
Variation in Pen-Level Prevalence of BRD Bacterial Pathogens and Antimicrobial Resistance Following Feedlot Arrival in Beef Calves
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Study Population
2.2. Differences in Bacterial Recovery between Years and Metaphylaxis Options
2.3. Differences in Bacterial Recovery over Time within Year and Metaphylaxis Options
2.4. Bacterial Co-Isolation Patterns in Years 2020 and 2021 at 1DOF and 13DOF
2.5. Differences in Antimicrobial Susceptibility of Bacteria between Years at 1DOF
2.6. Differences in Antimicrobial Susceptibility of Bacteria between Years at 13DOF
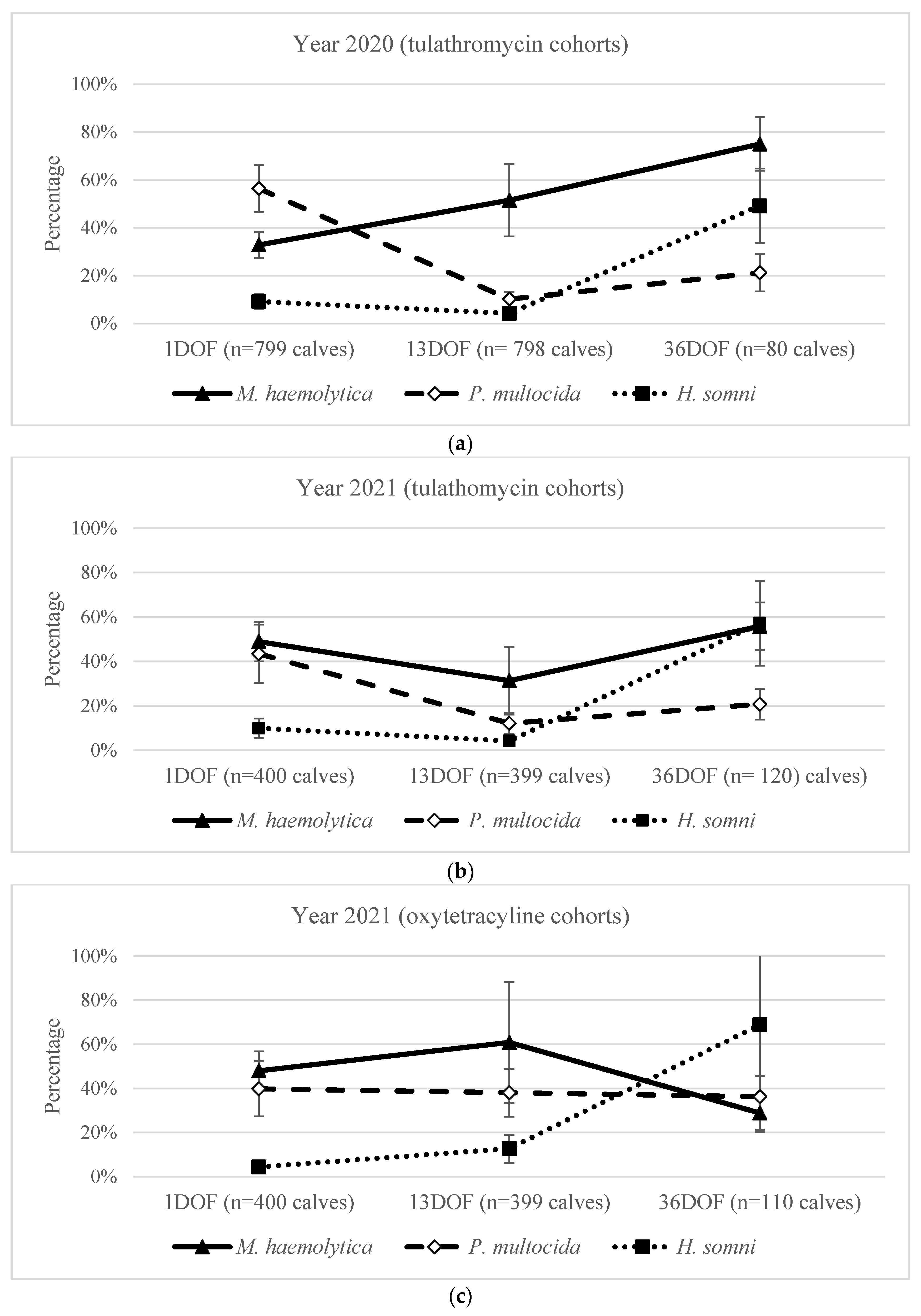
2.7. Within-Year Comparison of Bacterial and Antimicrobial Susceptibility from 1DOF to 36DOF
2.8. Pen-Level Clustering of Bacterial Recovery and Antimicrobial Resistance
2.9. Sample Size Estimates Generated from Simulation Models
3. Discussion
Study Limitations
4. Materials and Methods
4.1. Ethical Statement
4.2. Study Population
4.3. Calf Processing Procedure
4.4. Animal Housing and Management
4.5. Sampling Procedures
4.6. Bacteriology
4.7. Antimicrobial Susceptibility Testing
4.8. Treatment of Calves with BRD
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ives, S.E.; Richeson, J.T. Use of Antimicrobial Metaphylaxis for the Control of Bovine Respiratory Disease in High-Risk Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2015, 31, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Brault, S.A.; Hannon, S.J.; Gow, S.P.; Warr, B.N.; Withell, J.; Song, J.; Williams, C.M.; Otto, S.J.G.; Booker, C.W.; Morley, P.S. Antimicrobial Use on 36 Beef Feedlots in Western Canada: 2008–2012. Front. Vet. Sci. 2019, 6, 329. [Google Scholar] [CrossRef] [PubMed]
- Edwards, T.A. Control Methods for Bovine Respiratory Disease for Feedlot Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 273–284. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, A.M.; Hu, D.; Totton, S.C.; Scott, N.; Winder, C.B.; Wang, B.; Wang, C.; Glanville, J.; Wood, H.; White, B.; et al. A systematic review and network meta-analysis of bacterial and viral vaccines, administered at or near arrival at the feedlot, for control of bovine respiratory disease in beef cattle. Anim. Health Res. Rev. 2019, 20, 143–162. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.A.; Step, D.L.; Woolums, A.R. Bovine Respiratory Disease: Looking Back and Looking Forward, What Do We See? Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Nickell, J.S.; White, B.J. Metaphylactic Antimicrobial Therapy for Bovine Respiratory Disease in Stocker and Feedlot Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 285–301. [Google Scholar] [CrossRef] [PubMed]
- Aidara-Kane, A.; Angulo, F.J.; Conly, J.M.; Minato, Y.; Silbergeld, E.K.; McEwen, S.A.; Collignon, P.J.; Balkhy, H.; Collignon, P.; Conly, J.; et al. World Health Organization (WHO) guidelines on use of medically important antimicrobials in food-producing animals. Antimicrob. Resist. Infect. Control 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M. Assessing National Action Plans on Antimicrobial Resistance in Animal Production. What Lessons Can Be Drawn? OECD Food, Agriculture and Fisheries Papers, No. 153; OECD Publishing: Paris, France, 2021. [Google Scholar] [CrossRef]
- Klima, C.L.; Holman, D.B.; Cook, S.R.; Conrad, C.C.; Ralston, B.J.; Allan, N.; Anholt, R.M.; Niu, Y.D.; Stanford, K.; Hannon, S.J.; et al. Multidrug Resistance in Pasteurellaceae Associated With Bovine Respiratory Disease Mortalities in North America From 2011 to 2016. Front. Microbiol. 2020, 11, 606438. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.; Zaheer, R.; Klima, C.; McAllister, T.; Peters, D.; Niu, Y.D.; Ralston, B. Antimicrobial Resistance in Members of the Bacterial Bovine Respiratory Disease Complex Isolated from Lung Tissue of Cattle Mortalities Managed with or without the Use of Antimicrobials. Microorganisms 2020, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.L.; Sweeney, M.T. Antimicrobial Resistance in Bovine Respiratory Disease Pathogens: Measures, Trends, and Impact on Efficacy. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D.; Fulton, R.W.; Lehenbauer, T.W.; Step, D.L.; Confer, A.W. epidemiology of bovine respiratory disease: What is the evidence for predisposing factors? Can. Vet. J. = La Rev. Vet. Can. 2010, 51, 1095–1102. [Google Scholar]
- Snowder, G.D.; Van Vleck, L.D.; Cundiff, L.V.; Bennett, G.L. Bovine respiratory disease in feedlot cattle: Environmental, genetic, and economic factors. J. Anim. Sci. 2006, 84, 1999–2008. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, M.W.; Dargatz, D.A.; Wagner, B.A. Risk factors for initial respiratory disease in United States’ feedlots based on producer-collected daily morbidity counts. Can. Vet. J. 2008, 49, 373–378. [Google Scholar] [PubMed]
