Microbiome Analysis of Biofilms of Silver Nanoparticle-Dispersed Silane-Based Coated Carbon Steel Using a Next-Generation Sequencing Technique

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. Sampling Seawater
2.3. Biofilm Formation Test
2.4. DNA Extraction from Seawater and Biofilms
2.5. 16S rRNA Gene-Based Bacterial Community Analysis
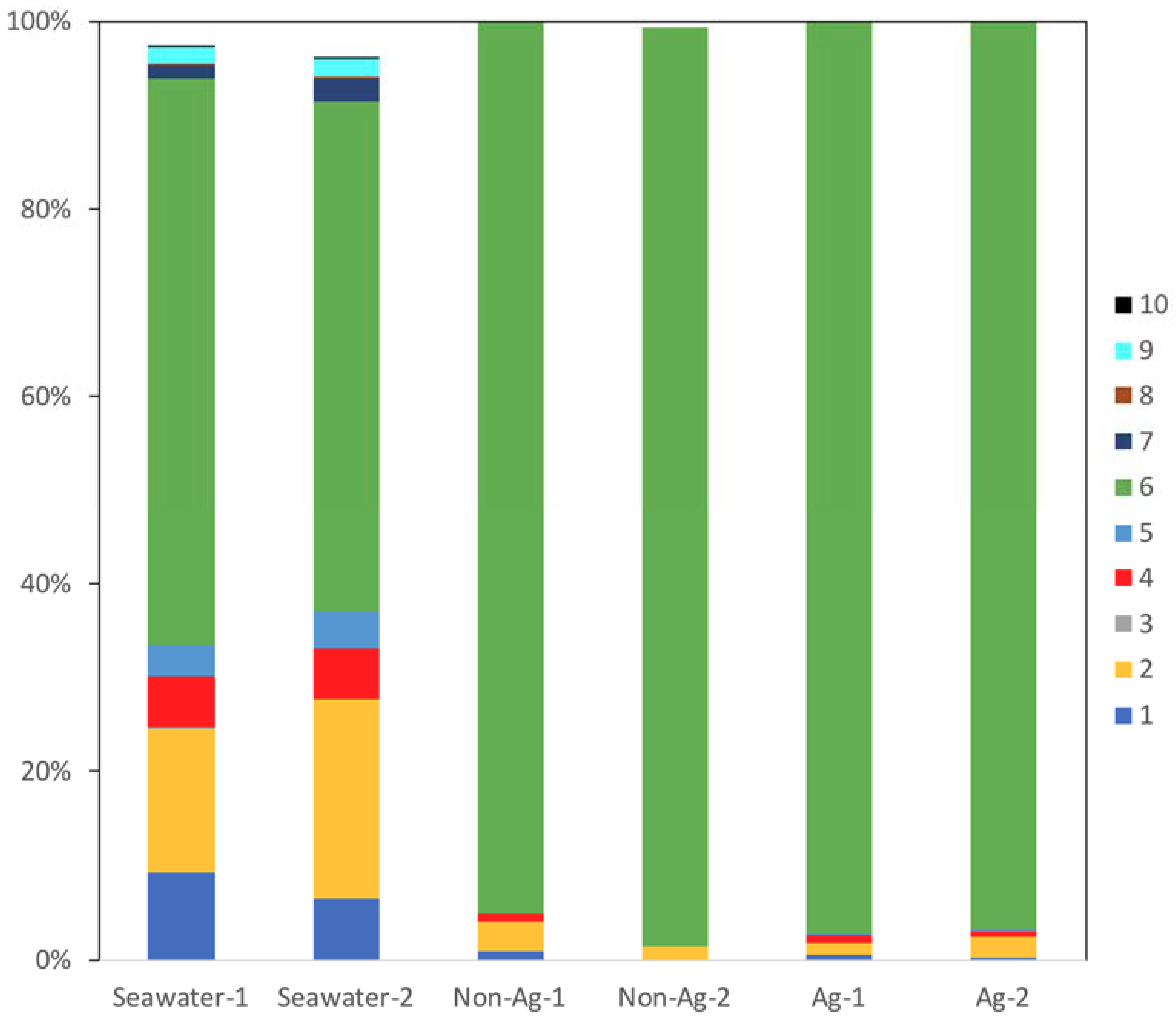
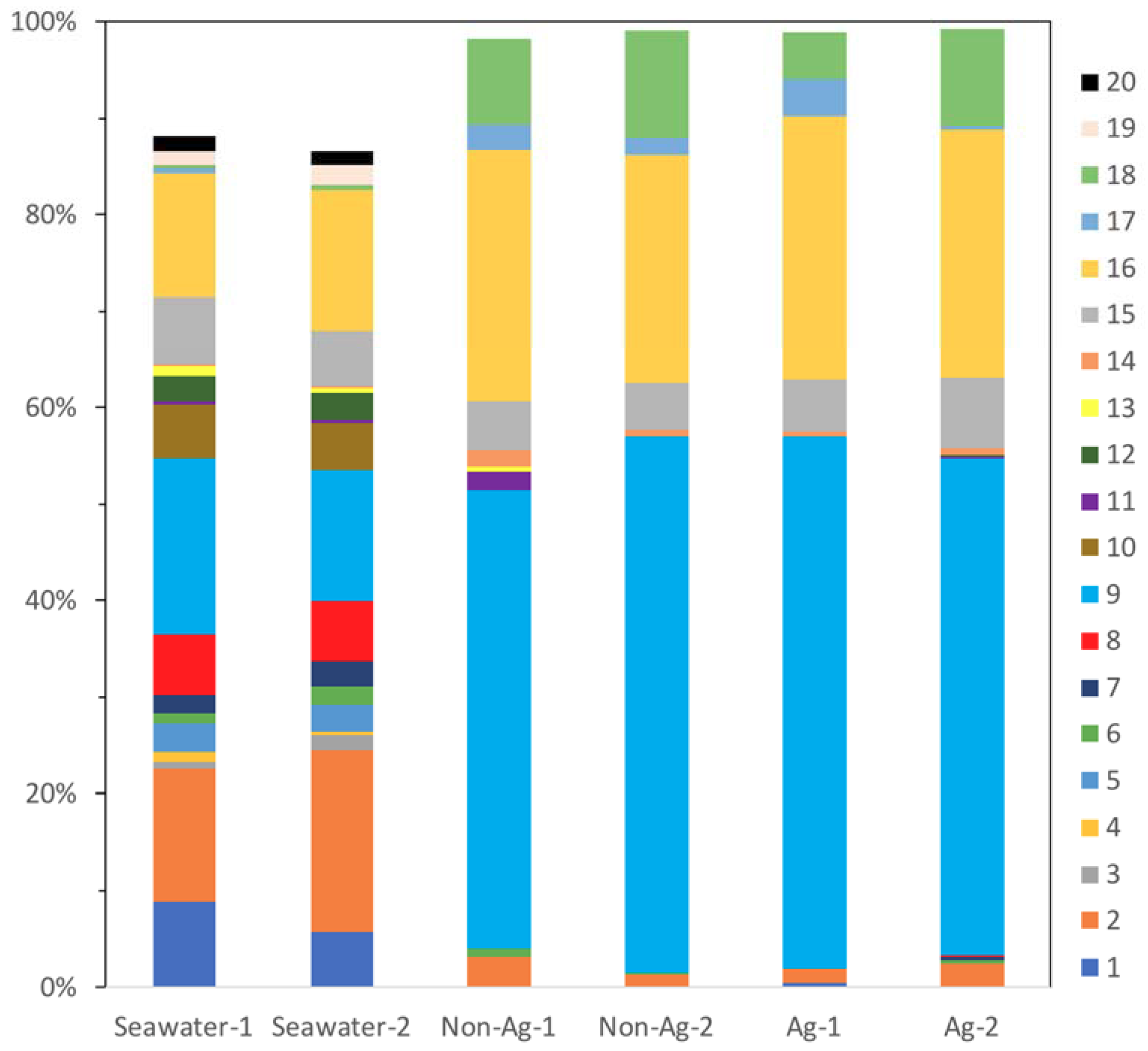
3. Results and Discussions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Flemming, H.C. Biofouling in water systems—Cases, causes and countermeasures. Appl. Microbiol. Biotechnol. 2002, 59, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Bixler, G.D.; Bhushan, B. Biofouling: Lessons from nature. Philos. Trans. A Math. Phys. Eng. Sci. 2012, 370, 2381–2417. [Google Scholar] [CrossRef] [PubMed]
- Kougo, T.; Kuroda, D.; Wada, N.; Ikegai, H.; Kanematsu, H. Biofouling of various metal oxides in marine environment. J. Phys. Conf. Ser. 2012, 352, 012048. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C. Why Microorganisms Live in Biofilms and the Problem of Biofouling. In Marine and Industrial Biofouling; Flemming, H.C., Murthy, S.P., Venkatesan, R., Coolsey, K.E., Eds.; Springer: Berlin, Germany, 2009; pp. 3–12. [Google Scholar]
- Delaunois, F.; Tosar, F.; Vitry, V. Corrosion behavior and biocorrosion of galvanized steel water distribution systems. Bioelectrochemistry 2014, 97, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Beech, I.B.; Sztyler, M.; Gaylarde, C.C.; Smith, W.L.; Sunner, J. Biofilms and biocorrosion. In Understanding Biocorrosion; Liengen, T., Féron, D., Basséguy, R., Beech, I.B., Eds.; Woodhead Publishing: Oxford, UK, 2015; pp. 33–56. [Google Scholar]
- Ogawa, A.; Kanematsu, H.; Sano, K.; Sakai, Y.; Ishida, K.; Beech, I.; Suzuki, O.; Tanaka, T. Effect of Silver or Copper Nanoparticles-Dispersed Silane Coatings on Biofilm Formation in Cooling Water Systems. Materials 2016, 9, 632. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Drive5. Available online: http://www.drive5.com/ (accessed on 12 August 2018).
