
Jugiones A–D: Antibacterial Xanthone–Anthraquinone Heterodimers from Australian Soil-Derived Penicillium shearii CMB-STF067



Abstract
:1. Introduction
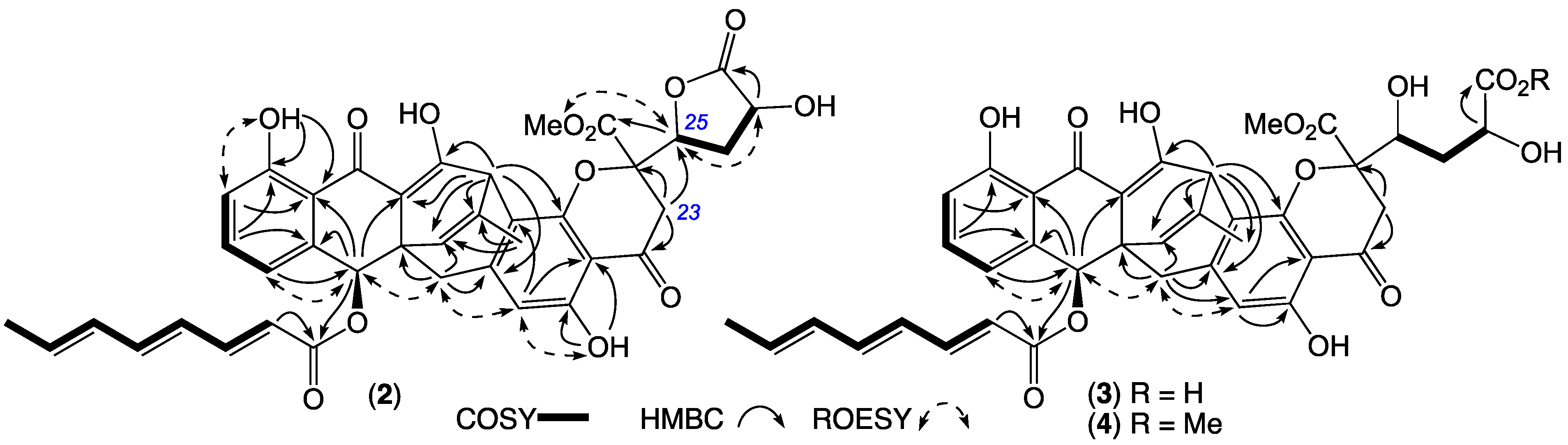
2. Results and Discussion
3. Materials and Methods
3.1. Collection and Isolation of CMB-STF067
3.2. Taxonomic Identification of CMB-STF067
3.3. UPLC-DAD Profiling of MATRIX Extracts
3.4. GNPS Molecular Network Profiling of MATRIX Extracts
3.5. Scaled-Up Cultivation and Chemical Fractionation of CMB-STF067
3.6. Antibacterial Assay
3.7. Antifungal Assay
3.8. Cytotoxic Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aron, A.T.; Gentry, E.C.; McPhail, K.L.; Nothias, L.-F.; Nothias-Esposito, M.; Bouslimani, A.; Petras, D.; Gauglitz, J.M.; Sikora, N.; Vargas, F.; et al. Reproducible Molecular Networking of Untargeted Mass Spectrometry Data Using GNPS. Nat. Protoc. 2020, 15, 1954–1991. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed]
- Pinedo-Rivilla, C.; Aleu, J.; Durán-Patrón, R. Cryptic Metabolites from Marine-Derived Microorganisms Using OSMAC and Epigenetic Approaches. Mar. Drugs 2022, 20, 84. [Google Scholar] [CrossRef]
- Tomm, H.A.; Ucciferri, L.; Ross, A.C. Advances in Microbial Culturing Conditions to Activate Silent Biosynthetic Gene Clusters for Novel Metabolite Production. J. Ind. Microbiol. Biotechnol. 2019, 46, 1381–1400. [Google Scholar] [CrossRef] [PubMed]
- Salim, A.A.; Khalil, Z.G.; Elbanna, A.H.; Wu, T.; Capon, R.J. Methods in Microbial Biodiscovery. Mar. Drugs 2021, 19, 503. [Google Scholar] [CrossRef]
- Kankanamge, S.; Khalil, Z.G.; Sritharan, T.; Capon, R.J. Noonindoles G–L: Indole Diterpene Glycosides from the Australian Marine-Derived Fungus Aspergillus noonimiae CMB-M0339. J. Nat. Prod. 2023, 86, 508–516. [Google Scholar] [CrossRef]
- Agampodi Dewa, A.; Khalil, Z.G.; Elbanna, A.H.; Capon, R.J. Chrysosporazines Revisited: Regioisomeric Phenylpropanoid Piperazine P-Glycoprotein Inhibitors from Australian Marine Fish-Derived Fungi. Molecules 2022, 27, 3172. [Google Scholar] [CrossRef]
- Salim, A.A.; Hussein, W.M.; Dewapriya, P.; Hoang, H.N.; Zhou, Y.; Samarasekera, K.; Khalil, Z.G.; Fairlie, D.P.; Capon, R.J. Talarolides Revisited: Cyclic Heptapeptides from an Australian Marine Tunicate-Associated Fungus, Talaromyces sp. CMB-TU011. Mar. Drugs 2023, 21, 487. [Google Scholar] [CrossRef]
- Sritharan, T.; Salim, A.A.; Capon, R.J. Miniaturized Cultivation Profiling Returns Indole-Sesquiterpene Michael Adducts from an Australian Soil-Derived Aspergillus terreus CMB-SWF012. J. Nat. Prod. 2023, 86, 2703–2709. [Google Scholar] [CrossRef]