- Noyes, N.R.; Benedict, K.M.; Gow, S.P.; Booker, C.W.; Hannon, S.J.; McAllister, T.A.; Morley, P.S. Mannheimia haemolytica in feedlot cattle: Prevalence of recovery and associations with antimicrobial use, resistance, and health outcomes. J. Vet. Intern. Med. 2015, 29, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.J.; White, B.J.; Amrine, D.E.; Larson, R.L.; Theurer, M.E.; Szasz, J.I.; Bryant, T.C.; Waggoner, J.W. Evaluation of First Treatment Timing, Fatal Disease Onset, and Days from First Treatment to Death Associated with Bovine Respiratory Disease in Feedlot Cattle. Vet. Sci. 2023, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.A. Impact of disease on feedlot performance: A review. J. Anim. Sci. 1998, 76, 272–274. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, C.; Timsit, E.; Uddin, M.S.; Guan, L.L.; Alexander, T.W. Comparison of pathogenic bacteria in the upper and lower respiratory tracts of cattle either directly transported to a feedlot or co-mingled at auction markets prior to feedlot placement. Front. Vet. Sci. 2023, 9, 1026470. [Google Scholar] [CrossRef] [PubMed]
- McMullen, C.; Alexander, T.W.; Orsel, K.; Timsit, E. Progression of nasopharyngeal and tracheal bacterial microbiotas of feedlot cattle during development of bovine respiratory disease. Vet. Microbiol. 2020, 248, 108826. [Google Scholar] [CrossRef] [PubMed]
- McMullen, C.; Orsel, K.; Alexander, T.W.; van der Meer, F.; Plastow, G.; Timsit, E. Evolution of the nasopharyngeal bacterial microbiota of beef calves from spring processing to 40 days after feedlot arrival. Vet. Microbiol. 2018, 225, 139–148. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, A.M.; Hu, D.; Totton, S.C.; Scott, N.; Winder, C.B.; Wang, B.; Wang, C.; Glanville, J.; Wood, H.; White, B.; et al. A systematic review and network meta-analysis of injectable antibiotic options for the control of bovine respiratory disease in the first 45 days post arrival at the feedlot. Anim. Health Res. Rev. 2019, 20, 163–181. [Google Scholar] [CrossRef] [PubMed]
- Nobrega, D.; Andres-Lasheras, S.; Zaheer, R.; McAllister, T.; Homerosky, E.; Anholt, R.M.; Dorin, C. Prevalence, Risk Factors, and Antimicrobial Resistance Profile of Respiratory Pathogens Isolated From Suckling Beef Calves to Reprocessing at the Feedlot: A Longitudinal Study. Front. Vet. Sci. 2021, 8, 764701. [Google Scholar] [CrossRef] [PubMed]
- Holman, D.B.; Yang, W.; Alexander, T.W. Antibiotic treatment in feedlot cattle: A longitudinal study of the effect of oxytetracycline and tulathromycin on the fecal and nasopharyngeal microbiota. Microbiome 2019, 7, 86. [Google Scholar] [CrossRef] [PubMed]
- Timsit, E.; Workentine, M.; Schryvers, A.B.; Holman, D.B.; van Der Meer, F.; Alexander, T.W. Evolution of the nasopharyngeal microbiota of beef cattle from weaning to 40days after arrival at a feedlot. Vet. Microbiol. 2016, 187, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Loerch, S.C.; Fluharty, F.L. Physiological changes and digestive capabilities of newly received feedlot cattle. J. Anim. Sci. 1999, 77, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Anholt, R.M.; Klima, C.; Allan, N.; Matheson-Bird, H.; Schatz, C.; Ajitkumar, P.; Otto, S.J.; Peters, D.; Schmid, K.; Olson, M.; et al. Antimicrobial Susceptibility of Bacteria That Cause Bovine Respiratory Disease Complex in Alberta, Canada. Front. Vet. Sci. 2017, 4, 207. [Google Scholar] [CrossRef] [PubMed]
- Crosby, W.B.; Karisch, B.B.; Hiott, L.M.; Pinnell, L.J.; Pittman, A.; Frye, J.G.; Jackson, C.R.; Loy, J.D.; Epperson, W.B.; Blanton, J.; et al. Tulathromycin metaphylaxis increases nasopharyngeal isolation of multidrug resistant Mannheimia haemolytica in stocker heifers. Front. Vet. Sci. 2023, 10, 1256997. [Google Scholar] [CrossRef] [PubMed]
- Woolums, A.; Karisch, B.; Frye, J.; Epperson, W.; Smith, D.; Blanton, J.; Austin, F.; Kaplan, R.; Hiott, L.; Woodley, T.; et al. Multidrug resistant Mannheimia haemolytica isolated from high-risk beef stocker cattle after antimicrobial metaphylaxis and treatment for bovine respiratory disease. Vet. Microbiol. 2018, 221, 143. [Google Scholar] [CrossRef] [PubMed]
- Snyder, E.; Credille, B.; Berghaus, R.; Giguère, S. Prevalence of multi drug antimicrobial resistance in isolated from high-risk stocker cattle at arrival and two weeks after processing. J. Anim. Sci. 2017, 95, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Briggs, R.E.; Frank, G.H.; Purdy, C.W.; Zehr, E.S.; Loan, R.W. Rapid spread of a unique strain of Pasteurella haemolytica serotype 1 among transported calves. Am. J. Vet. Res. 1998, 59, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Holman, D.B.; Timsit, E.; Booker, C.W.; Alexander, T.W. Injectable antimicrobials in commercial feedlot cattle and their effect on the nasopharyngeal microbiota and antimicrobial resistance. Vet. Microbiol. 2018, 214, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Harner, J.; Boyer, W.; Davidson, J.; George, H.; Graber, R.; Murphy, J.P. Guidelines for Planning Cattle Feedlots. 2017. Available online: https://www.kcare.k-state.edu/pubs/livestock_management/Guidelines%20for%20Planning%20Cattle%20Feedlots%20MF3392.pdf (accessed on 5 August 2023).