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FASTX-Toolkit, FASTQ/A Short-Reads Pre-Processing Tools. Available online: http://hannonlab.cshl.edu/fastx_toolkit/ (accessed on 12 August 2018).
- Joshi, N.A.; Fass, J.N. Sickle: A Sliding-Window, Adaptive, Quality-Based Trimming Tool for FastQ Files (Version 1.33) [Software]. 2011. Available online: https://github.com/najoshi/sickle (accessed on 12 August 2018).
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- FLASh Fast Length Ajustment of Short Reads. Available online: https://ccb.jhu.edu/software/FLASH/ (accessed on 12 August 2018).
- The Greengenes Database. Available online: http://greengenes.secondgenome.com (accessed on 12 August 2018).
- Barns, Sue and Siegfried Burggraf: Crenarchaeota. Version 01 January 1997. Available online: http://tolweb.org/Crenarchaeota/9/1997.01.01 (accessed on 15 September 2018).
- Tree of Life Web Project. Euryarchaeota. Version 01 January 1995 (Temporary). 1995. Available online: http://tolweb.org/Euryarchaeota/10/1995.01.01 (accessed on 15 September 2018).
- Huttunen-Saarivirta, E.; Rajala, P.; Marja-Aho, M.; Maukonen, J.; Sohlberg, E.; Carpen, L. Ennoblement, corrosion, and biofouling in brackish seawater: Comparison between six stainless steel grades. Bioelectrochemistry 2018, 120, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Kim, D.; Lee, T. Microbial diversity in biofilms on water distribution pipes of different materials. Water Sci. Technol. 2010, 61, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Wemheuer, B. Diversity and Ecology of the Roseobacter Clade and Other Marine Microbes as Revealed by Metagenomic and Metatranscriptomic Approaches. Ph.D. Thesis, Georg-August University School of Science, Gottingen, Germany, 2015. [Google Scholar]
- Elifantz, H.; Horn, G.; Ayon, M.; Cohen, Y.; Minz, D. Rhodobacteraceae are the key members of the microbial community of the initial biofilm formed in Eastern Mediterranean coastal seawater. FEMS Microbiol. Ecol. 2013, 85, 348–357. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C.C.R. Marine Biofilms: A Successful Microbial Strategy with Economic Implications. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef]
- Woo, P.C.; Teng, J.L.; Leung, K.W.; Lau, S.K.; Woo, G.K.; Wong, A.C.; Wong, M.K.; Yuen, K.Y. Anaerospora hongkongensis gen. nov. sp. nov., a novel genus and species with ribosomal DNA operon heterogeneity isolated from an intravenous drug abuser with pseudobacteremia. Microbiol. Immunol. 2005, 49, 311–339. [Google Scholar] [CrossRef]
- BacDive Strain Identifier BacDive ID: 23337. Available online: https://bacdive.dsmz.de/strain/23337 (accessed on 12 August 2018).