- Ueda, J.-Y.; Takagi, M.; Shin-Ya, K. New Xanthoquinodin-Like Compounds, JBIR-97,-98 and-99, Obtained from Marine Sponge-Derived Fungus Tritirachium sp. SpB081112MEf2. J. Antibiot. 2010, 63, 615–618. [Google Scholar] [CrossRef]
- Wu, H.; Liu, J.; Duan, N.; Han, R.; Zhang, X.; Leng, X.; Liu, W.; Han, L.; Li, X.; Xing, S. Isolation and Purification of an Antibiotic Polyketide JBIR-99 from the Marine Fungus Meyerozyma Guilliermondii by High-Speed Counter-Current Chromatography. Int. J. Pharm. Chem. 2021, 7, 52–57. [Google Scholar] [CrossRef]
- Wu, B.; Wiese, J.; Wenzel-Storjohann, A.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Engyodontochones, Antibiotic Polyketides from the Marine Fungus Engyodontium album Strain LF069. Chem. Eur. J. 2016, 22, 7452–7462. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Marin-Felix, Y.; Surup, F.; Stchigel, A.M.; Stadler, M. Seven New Cytotoxic and Antimicrobial Xanthoquinodins from Jugulospora vestita. J. Fungi 2020, 6, 188. [Google Scholar] [CrossRef] [PubMed]
- Capon, R.J. Extracting Value: Mechanistic Insights into the Formation of Natural Product Artifacts—Case Studies in Marine Natural Products. Nat. Prod. Rep. 2020, 37, 55–79. [Google Scholar] [CrossRef] [PubMed]
- Tabata, N.; Tomoda, H.; Matsuzaki, K.; Omura, S. Structure and Biosynthesis of Xanthoquinodins, Anticoccidial Antibiotics. J. Am. Chem. Soc. 1993, 115, 8558–8564. [Google Scholar] [CrossRef]
- Milat, M.; Prangé, T.; Ducrot, P.H.; Tabet, J.; Einhorn, J.; Blein, J.; Lallemand, J. Structures of the Beticolins, the Yellow Toxins Produced by Cercospora beticola. J. Am. Chem Soc. 1992, 114, 1478–1479. [Google Scholar] [CrossRef]
- Jalal, M.A.; Hossain, M.B.; Robeson, D.J.; Van der Helm, D. Cercospora Beticola Phytotoxins: Cebetins That Are Photoactive, Magnesium Ion-Binding, Chlorinated Anthraquinone-Xanthone Conjugates. J. Am. Chem. Soc. 1992, 114, 5967–5971. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Tabata, N.; Tomoda, H.; Iwai, Y.; Tanaka, H.; Ōmura, S. The Structure of Xanthoquinodin A1, a Novel Anticoccidial Antibiotic Having a New Xanthone-Anthraquinone Conjugate System. Tetrahedron Lett. 1993, 34, 8251–8254. [Google Scholar] [CrossRef]
- Tabata, N.; Tomoda, H.; Iwai, Y.; Omura, S. Xanthoquinodin B3, a New Anticoccidial Agent Produced by Humicola sp. FO-888. J. Antibiot. 1996, 49, 267–271. [Google Scholar] [CrossRef]
- Chen, G.-D.; Chen, Y.; Gao, H.; Shen, L.-Q.; Wu, Y.; Li, X.-X.; Li, Y.; Guo, L.-D.; Cen, Y.-Z.; Yao, X.-S. Xanthoquinodins from the Endolichenic Fungal Strain Chaetomium elatum. J. Nat. Prod. 2013, 76, 702–709. [Google Scholar] [CrossRef]