- Pound-Maker. Feedlot Virtual Tour. Available online: https://pound-maker.ca/pages/feedlot-virtual-tour (accessed on 30 January 2024).
- Andrés-Lasheras, S.; Ha, R.; Zaheer, R.; Lee, C.; Booker, C.W.; Dorin, C.; Van Donkersgoed, J.; Deardon, R.; Gow, S.; Hannon, S.J.; et al. Prevalence and Risk Factors Associated With Antimicrobial Resistance in Bacteria Related to Bovine Respiratory Disease—A Broad Cross-Sectional Study of Beef Cattle at Entry Into Canadian Feedlots. Front. Vet. Sci. 2021, 8, 692646. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. Categorisation of Antibiotics in the European Union; EMA/CVMP/CHMP/682198/2017; European Medicines Agency: Amsterdam, The Netherlands, 2019. Available online: https://www.ema.europa.eu/en/documents/report/categorisation-antibiotics-european-union-answer-request-european-commission-updating-scientific-advice-impact-public-health-and-animal-health-use-antibiotics-animals_en.pdf (accessed on 12 June 2022).
- Barrett, J.R.; Innes, G.K.; Johnson, K.A.; Lhermie, G.; Ivanek, R.; Greiner Safi, A.; Lansing, D. Consumer perceptions of antimicrobial use in animal husbandry: A scoping review. PLoS ONE 2021, 16, e0261010. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Lasheras, S.; Jelinski, M.; Zaheer, R.; McAllister, T.A. Bovine Respiratory Disease: Conventional to Culture-Independent Approaches to Studying Antimicrobial Resistance in North America. Antibiotics 2022, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Erickson, N.E.; Ngeleka, M.G.; Lubbers, B.V.; Trokhymchuk, A. Changes in the rates of field isolation and antimicrobial susceptibility of bacterial pathogens collected from fall-placed feedlot steers between arrival at the feedlot and 90 to 120 days on feed. Bov. Pract. 2017, 51, 165–173. [Google Scholar] [CrossRef]
- Wennekamp, T.R.; Waldner, C.L.; Windeyer, M.C.; Larson, K.; Trokhymchuk, A.; Campbell, J.R. Antimicrobial resistance in bovine respiratory disease: Auction market- and ranch-raised calves. Can. Vet. J. 2022, 63, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; McMullen, C.; Timsit, E.; Hallewell, J.; Orsel, K.; van der Meer, F.; Yan, S.; Alexander, T.W. Genetic relatedness and antimicrobial resistance in respiratory bacteria from beef calves sampled from spring processing to 40 days after feedlot entry. Vet. Microbiol. 2020, 240, 108478. [Google Scholar] [CrossRef] [PubMed]
- Holman, D.B.; Timsit, E.; Alexander, T.W. The nasopharyngeal microbiota of feedlot cattle. Sci. Rep. 2015, 5, 15557. [Google Scholar] [CrossRef] [PubMed]
- Ribble, C.S.; Meek, A.H.; Janzen, E.D.; Guichon, P.T.; Jim, G.K. Effect of time of year, weather, and the pattern of auction market sales on fatal fibrinous pneumonia (shipping fever) in calves in a large feedlot in Alberta (1985–1988). Can. J. Vet. Res. 1995, 59, 167–172. [Google Scholar] [PubMed]
- Timsit, E.; Workentine, M.; van der Meer, F.; Alexander, T. Distinct bacterial metacommunities inhabit the upper and lower respiratory tracts of healthy feedlot cattle and those diagnosed with bronchopneumonia. Vet. Microbiol. 2018, 221, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Apley, M.D. Treatment of Calves with Bovine Respiratory Disease: Duration of Therapy and Posttreatment Intervals. Vet. Clin. N. Am. Food Anim. Pract. 2015, 31, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Hannon, S.J.; Brault, S.A.; Otto, S.J.G.; Morley, P.S.; McAllister, T.A.; Booker, C.W.; Gow, S.P. Feedlot Cattle Antimicrobial Use Surveillance Network: A Canadian Journey. Front. Vet. Sci. 2020, 7, 596042. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration (FDA). NARMS Now. Rockville, MD: U.S. Department of Health and Human Services. Available online: https://www.fda.gov/animal-veterinary/national-antimicrobial-resistance-monitoring-system/narms-now-integrated-data (accessed on 19 February 2022).