- Okshevsky, M.; Regina, V.R.; Meyer, R.L. Extracellular DNA as a target for biofilm control. Curr. Opin. Biotechnol. 2015, 33, 73–80. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Seawater-1 | Seawater-2 | Non-Ag-1 | Non-Ag-2 | Ag-1 | Ag-2 | Phyla |
|---|---|---|---|---|---|---|
| 0.58 | 0.54 | 0.12 | 0.00 | 0 | 0 | Crenarchaeota |
| 1.20 | 1.90 | 0 | 0 | 0 | 0 | Euryarchaeota |
| 0.02 | 0.03 | 0 | 0.11 | 0 | 0 | Parvarchaeota |
| Seawater-1 | Seawater-2 | Non-Ag-1 | Non-Ag-2 | Ag-1 | Ag-2 | Families | Genera |
|---|---|---|---|---|---|---|---|
| 5.29 | 3.79 | 4.77 | 4.51 | 3.86 | 3.30 | Rhodobacteraceae | Other |
| 5.50 | 4.11 | 3.32 | 2.53 | 3.67 | 1.66 | - | |
| 0.29 | 0 | 0 | 0 | 0 | 0 | Amaricoccus | |
| 6.15 | 4.33 | 34.86 | 44.79 | 47.15 | 40.30 | Anaerospora | |
| 0.06 | 0.04 | 0 | 0.01 | 0.01 | 0 | Loktanella | |
| 0 | 0.02 | 0 | 0 | 0 | 0 | Octadecabacter | |
| 0.01 | 0.01 | 0.02 | 0.42 | 0.01 | 0.03 | Paracoccus | |
| 0.89 | 1.18 | 4.42 | 3.31 | 0.46 | 6.05 | Pseudoruegeria | |
| 0.03 | 0.02 | 0.01 | 0 | 0 | 0 | Rhodobacter | |
| 0 | 0 | 0.01 | 0.01 | 0.01 | 0.01 | Rhodovulum | |
| 0.01 | 0 | 0 | 0.01 | 0.01 | 0.01 | Roseivivax | |
| 0.86 | 0.59 | 0 | 0 | 0 | 0 | Rhodospirillaceae | - |
| 0.04 | 0 | 0 | 0 | 0 | 0 | Magnetospirillum |
| Seawater-1 | Seawater-2 | Non-Ag-1 | Non-Ag-2 | Ag-1 | Ag-2 | Families | Genera |
|---|---|---|---|---|---|---|---|
| 0 | 0 | 1.80 | 0.65 | 0.81 | 1.09 | Oceanospirillaceae | Other |
| 1.72 | 1.70 | 1.89 | 2.58 | 3.72 | 3.92 | - | |
| 0 | 0 | 0 | 0.26 | 0 | 0.15 | Amphritea | |
| 0.28 | 0.32 | 21.76 | 19.56 | 18.97 | 19.60 | Marinomonas | |
| 0 | 0 | 0 | 0 | 0 | 0.27 | Neptunomonas | |
| 0 | 0.01 | 0 | 0 | 0 | 0 | Oceanospirillum | |
| 0 | 0 | 0.64 | 0.30 | 2.40 | 0.06 | Oleispira | |
| 0 | 0 | 0 | 0.31 | 0.56 | 0.41 | - | |
| 0.01 | 0.02 | 0 | 0 | 0 | 0 | SUP05 | - |
| Seawater-1 | Seawater-2 | Non-Ag-1 | Non-Ag-2 | Ag-1 | Ag-2 | Families | Genera |
|---|---|---|---|---|---|---|---|
| 0 | 0 | 0 | 0.01 | 0 | 0.01 | Pseudoalteromonadaceae | - |
| 0.03 | 0.04 | 1.21 | 1.68 | 0.24 | 0.90 | Pseudoalteromonas | |
| 0.41 | 0.43 | 7.30 | 9.47 | 3.27 | 9.17 | Vibrionaceae | Other |
| 0 | 0 | 0.01 | 0.02 | 0 | 0.01 | - | |
| 0.07 | 0.01 | 0.05 | 0 | 1.45 | 0 | Photobacterium | |
| 0.01 | 0.02 | 0.21 | 0.03 | 0.01 | 0.08 | Vibrio |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogawa, A.; Takakura, K.; Sano, K.; Kanematsu, H.; Yamano, T.; Saishin, T.; Terada, S. Microbiome Analysis of Biofilms of Silver Nanoparticle-Dispersed Silane-Based Coated Carbon Steel Using a Next-Generation Sequencing Technique. Antibiotics 2018, 7, 91. https://doi.org/10.3390/antibiotics7040091
Ogawa A, Takakura K, Sano K, Kanematsu H, Yamano T, Saishin T, Terada S. Microbiome Analysis of Biofilms of Silver Nanoparticle-Dispersed Silane-Based Coated Carbon Steel Using a Next-Generation Sequencing Technique. Antibiotics. 2018; 7(4):91. https://doi.org/10.3390/antibiotics7040091
Chicago/Turabian StyleOgawa, Akiko, Keito Takakura, Katsuhiko Sano, Hideyuki Kanematsu, Takehiko Yamano, Toshikazu Saishin, and Satoshi Terada. 2018. "Microbiome Analysis of Biofilms of Silver Nanoparticle-Dispersed Silane-Based Coated Carbon Steel Using a Next-Generation Sequencing Technique" Antibiotics 7, no. 4: 91. https://doi.org/10.3390/antibiotics7040091