- Sadorn, K.; Saepua, S.; Boonyuen, N.; Boonruangprapa, T.; Rachtawee, P.; Pittayakhajonwut, P. Antimicrobial Activity and Cytotoxicity of Xanthoquinodin Analogs from the Fungus Cytospora eugeniae BCC42696. Phytochemistry 2018, 151, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Tantapakul, C.; Promgool, T.; Kanokmedhakul, K.; Soytong, K.; Song, J.; Hadsadee, S.; Jungsuttiwong, S.; Kanokmedhakul, S. Bioactive Xanthoquinodins and Epipolythiodioxopiperazines from Chaetomium globosum 7s-1, an Endophytic Fungus Isolated from Rhapis cochinchinensis (Lour.) Mart. Nat. Prod. Res. 2020, 34, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Collins, J.E.; Hulverson, M.A.; Aguila, L.K.T.; Kim, C.M.; Wendt, K.L.; Chakrabarti, D.; Ojo, K.K.; Wood, G.E.; Van Voorhis, W.C.; et al. Appraisal of Fungus-Derived Xanthoquinodins as Broad-Spectrum Anti-Infectives Targeting Phylogenetically Diverse Human Pathogens. J. Nat. Prod. 2023, 86, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Palasarn, S.; Auncharoen, P.; Komwijit, S.; Gareth Jones, E.B. Acremoxanthones a and B, Novel Antibiotic Polyketides from the Fungus Acremonium sp. Bcc 31806. Tetrahedron Lett. 2009, 50, 284–287. [Google Scholar] [CrossRef]
- Ayers, S.; Graf, T.N.; Adcock, A.F.; Kroll, D.J.; Shen, Q.; Swanson, S.M.; Matthew, S.; Carcache de Blanco, E.J.; Wani, M.C.; Darveaux, B.A.; et al. Cytotoxic Xanthone–Anthraquinone Heterodimers from an Unidentified Fungus of the Order Hypocreales (MSX 17022). J. Antibiot. 2012, 65, 3–8. [Google Scholar] [CrossRef]
- Meléndez-González, C.; Murià-González, M.J.; Anaya, A.L.; Hernández-Bautista, B.E.; Hernández-Ortega, S.; González, M.C.; Glenn, A.E.; Hanlin, R.T.; Macías-Rubalcava, M.L. Acremoxanthone E, a Novel Member of Heterodimeric Polyketides with a Bicyclo[3.2.2]Nonene Ring, Produced by Acremonium camptosporum W. Gams (Clavicipitaceae) Endophytic Fungus. Chem. Biodivers. 2015, 12, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Intaraudom, C.; Bunbamrung, N.; Dramae, A.; Boonyuen, N.; Komwijit, S.; Rachtawee, P.; Pittayakhajonwut, P. Acremonidins F–H and Acremoxanthones F–G, Antimicrobial Substances from the Insect Fungus Verticillium sp. BCC33181. Tetrahedron 2016, 72, 1415–1421. [Google Scholar] [CrossRef]
- He, H.; Bigelis, R.; Solum, E.H.; Greenstein, M.; Carter, G.T. Acremonidins, New Polyketide-Derived Antibiotics Produced by Acremonium sp., LL-Cyan 416. J. Antibiot. 2003, 56, 923–930. [Google Scholar] [CrossRef]