- Gow, S.; Bergen, R.; Booker, C.; Butters, A.; Dorin, C.; Dimmers, G.; Erickson, N.; Hannon, S.; Hendrick, S.; Ramsay, D.; et al. National surveillance of antimicrobial use and antimicrobial resistance in Canadian feedlots. In Proceedings of the Fifty-Fourth Annual Conference, American Association of Bovine Practitioners, Salt Lake City, UT, USA, 7–9 October 2021. [Google Scholar]
- Younes, J.A.; Ramsay, D.E.; Lacoste, S.; Deschner, D.; Hill, J.E.; Campbell, J.; Waldner, C.L. Changes in the phenotypic susceptibility of Mannheimia haemolytica isolates to macrolide antimicrobials during the early feeding period following metaphylactic tulathromycin use in western Canadian feedlot calves. Can. Vet. J. 2022, 63, 920–928. [Google Scholar] [PubMed]
- Timsit, E.; Christensen, H.; Bareille, N.; Seegers, H.; Bisgaard, M.; Assie, S. Transmission dynamics of Mannheimia haemolytica in newly-received beef bulls at fattening operations. Vet. Microbiol. 2013, 161, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Klima, C.L.; Alexander, T.W.; Hendrick, S.; McAllister, T.A. Characterization of Mannheimia haemolytica isolated from feedlot cattle that were healthy or treated for bovine respiratory disease. Can. J. Vet. Res. = Rev. Can. De Rech. Vet. 2014, 78, 38. [Google Scholar]
- Babcock, A.H.; White, B.J.; Dritz, S.S.; Thomson, D.U.; Renter, D.G. Feedlot health and performance effects associated with the timing of respiratory disease treatment1. J. Anim. Sci. 2009, 87, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Dohoo, I.R.; Martin, W.; Stryhn, H. Veterinary Epidemiologic Research, 2nd ed.; Charlotte, P.E.I., Ed.; VER, Inc.: Charlottetown, PE, Canada, 2014. [Google Scholar]
- Stevenson, M.A. Sample Size Estimation in Veterinary Epidemiologic Research. Front. Vet. Sci. 2021, 7, 539573. [Google Scholar] [CrossRef] [PubMed]
- Capik, S.F.; White, B.J.; Lubbers, B.V.; Apley, M.D.; DeDonder, K.D.; Larson, R.L.; Harhay, G.P.; Chitko-McKown, C.G.; Harhay, D.M.; Kalbfleisch, T.S.; et al. Comparison of the diagnostic performance of bacterial culture of nasopharyngeal swab and bronchoalveolar lavage fluid samples obtained from calves with bovine respiratory disease. Am. J. Vet. Res. 2017, 78, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Pass, D.A.; Thompson, R.G. Wide distribution of Pasteurella haemolytica type 1 over the nasal mucosa of cattle. Can. J. Comp. Med. 1971, 35, 181–186. [Google Scholar] [PubMed]
- Magwood, S.E.; Barnum, D.A.; Thomson, R.G. Nasal bacterial flora of calves in healthy and in pneumonia-prone herds. Can. J. Comp. Med. 1969, 33, 237–243. [Google Scholar] [PubMed]
- Doyle, D.; Credille, B.; Lehenbauer, T.W.; Berghaus, R.; Aly, S.S.; Champagne, J.; Blanchard, P.; Crossley, B.; Berghaus, L.; Cochran, S.; et al. Agreement Among 4 Sampling Methods to Identify Respiratory Pathogens in Dairy Calves with Acute Bovine Respiratory Disease. J. Vet. Intern. Med. 2017, 31, 954–959. [Google Scholar] [CrossRef] [PubMed]
- DeRosa, D.C.; Mechor, G.D.; Staats, J.J.; Chengappa, M.M.; Shryock, T.R. Comparison of Pasteurella spp. simultaneously isolated from nasal and transtracheal swabs from cattle with clinical signs of bovine respiratory disease. J. Clin. Microbiol. 2000, 38, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Godinho, K.S.; Sarasola, P.; Renoult, E.; Tilt, N.; Keane, S.; Windsor, G.D.; Rowan, T.G.; Sunderland, S.J. Use of deep nasopharyngeal swabs as a predictive diagnostic method for natural respiratory infections in calves. Vet. Rec. 2007, 160, 22–25. [Google Scholar] [CrossRef] [PubMed]
- McDaneld, T.G.; Kuehn, L.A.; Keele, J.W. Evaluating the microbiome of two sampling locations in the nasal cavity of cattle with bovine respiratory disease complex (BRDC). J. Anim. Sci. 2018, 96, 1281–1287. [Google Scholar] [CrossRef] [PubMed]
- McMullen, C.; Alexander, T.W.; Léguillette, R.; Workentine, M.; Timsit, E. Topography of the respiratory tract bacterial microbiota in cattle. Microbiome 2020, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Pardon, B.; Buczinski, S. Bovine Respiratory Disease Diagnosis: What Progress Has Been Made in Infectious Diagnosis? Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Limmathurotsakul, D.; Jamsen, K.; Arayawichanont, A.; Simpson, J.A.; White, L.J.; Lee, S.J.; Wuthiekanun, V.; Chantratita, N.; Cheng, A.; Day, N.P.J.; et al. Defining the true sensitivity of culture for the diagnosis of melioidosis using Bayesian latent class models. PLoS ONE 2010, 5, e12485. [Google Scholar] [CrossRef] [PubMed]
- Singer, R.S.; Johnson, W.O.; Jeffrey, J.S.; Chin, R.P.; Carpenter, T.E.; Atwill, E.R.; Hirsh, D.C. A statistical model for assessing sample size for bacterial colony selection: A case study of Escherichia coli and avian cellulitis. J. Vet. Diagn. Investig. 2000, 12, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Carter, H.F.; Wills, R.W.; Scott, M.A.; Thompson, A.C.; Singer, R.S.; Loy, J.D.; Karisch, B.B.; Epperson, W.B.; Woolums, A.R. Assessment of Diversity of Antimicrobial Resistance Phenotypes and Genotypes of Mannheimia haemolytica Isolates From Bovine Nasopharyngeal Swabs. Front. Vet. Sci. 2022, 9, 883389. [Google Scholar] [CrossRef] [PubMed]
- Loy, J.D.; Clawson, M.L. Rapid typing of Mannheimia haemolytica major genotypes 1 and 2 using MALDI-TOF mass spectrometry. J. Microbiol. Methods 2017, 136, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Capik, S.F.; White, B.J.; Lubbers, B.V.; Apley, M.D.; Mosier, D.A.; Larson, R.L.; Murray, R.W. Characterization of Mannheimia haemolytica in beef calves via nasopharyngeal culture and pulsed-field gel electrophoresis. J. Vet. Diagn. Investig. 2015, 27, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Maunsell, F.P.; Woolums, A.R.; Francoz, D.; Rosenbusch, R.F.; Step, D.L.; Wilson, D.J.; Janzen, E.D. Mycoplasma bovis Infections in Cattle. J. Vet. Intern. Med. 2011, 25, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Booker, C.W.; Abutarbush, S.M.; Morley, P.S.; Jim, G.K.; Pittman, T.J.; Schunicht, O.C.; Perrett, T.; Wildman, B.K.; Fenton, R.K.; Guichon, P.T.; et al. Microbiological and histopathological findings in cases of fatal bovine respiratory disease of feedlot cattle in Western Canada. Can. Vet. J. 2008, 49, 473–481. [Google Scholar] [PubMed]
- Sarchet, J.J.; Pollreisz, J.P.; Bechtol, D.T.; Blanding, M.R.; Saltman, R.L.; Taube, P.C. Limitations of bacterial culture, viral PCR, and tulathromycin susceptibility from upper respiratory tract samples in predicting clinical outcome of tulathromycin control or treatment of bovine respiratory disease in high-risk feeder heifers. PLoS ONE 2022, 17, e0247213. [Google Scholar] [CrossRef] [PubMed]
- National Farm Animal Council. Code of Practice for the Care and Handling of Beef Cattle; National Farm Animal Council: Lacombe, AB, Canada, 2013.
