev, I.S. Mechanism of action of the extracellular bacteriolytic enzymes of Lysobacter sp. on Gram-positive bacteria: Role of the cell wall anionic polymers of the target bacteria. Mikrobiologiia 2004, 73, 479–485. [Google Scholar] [CrossRef]
- Ogura, J.; Toyoda, A.; Kurosawa, T.; Chong, A.L.; Chohnan, S.; Masaki, T. Purification, characterization, and gene analysis of cellulase (Cel8A) from Lysobacter sp. IB-9374. Biosci. Biotechnol. Biochem. 2006, 70, 2420–2428. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hull, M.E. Studies on milk proteins. II. Colorimetric determination of the partial hydrolysis of the proteins in milk. J. Diary Sci. 1974, 30, 881–884. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Käll, L.; Krogh, A.; Sonnhammer, E.L. Advantages of combined transmembrane topology and signal peptide prediction—The Phobius web server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [Green Version]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protease | Region | Signal Peptide Length | ||||
|---|---|---|---|---|---|---|
| N | H | C | ||||
| Length, aa | Net Charge | Length, aa | Hydropathy Index | Length, aa | ||
| L. capsici VKM B-2533T GenBank no. QGQ32939.1 | 9 MKAIS GARI | +2 | 12 TLLAGCVLIAIC | 2.675 | 5 GGAAA | 26 |
| Lysobacter sp. IB-9374 UniProtKB Q8RQS6 | 6 MKKISK | +3 | 12 AGLGLALMCALA | 2.167 | 8 TIGGNASA | 26 |
| L. enzymogenes M497-1 UniProtKB-P27458 | 6 MKKISK | +3 | 12 AGLGLALVCALA | 2.358 | 12 TIGGNA ARRATA | 30 |
| Metal salt | Inhibition, % |
|---|---|
| CaCl2 | 25.0 ± 2.5 |
| ZnCl2 | 93.0 ± 1.4 |
| CuSO4 | 99.10 ± 0.07 |
| NiSO4 | 54.5 ± 5.4 |
| FeSO4 | 69.4 ± 3.0 |
| MgCl2 | 28.7 ± 3.2 |
| MnCl2 | 39.2 ± 2.7 |
| Agent | Concentration, % | Residual Activity, % |
|---|---|---|
| SDS | 1 | 54.9 ± 3.7 |
| 5 | 0 | |
| Triton X-100 | 1 | 109.0 ± 2.3 |
| 5 | 10.9 ± 1.9 | |
| H2O2 | 1 | 67.0 ± 1.3 |
| 5 | 26.0 ± 2.1 |
| Inhibitor | Concentration, mM | Inhibition, % |
|---|---|---|
| 1,10-Phenanthroline | 1.0 | 41.0 ± 3.6 |
| 2.5 | 84.0 ± 2.4 | |
| 5.0 | 100.0 | |
| p-CMB | 1.0 | 79.0 ± 1.7 |
| 5.0 | 100.0 | |
| PMSF | 5.0 | 75 ± 0.28 |
| 10.0 | 100.0 | |
| EDTA * | 10.0 | 0 |
| 50.0 | 100.0 |
| Parameter | Lysobacter sp. IB-9374 * [25] | L. enzymogenes M497-1 ** [24,30] | L. capsici VKM B-2533T *** (This Work) |
|---|---|---|---|
| pH optimum | 8.0 | 6.5 (substrate: FAGLA) 10.0 (substrates: S. aureus, M. luteus) | 9.0 |
| Molarity optimum | ND | 35 mM (substrate: S. aureus) 20 mM (substrate: M. luteus) | 5 mM |
| Temperature optimum | 65 °C | ND | 50 °C |
| Thermal stability of enzyme/enzyme half-inactivation temperature | Up to 50 °C | ND | 57 °C |
| pH stability | 6–12 | ND | 4–7; 10–11 |