- Adusumilli, R.; Mallick, P. Data Conversion with Proteowizard Msconvert. In Proteomics: Methods and Protocols; Comai, L., Katz, J.E., Mallick, P., Eds.; Springer: New York, NY, USA, 2017; pp. 339–368. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (1) δH, Mult (J in Hz) | (2) δH, Mult (J in Hz) | (3) δH, Mult (J in Hz) | (4) δH, Mult (J in Hz) | |
|---|---|---|---|---|
| 2 | 6.97, dd (8.6, 0.9) | 6.97, dd (8.4, 0.7) | 6.94, d (8.4) | 6.95, d (8.3) |
| 3 | 7.45, dd (7.5, 8.6) | 7.44, dd (8.4, 7.3) | 7.48, dd (8.4, 7.3) | 7.49, dd (8.3, 7.2) |
| 4 | 7.09, dd (7.5, 0.9) | 7.09, dd (7.3, 0.7) | 7.09, d (7.3) | 7.09, d (7.2) |
| 10 | 6.05, s | 6.05, s | 6.07, br s | 6.08, s |
| 12 | 4.52, d (1.5) | 4.48, d (1.4) | 4.74, s | 4.71, s |
| 14 | 5.68, dq (1.5, 1.5) | 5.68, dq (1.4, 1.4) | 5.68, dq (1.4, 1.4) | 5.70, s |
| 15a | 2.77, d (17.8) | 2.77, d (17.8) | 2.83, d (17.8) | 2.84, d (17.8) |
| 15b | 2.68, d (17.8) | 2.67, d (17.8) | 2.60, d (17.8) | 2.61, d (17.8) |
| 21 | 6.17, s | 6.16, s | 6.11, s | 6.12, s |
| 23a | - | 3.18, d (17.0) | 3.22, d (17.3) | a. 3.21, d (17.3) |
| 23b | - | 3.02, d (17.0) | 3.10, d (17.3) | b. 3.08, d (17.3) |
| 25 | 4.59, dd (4.4, 1.8) | 4.96, dd (9.8, 6.2) | 4.31, br d (10.9) | 4.33, d (10.8) |
| 26a | 2.62, ddd (14.1, 6.7, 4.4) | 2.74, ddd (13.0, 8.9, 6.2) | 2.04, ddd (14.3, 10.9, 3.0) | 1.99, dd (13.2, 10.8) |
| 26b | 1.98, ddd (14.1, 10.2, 1.8) | 2.34, ddd (13.0, 9.8, 9.8) | 1.85, dd (14.3, 9.8) | 1.87, dd (13.2, 10.8) |
| 27 | 4.81, dd (10.2, 6.7) | 4.70, dd (9.8, 8.9) | 4.25, dd (9.8, 3.0) | 4.45, dd (10.8, 2.4) |
| 29 | 1.86, d (1.5) | 1.88, dd (1.4, 1.4) | 1.91, br s | 1.91, br s |
| 2′ | 5.75, d (13.9) | 5.74, d (15.2) | 5.76, d (15.2) | 5.75, d (15.2) |
| 3′ | 7.24, dd (13.9, 11.2) | 7.23, dd (15.2, 11.2) | 7.22, dd (15.2, 11.3) | 7.22, dd (15.2, 11.3) |
| 4′ | 6.12, dd (14.8, 11.2) | 6.12, dd (14.8, 11.2) | 6.21, dd (14.9, 11.3) | 6.21, dd (14.8, 11.3) |
| 5′ | 6.52, dd (14.8, 10.8) | 6.52, dd (14.8, 10.8) | 6.58, dd (14.9, 10.7) | 6.58, dd (14.8, 11.0) |
| 6′ | 6.12, dd (15.0, 10.8) | 6.13, dd (14.8, 10.8) | 6.16, dd (14.9, 10.7) | 6.16, dd (15.0, 11.0) |
| 7′ | 5.95, dq (15.0, 6.8) | 5.95, dq (14.8, 6.9) | 5.98, dq (14.9, 6.8) | 5.98, dq (15.0, 6.8) |
| 8′ | 1.82, d (6.8) | 1.82, d (6.9) | 1.79, d (6.8) | 1.79, d (6.8) |
| 30-OMe | 3.75, s | 3.76, s | 3.67, s | 3.66, s |
| 28-OMe | - | - | - | 3.75, s |
| 1-OH | 11.58, s | 11.65, s | ||
| 11-OH | 14.21 A, br s | 14.11, s | ||
| 20-OH | 11.05, s | 11.31, s | ||
| 28-OH | 14.06 A, br s | - |
| (1) δC, Mult (CDCl3) | (2) δC, Mult (CDCl3) | (2) δC, Mult (methanol-d4) | (3) δC, Mult (methanol-d4) | (4) δC, Mult (methanol-d4) | |