- Laboratory Standards Insitute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacterial Isolated from Animals, 6th ed.; Clinical and CLSI Supplement VET01S; Laboratory Standards Insitute: Wayne, PA, USA, 2023. [Google Scholar]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Editorial: Assessing the antimicrobial susceptibility of bacteria obtained from animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Step, D.L.; Krehbiel, C.R.; DePra, H.A.; Cranston, J.J.; Fulton, R.W.; Kirkpatrick, J.G.; Gill, D.R.; Payton, M.E.; Montelongo, M.A.; Confer, A.W. Effects of commingling beef calves from different sources and weaning protocols during a forty-two-day receiving period on performance and bovine respiratory disease. J. Anim. Sci. 2008, 86, 3146–3158. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Year | Pen | Unique Herds of Origin | Avg. Weight (kg) | Standard Dev. (kg) |
|---|---|---|---|---|
| 2020 | 1 | 34 | 257 | 13 |
| 2 | 81 | 243 | 10 | |
| 3 | 48 | 254 | 11 | |
| 4 | 43 | 253 | 13 | |
| 5 | 31 | 260 | 9 | |
| 6 | 38 | 262 | 11 | |
| 7 | 41 | 256 | 13 | |
| 8 | 30 | 239 | 13 | |
| Combined | 292 | 253 | 14 | |
| 2021 | 9 | 31 | 223 | 18 |
| 10 | 36 | 229 | 14 | |
| 11 | 33 | 222 | 14 | |
| 12 | 35 | 229 | 10 | |
| 13 | 23 | 220 | 18 | |
| 14 | 12 | 223 | 20 | |
| 15 | 38 | 228 | 13 | |
| 16 | 38 | 230 | 12 | |
| Combined | 208 | 225 | 15 |
| Metaphylaxis Drug | Pen | Sampling Time | No. Calves | Recovery Rates of BRD Pathogens from Calves | ||
|---|---|---|---|---|---|---|
| M. haemolytica | P. multocida | H. somni | ||||
| tulathromycin | 1 | 1DOF | 100 | 26% | 32% | 4% |
| 13DOF | 100 | 77% | 2% | 0% | ||
| 36DOF | 10 | 80% | 20% | 30% | ||
| tulathromycin | 2 | 1DOF | 100 | 35% | 47% | 3% |
| 13DOF | 100 | 84% | 7% | 1% | ||
| 36DOF | 10 | 70% | 10% | 50% | ||
| tulathromycin | 3 | 1DOF | 100 | 47% | 48% | 6% |
| 13DOF | 100 | 70% | 13% | 0% | ||
| 36DOF | 10 | 80% | 40% | 30% | ||
| tulathromycin | 4 | 1DOF | 100 | 47% | 55% | 4% |
| 13DOF | 100 | 80% | 21% | 4% | ||
| 36DOF | 10 | 80% | 20% | 60% | ||
| tulathromycin | 5 | 1DOF | 100 | 33% | 67% | 12% |
| 13DOF | 100 | 14% | 11% | 2% | ||
| 36DOF | 10 | 80% | 10% | 40% | ||
| tulathromycin | 6 | 1DOF | 99 | 33% | 63% | 7% |
| 13DOF | 99 | 9% | 8% | 5% | ||
| 36DOF | 10 | 70% | 20% | 30% | ||
| tulathromycin | 7 | 1DOF | 100 | 28% | 70% | 12% |
| 13DOF | 99 | 30% | 11% | 12% | ||
| 36DOF | 10 | 70% | 30% | 100% | ||
| tulathromycin | 8 | 1DOF | 100 | 15% | 70% | 29% |
| 13DOF | 100 | 57% | 8% | 11% | ||
| 36DOF | 10 | 70% | 20% | 50% | ||
| Metaphylaxis Drug | Pen | Sampling Time | No. Calves | Recovery Rates of BRD Pathogens from Calves | ||
|---|---|---|---|---|---|---|
| M. haemolytica | P. multocida | H. somni | ||||
| oxytetracycline | 9 | 1DOF | 100 | 60% | 23% | 6% |
| 13DOF | 100 | 61% | 35% | 10% | ||
| 36DOF | 30 | 33% | 33% | 70% | ||
| oxytetracycline | 10 | 1DOF | 100 | 49% | 29% | 0% |
| 13DOF | 100 | 52% | 60% | 5% | ||
| 36DOF | 30 | 13% | 57% | 73% | ||
| oxytetracycline | 11 | 1DOF | 100 | 40% | 35% | 4% |
| 13DOF | 100 | 47% | 18% | 19% | ||
| 37DOF | 30 | 47% | 20% | 70% | ||
| tulathromycin | 12 | 1DOF | 100 | 58% | 39% | 7% |
| 13DOF | 100 | 25% | 7% | 4% | ||
| 36DOF | 30 | 63% | 13% | 70% | ||
| tulathromycin | 13 | 1DOF | 100 | 47% | 29% | 13% |
| 13DOF | 100 | 17% | 14% | 21% | ||
| 36DOF | 30 | 67% | 27% | 53% | ||
| tulathromycin | 14 | 1DOF | 99 | 48% | 51% | 8% |
| 13DOF | 99 | 33% | 14% | 6% | ||
| 36DOF | 30 | 50% | 23% | 73% | ||
| tulathromycin | 15 | 1DOF | 100 | 43% | 56% | 9% |
| 13DOF | 99 | 44% | 12% | 9% | ||
| 36DOF | 30 | 43% | 20% | 40% | ||
| oxytetracycline | 16 | 1DOF | 100 | 43% | 73% | 7% |
| 12DOF | 99 | 61% | 41% | 13% | ||
| 30DOF | 20 | 20% | 35% | 60% | ||
| Bacteria | OR 2021 vs. 2020 | 95% CI | p-Value |
|---|---|---|---|
| M. haemolytica | 1.9 | 1.3, 2.8 | <0.001 |
| P. multocida | 0.6 | 0.3, 0.97 | 0.04 |