| Inhibitors | Chelating and reducing agents. Insensitivity to inhibitors of serine, cysteine, aspartate proteases. Substrate: neuromidin (L. enzymogenes) and M. luteus (Lysobacter sp. IB-9374) | Chelating agents, inhibitors of serine and cysteine proteases | |
| Metals | Ca 2+, Mg2+, Ba2+, Fe2+, do not affect activity Mn2+, Ni2+, Co2+, 30% inactivation Zn2+, Cd2+, Hg2+, Cu2+, 95% inactivation | ND | Ca2+, Mg2+, up to 30% inactivation; Mn2+, Ni2+, up to 55% inactivation; Fe2+, 69% inactivation; Zn2+, Cu2+, >93% inactivation |
| Protease (Producer) | Spectrum of Bacteriolytic Action | Hydrolysis of the Bond in Peptidoglycan | Hydrolysis of the Bond in Proteins and Peptides | Ref. |
|---|---|---|---|---|
| Blp of L. enzymogenes M497-1 and ATCC29487; Lysobacter sp. IB-9374 | Freeze-dried cells of M. luteus IF0 3333; S. aureus IAM 12544; S. caseolyticus ATCC 13548; Microbacterium arborescens JCM 5884; B. subtilis JCM 1465; Arthrobacter globiformis JCM 1332; Enterococcus faecalis JCM 5803; Corynebacterium aquaticum JCM 136; Lactobacillus sake JCMl157; L. plantarum JCM 1149; L. casei JCM 1134; Lactococcus lactis JCM 5805; Pediococcus acidilactici JCM 2032; E. coli JCM 1649; Xanthomonas sp. IF0 3085; Beijiernckia indica IF0 3744; Enterobacter aerogenes JCM1235 | Gly-Gly, D-Ala-Gly S. aureus; D-Ala-L-Ala of M. luteus peptide bridge | Gly-Gly; Gly-Asn; Gly-Asp; Gly-His; Gly-Trp; Gly-Leu; Gly-Phe; Asn-Leu; Val-Cys | [24,25,30,33] |
| BlpLc | Living cells of S. aureus 55 (MRSA; S. aureus 209P; M. luteus B1819; M. roseus B1236 | Not determined | Gly-Gly; Gly-Asn; Gly-His; Gly-Phe; Gly-Ala *; Gly-Lys *; Gly-Ser *;Gly-Thr *; Lys-Glu *; Glu-Leu *; Ser-Lys *; Phe-Thr *; Ala-Val *; Ala-Ala *; Ala-His *; Ala-Ser *; Val-Ser * | This work |
| Staphylolysin LasA Pseudomonas aeruginosa (Paks I, FRD2, FRD2128) | Living cells of S. aureus (MRSA); Freeze-dried cells of S. saprophyticus CECT 235; S. epidermidis CECT 232; S. warneri CECT 236; Streptomyces griseus CECT 3112 | Gly-Gly S. aureus | Gly-Gly; Gly-Ala; Gly-Phe | [34,35,36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afoshin, A.S.; Konstantinov, M.A.; Toropygin, I.Y.; Kudryakova, I.V.; Vasilyeva, N.V. β-Lytic Protease of Lysobacter capsici VKM B-2533T. Antibiotics 2020, 9, 744. https://doi.org/10.3390/antibiotics9110744
Afoshin AS, Konstantinov MA, Toropygin IY, Kudryakova IV, Vasilyeva NV. β-Lytic Protease of Lysobacter capsici VKM B-2533T. Antibiotics. 2020; 9(11):744. https://doi.org/10.3390/antibiotics9110744
Chicago/Turabian StyleAfoshin, Alexey S., Mihail A. Konstantinov, Ilya Yu. Toropygin, Irina V. Kudryakova, and Natalia V. Vasilyeva. 2020. "β-Lytic Protease of Lysobacter capsici VKM B-2533T" Antibiotics 9, no. 11: 744. https://doi.org/10.3390/antibiotics9110744