| H. somni | 0.7 | 0.4, 1.5 | 0.42 |
| Bacteria | Year/Metaphylaxis Comparison | OR | 95% CI | p-Value |
|---|---|---|---|---|
| M. haemolytica | 2021/tulathromycin vs. 2020/tulathromycin | 0.4 | 0.1, 1.3 | 0.12 |
| 2021/oxytetracycline vs. 2020/tulathromycin | 1.2 | 0.3, 4.0 | 0.82 | |
| 2021/oxytetracycline vs. 2021/tulathromycin | 3.1 | 0.7, 13 | 0.12 | |
| P. multocida | 2021/tulathromycin vs. 2020/tulathromycin | 1.2 | 0.6, 2.4 | 0.55 |
| 2021/oxytetracycline vs. 2020/tulathromycin | 5.5 | 2.9, 9.3 | <0.001 | |
| 2021/oxytetracycline vs. 2021/tulathromycin | 4.4 | 2.1, 9.3 | <0.001 | |
| H. somni | 2021/tulathromycin vs. 2020/tulathromycin | 2.6 | 0.6, 2.4 | 0.06 |
| 2021/oxytetracycline vs. 2020/tulathromycin | 3.3 | 1.2, 3.7 | 0.018 | |
| 2021/oxytetracycline vs. 2021/tulathromycin | 1.3 | 0.4, 3.7 | 0.67 |
| Bacteria | Year/Metaphylaxis | DOF Comparison | OR | 95% CI | p-Value |
|---|---|---|---|---|---|
| M. haemolytica | 2020/tulathromycin | 13DOF vs. 1DOF | 2.4 | 1.9, 2.9 | <0.001 |
| 36DOF vs. 1DOF | 6.8 | 4.0, 12 | <0.001 | ||
| 36DOF vs. 13DOF | 2.8 | 1.7, 4.8 | <0.001 | ||
| 2021/tulathromycin | 13DOF vs. 1DOF | 0.5 | 0.3, 0.6 | <0.001 | |
| 36DOF vs. 1DOF | 1.3 | 0.9, 1.9 | 0.19 | ||
| 36DOF vs. 13DOF | 2.9 | 1.9, 4.3 | <0.001 | ||
| 2021/oxytetracycline | 13DOF vs. 1DOF | 1.3 | 1.0, 1.7 | 0.041 | |
| 36DOF vs. 1DOF | 0.5 | 0.3, 0.7 | <0.001 | ||
| 36DOF vs. 13DOF | 0.3 | 0.2, 0.5 | <0.001 | ||
| P. multocida | 2020/tulathromycin | 13DOF vs. 1DOF | 0.1 | 0.1, 0.1 | <0.001 |
| 36DOF vs. 1DOF | 0.2 | 0.1, 0.3 | <0.001 | ||
| 36DOF vs. 13DOF | 2.3 | 1.3, 4 | 0.004 | ||
| 2021/tulathromycin | 13DOF vs. 1DOF | 0.2 | 0.1, 0.2 | <0.001 | |
| 36DOF vs. 1DOF | 0.3 | 0.2, 0.5 | <0.001 | ||
| 36DOF vs. 13DOF | 1.9 | 1.2, 3.2 | 0.01 | ||
| 2021/oxytetracycline | 13DOF vs. 1DOF | 0.9 | 0.7, 1.2 | 0.65 | |
| 36DOF vs. 1DOF | 0.9 | 0.6, 1.3 | 0.55 | ||
| 36DOF vs. 13DOF | 0.9 | 0.6, 1.4 | 0.75 | ||
| H. somni | 2020/tulathromycin | 13DOF vs. 1DOF | 0.5 | 0.3, 0.7 | <0.001 |
| 36DOF vs. 1DOF | 9.8 | 5.7, 17 | <0.001 | ||
| 36DOF vs. 13DOF | 22 | 12, 40 | <0.001 | ||
| 2021/tulathromycin | 13DOF vs. 1DOF | 1.1 | 0.7, 1.7 | 0.70 | |
| 36DOF vs. 1DOF | 13 | 7.7, 22 | <0.001 | ||
| 36DOF vs. 13DOF | 12 | 7.1, 20 | <0.001 | ||
| 2021/oxytetracycline | 13DOF vs. 1DOF | 2.7 | 1.6, 4.6 | <0.001 | |
| 36DOF vs. 1DOF | 44 | 23, 84 | <0.001 | ||
| 36DOF vs. 13DOF | 16 | 9.3, 27 | <0.001 |
| Number (%) of Calves with Bacterial Co-Isolation Pattern | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Meta. 1 | Time Point | No. Calves | Neg. Culture | MH | PM | HS | MH + PM | MH + HS | PM + HS | MH + PM + HS |
| 2020 | Tula | 1DOF | 799 | 183 (23%) | 133 (17%) | 286 (36%) | 26 (3%) | 120 (15%) | 6 (0.8%) | 40 (5%) | 5 (0.6%) |
| 13DOF | 798 | 322 (40%) | 366 (46%) | 32 (4%) | 19 (2%) | 43 (5%) | 10 (1%) | 4 (0.5%) | 2 (0.3%) | ||
| 2021 | Tula | 1DOF | 400 | 83 (21%) | 117 (29%) | 94 (24%) | 19 (5%) | 69 (17%) | 7 (12%) | 8 (2%) | 3 (0.8%) |
| 13DOF | 399 | 219 (55%) | 102 (26%) | 26 (6.5%) | 27 (7%) | 12 (3%) | 4 (1%) | 8 (2%) | 1 (0.3%) | ||
| 2021 | Oxy | 1DOF | 400 | 102 (26%) | 131 (33%) | 96 (24%) | 5 (1%) | 54 (14%) | 2 (0.5%) | 5 (1%) | 5 (1%) |
| 13DOF | 399 | 84 (21%) | 133 (33%) | 71 (18%) | 13 (3%) | 64 (16%) | 15 (4%) | 11 (3%) | 8 (2%) | ||
| All Years | All Groups | 1DOF | 1599 | 368 (23%) | 381 (24%) | 476 (30%) | 50 (3%) | 243 (15%) | 15 (0.9%) | 53 (3%) | 13 (0.8%) |
| 13DOF | 1596 | 625 (39%) | 601 (38%) | 129 (8%) | 59 (3%) | 119 (8%) | 29 (2%) | 23 (1%) | 11 (0.7%) | ||
| Time Point | No. Calves | Bacteria 2 | Number (%) of Calves with Isolates Resistant to Select Antimicrobials 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AMP | DANO | FLOR | SPECT | TET | GAM | TILD | TILM | TUL | |||
| 1DOF | 799 | MH | 1 (0.1%) | 0 | 0 | 0 | 0 | 1 (0.1%) | 0 | 0 | 1 (0.1%) |
| 799 | PM | 15 (2%) | 0 | 0 | 31 (4%) | 28 (4%) | 0 | 0 | NI | 0 | |
| 799 | HS | 0 | NI | 0 | 1 (0.1%) | 0 | 0 | 4 (0.5%) | NI | 6 (0.8%) | |
| 13DOF | 798 | MH | 2 (0.3%) | 0 | 1 (0.1%) | 0 | 1 (0.1%) | 351 (44%) | 3 (0.4%) | 25 (3%) | 341 (43%) |
| 798 | PM | 7 (0.9%) | 0 | 0 | 10 (1%) | 8 (1%) | 1 (0.1%) | 1 (0.1%) | NI | 1 (0.1%) | |
| 798 | HS | 0 | NI | 0 | 0 | 0 | 1 (0.1%) | 2 (0.3%) | NI | 2 (0.3%) | |
| 36DOF | 80 | MH | 2 (2.5%) | 0 | 0 | 0 | 1 (1.3%) | 44 (55%) | 1 (1.3%) | 7 (8.8%) | 40 (50%) |
| 80 | PM | 0 | 0 | 0 | 3 (3.8%) | 3 (3.8%) | 0 | 0 | NI | 0 | |
| 80 | HS | 0 | NI | 0 | 0 | 0 | 0 | 0 | NI | 0 | |
| Number (%) of Calves with Isolates Resistant to Select Antimicrobials 1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Meta. 2 | Time Point | No. Calves | Bacteria 3 | AMP | PEN | DANO | SPECT | TET | GAM | TILD | TILM | TUL |
| Tula | 1DOF | 399 | MH | 2 (0.5%) | 0 | 2 (0.5%) | 0 | 1 (0.3%) | 0 | 1 (0.3%) | 2 (0.5%) | 0 |
| 399 | PM | 9 (2%) | 0 | 0 | 5 (1%) | 3 (0.8%) | 0 | 0 | NI | 0 | ||
| 399 | HS | 0 | 0 | NI | 1 (0.3%) | 0 | 0 | 0 | NI | 0 | ||
| 13DOF | 399 | MH | 0 | 0 | 0 | 0 | 18 (5%) | 27 (7%) | 38 (10%) | 43 (11%) | 27 (7%) | |
| 399 | PM | 6 (2%) | 0 | 0 | 0 | 0 | 0 | 0 | NI | 0 | ||
| 399 | HS | 1 (0.3%) | 0 | NI | 1 (0.3%) | 1 (0.3%) | 0 | 0 | NI | 0 | ||
| 36DOF | 119 | MH | 1 (0.8%) | 0 | 0 | 0 | 23 (19.3%) | 10 (8.4%) | 12 (10.1%) | 12 (10.1%) | 10 (8.4%) | |
| 119 | PM | 0 | 0 | 0 | 0 | 0 | 0 | 0 | NI | 0 | ||
| 119 | HS | 3 (2.5%) | 1 (0.8%) | 0 | 0 | 6 (5.0%) | 0 | 0 | NI | 0 | ||
| Oxy | 1DOF | 399 | MH | 0 | 1 (0.3%) | 0 | 0 | 1 (0.3%) | 0 | 0 | 0 | 0 |
| 399 | PM | 8 (2%) | 0 | 0 | 11 (3%) | 9 (2%) | 0 | 0 | NI | 0 | ||
| 399 | HS | 0 | 0 | NI | 0 | 1 (0.3%) | 0 | 0 | NI | 0 | ||
| 13DOF | 398 | MH | 1 (0.3%) | 2 (0.5%) | 0 (0%) | 0 | 3 (0.8%) | 3 (0.8%) | 3 (0.8%) | 3 (0.8%) | 3 (0.8%) | |
| 398 | PM | 3 (0.8%) | 0 | 0 (0%) | 53 (13%) | 52 (13%) | 0 | 0 | NI | 0 | ||
| 398 | HS | 0 | 0 | NI | 0 | 14 (4%) | 0 | 0 | NI | 0 | ||
| 36DOF | 110 | MH | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 110 | PM | 2 (1.8%) | 1 (0.9%) | 0 | 15 (13.6%) | 17 (15.5%) | 0 | 0 | NI | 0 | ||
| 110 | HS | 1 (0.9%) | 0 | NI | 0 | 11 (10%) | 0 | 0 | NI | 0 | ||
| AMR Outcome of Interest | Pairwise Comparison of Year/Metaphylaxis | OR | 95% CI | p-Value |
|---|---|---|---|---|
| M. haemolytica tulathromycin | 2021/tulathromycin vs. 2020/tulathromycin | 0.1 | 0.03, 0.7 | 0.02 |
| 2021/oxytetracycline vs. 2020/tulathromycin | 0.06 | 0.01, 0.4 | 0.004 | |
| 2021/oxytetracycline vs. 2021/tulathromycin | 0.4 | 0.05, 3.2 | 0.39 | |
| M. haemolytica gamithromycin | 2021/tulathromycin vs. 2020/tulathromycin | 0.1 | 0.03, 0.7 | 0.018 |
| 2021/oxytetracycline vs. 2020/tulathromycin | 0.06 | 0.01, 0.4 | 0.004 | |
| 2021/oxytetracycline vs. 2021/tulathromycin | 0.4 | 0.05, 3.1 | 0.39 | |
| P. multocida tetracycline * | 2021/tulathromycin vs. 2020/tulathromycin ** | 0.2 | 0, 0.90 | 0.08 |
| 2021/oxytetracycline vs. 2020/tulathromycin | 15 | 6.9, 37 | <0.0001 | |
| 2021/oxytetracycline vs. 2021/tulathromycin ** | 83 | 19, ∞ | <0.0001 |
| AMR Outcome of Interest | Year/Metaphylaxis | DOF Comparison | OR | 95% CI | p-Value |
|---|---|---|---|---|---|
| Tulathromycin | 2020/tulathromycin | 13DOF vs. 1DOF | 151 | 36, 638 | <0.001 |
| 36DOF vs. 1DOF | 206 | 46, 915 | <0.001 | ||
| 36DOF vs. 13DOF | 1.4 | 0.9, 2.1 | 0.14 | ||
| 2021/tulathromycin * | 13DOF vs. 1DOF ** | 41 | 9.1, ∞ | <0.001 | |
| 36DOF vs. 1DOF ** | 50 | 10, ∞ | <0.001 | ||
| 36DOF vs. 13DOF | 1.3 | 0.5, 3.4 | 0.67 | ||
| 2021/oxytetracycline | 13DOF vs. 1DOF ** | 3.9 | 0.6, ∞ | 0.25 | |
| 36DOF vs. 1DOF | · | · | · | ||
| 36DOF vs. 13DOF ** | 0.9 | 0, 6.2 | 0.96 | ||
| Gamithromycin | 2020/tulathromycin | 13DOF vs. 1DOF | 160 | 38, 672 | <0.001 |
| 36DOF vs. 1DOF | 260 | 58, 1151 | <0.001 | ||
| 36DOF vs. 13DOF | 1.6 | 1.1, 2.5 | 0.02 | ||
| 2021/tulathromycin * | 13DOF vs. 1DOF ** | 41 | 9.1, ∞ | <0.001 | |
| 36DOF vs. 1DOF ** | 50 | 10, ∞ | <0.001 | ||
| 36DOF vs. 13DOF | 1.3 | 0.5, 3.4 | 0.66 | ||
| 2021/oxytetracyclin | 13DOF vs. 1DOF ** | 3.9 | 0.6, ∞ | 0.25 | |
| 36DOF vs. 1DOF | · | · | · | ||
| 36DOF vs. 13DOF ** | 0.90 | 0, 6.2 | 0.96 | ||
| Tildipirosin | 2020/tulathromycin * | 13DOF vs. 1DOF ** | 3.9 | 0.6, ∞ | 0.25 |
| 36DOF vs. 1DOF ** | 9.9 | 0.5, ∞ | 0.18 | ||
| 36DOF vs. 13DOF | 3.3 | 0.06, 42 | 0.64 | ||
| 2021/tulathromycin | 13DOF vs. 1DOF | 10 | 3.0, 34 | <0.001 | |
| 36DOF vs. 1DOF | 11 | 3.0, 37 | <0.001 | ||
| 36DOF vs. 13DOF | 1.0 | 0.7, 1.6 | 0.85 | ||
| 2021/oxytetracycline * | 13DOF vs. 1DOF ** | 3.9 | 0.6, ∞ | 0.12 | |
| 36DOF vs. 1DOF | · | · | · | ||
| 36DOF vs. 13DOF ** | 0.9 | 0, 6.2 | 0.48 | ||
| Tilmicosin | 2020/tulathromycin * | 13DOF vs. 1DOF ** | 37 | 8.1, ∞ | <0.001 |
| 36DOF vs. 1DOF ** | 103 | 20, ∞ | <0.001 | ||
| 36DOF vs. 13DOF | 3.0 | 1.0, 7.4 | 0.04 | ||
| 2021/tulathromycin | 13DOF vs. 1DOF | 6.6 | 2.9, 15 | <0.001 | |
| 36DOF vs. 1DOF | 6.4 | 2.6, 16 | <0.001 | ||
| 36DOF vs. 13DOF | 1.0 | 0.6, 1.5 | 0.84 | ||
| 2021/oxytetracycline * | 13DOF vs. 1DOF ** | 3.9 | 0.6, ∞ | 0.25 | |
| 36DOF vs. 1DOF | · | · | · | ||
| 36DOF vs. 13DOF ** | 0.9 | 0, 6.2 | 0.96 | ||
| Tetracycline | 2020/tulathromycin * | 13DOF vs. 1DOF ** | 1.0 | 0.05, ∞ | 1.00 |
| 36DOF vs. 1DOF ** | 10 | 0.5, ∞ | 0.18 | ||
| 36DOF vs. 13DOF | 10 | 0.1, 1000 | 0.35 | ||
| 2021/tulathromycin | 13DOF vs. 1DOF | 7.2 | 1.9, 27 | 0.001 | |
| 36DOF vs. 1DOF | 27 | 6.3, 115 | <0.001 | ||
| 36DOF vs. 13DOF | 3.7 | 2, 7 | <0.001 | ||
| 2021/oxytetracycline * | 13DOF vs. 1DOF | 3.0 | 0.2, 159 | 0.62 | |
| 36DOF vs. 1DOF ** | 3.6 | 0, 69 | 1.57 | ||
| 36DOF vs. 13DOF ** | 0.9 | 0, 6.2 | 0.96 |
| AMR Outcome of Interest | Year/Metaphylaxis | DOF Comparison | OR | 95% CI | p-Value |
|---|---|---|---|---|---|
| Tetracycline | 2020/tulathromycin | 13DOF vs. 1DOF | 0.4 | 0.2, 0.7 | 0.001 |
| 36DOF vs. 1DOF | 1.1 | 0.4, 2.7 | 0.83 | ||
| 36DOF vs. 13DOF | 2.9 | 1, 8.2 | 0.052 | ||
| 2021/tulathromycin * | 13DOF vs. 1DOF ** | 0.3 | 0, 1.7 | 0.25 | |
| 36DOF vs. 1DOF ** | 0.9 | 0, 5.8 | 0.91 | ||
| 36DOF vs. 13DOF | · | · | · | ||
| 2021/oxytetracycline | 13DOF vs. 1DOF | 5.1 | 2.8, 9.3 | ≤0.001 | |
| 36DOF vs. 1DOF | 6.0 | 2.9, 12.6 | ≤0.001 | ||
| 36DOF vs. 13DOF | 1.2 | 0.7, 1.9 | 0.50 | ||
| Spectinomycin | 2020/tulathromycin | 13DOF vs. 1DOF | 0.41 | 0.2, 0.7 | 0.001 |
| 36DOF vs. 1DOF | 1.02 | 0.4, 2.6 | 0.97 | ||
| 36DOF vs. 13DOF | 2.5 | 0.9, 6.9 | 0.084 | ||
| 2021/tulathromycin * | 13DOF vs. 1DOF ** | 0.1 | 0, 0.8 | 0.06 | |
| 36DOF vs. 1DOF ** | 0.5 | 0, 2.8 | 0.54 | ||
| 36DOF vs. 13DOF | · | · | · | ||
| 2021/oxytetracycline | 13DOF vs. 1DOF | 4.3 | 2.5, 7.5 | ≤0.001 | |
| 36DOF vs. 1DOF | 4.3 | 2.2, 8.3 | ≤0.001 | ||
| 36DOF vs. 13DOF | 1.0 | 0.6, 1.6 | 0.97 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abi Younes, J.N.; Campbell, J.R.; Otto, S.J.G.; Gow, S.P.; Woolums, A.R.; Jelinski, M.; Lacoste, S.; Waldner, C.L. Variation in Pen-Level Prevalence of BRD Bacterial Pathogens and Antimicrobial Resistance Following Feedlot Arrival in Beef Calves. Antibiotics 2024, 13, 322. https://doi.org/10.3390/antibiotics13040322
Abi Younes JN, Campbell JR, Otto SJG, Gow SP, Woolums AR, Jelinski M, Lacoste S, Waldner CL. Variation in Pen-Level Prevalence of BRD Bacterial Pathogens and Antimicrobial Resistance Following Feedlot Arrival in Beef Calves. Antibiotics. 2024; 13(4):322. https://doi.org/10.3390/antibiotics13040322
Chicago/Turabian StyleAbi Younes, Jennifer N., John R. Campbell, Simon J. G. Otto, Sheryl P. Gow, Amelia R. Woolums, Murray Jelinski, Stacey Lacoste, and Cheryl L. Waldner. 2024. "Variation in Pen-Level Prevalence of BRD Bacterial Pathogens and Antimicrobial Resistance Following Feedlot Arrival in Beef Calves" Antibiotics 13, no. 4: 322. https://doi.org/10.3390/antibiotics13040322