|---|---|---|---|---|---|
| 1 | 161.9, C | 161.9, C | 162.9, C | 162.8, C | 163.1, C |
| 2 | 119.5, CH | 119.5, CH | 120.1, CH | 120.0, CH | 120.1, CH |
| 3 | 136.3, CH | 136.2, CH | 137.2, CH | 137.0, CH | 137.1, CH |
| 4 | 122.2, CH | 122.2, CH | 123.1, CH | 123.0, CH | 123.1, CH |
| 5 | 137.2, C | 137.2, C | 138.7, C | 138.7, C | 138.7, C |
| 6 | 115.2, C | 115.2, C | 116.1, C | 116.2, C | 116.2, C |
| 7 | 186.7, C | 186.9, C | 188.3, C | ND | ND |
| 8 | 106.6, C | 106.4, C | 107.6, C | 107.5 B, C | 107.5 B, C |
| 9 | 41.5, C | 41.5, C | 42.7, C | 42.8, C | 42.7, C |
| 10 | 72.8, CH | 72.8, CH | 74.3, CH | 74.5, CH | 74.4, CH |
| 11 | 185.8, C | 185.8, C | 186.4, C | 187.4 B, C | 187.3 B, C |
| 12 | 43.4, CH | 43.3, CH | 44.3, CH | 44.4, CH | 44.4, CH |
| 13 | 141.4, C | 141.6, C | 142.9, C | 143.2, C | 143.0, C |
| 14 | 125.9, CH | 125.8, CH | 126.8, CH | 126.6, CH | 126.8, CH |
| 15 | 35.9, CH2 | 36.0, CH2 | 36.4, CH2 | 36.6, CH2 | 36.5, CH2 |
| 16 | 148.7, C | 149.6, C | 150.4, C | 150.0, C | 150.1, C |
| 17 | 115.3 A, C | 115.0, C | 116.2, C | 116.2 B, C | 116.1, C |
| 18 | 154.1, C | 155.2, C | 156.9, C | 157.6, C | 157.4, C |
| 19 | 105.4, C | 105.9, C | 106.8, C | 106.9, C | 106.9, C |
| 20 | 160.4, C | 160.2, C | 161.1, C | 161.0, C | 161.1, C |
| 21 | 115.2 A, CH | 114.4, CH | 114.6, CH | 114.1, CH | 114.2, CH |
| 22 | 188.0, C | 193.9, C | 196.1, C | 198.1, C | 197.9, C |
| 23 | 99.8, C | 38.2, C | 39.6, CH2 | 40.6, CH2 | 40.3, CH2 |
| 24 | 84.3, C | 84.2, C | 85.4, C | 89.2, C | 89.2, C |
| 25 | 68.9, CH | 77.7, CH | 78.8, CH | 72.8, CH | 71.7, CH |
| 26 | 32.4, CH2 | 31.5, CH2 | 32.4, CH2 | 37.7, CH2 | 36.9, CH2 |
| 27 | 64.1, CH | 67.7, CH | 68.4, CH | 69.5, CH | 68.3, CH |
| 28 | 176.8, C | 175.4, C | 177.6, C | 180.3, C | 176.6, C |
| 29 | 21.0, CH3 | 21.0, CH3 | 20.8, CH3 | 20.7, CH3 | 20.7, CH3 |
| 30 | 170.5, C | 168.8, C | 170.6, C | 171.9, C | 171.8, C |
| 1′ | 166.7, C | 166.7, C | 167.8, C | 167.8, C | 167.8, C |
| 2′ | 119.2, CH | 119.3, CH | 119.9, CH | 120.0, CH | 120.0, CH |
| 3′ | 146.5, CH | 146.4, CH | 147.6, CH | 147.6, CH | 147.6, CH |
| 4′ | 127.5, CH | 127.5, CH | 128.6, CH | 128.6, CH | 128.6, CH |
| 5′ | 142.2, CH | 142.2, CH | 143.4, CH | 143.4, CH | 143.4, CH |
| 6′ | 131.4, CH | 131.4, CH | 132.5, CH | 132.5, CH | 132.5, CH |
| 7′ | 136.1, CH | 136.0, CH | 136.7, CH | 136.6, CH | 136.7, CH |
| 8′ | 18.8, CH3 | 18.8, CH3 | 18.6, CH3 | 18.6, CH3 | 18.6, CH3 |
| 30-OMe | 54.0, CH3 | 54.2, CH3 | 54.0, CH3 | 53.4, CH3 | 53.5, CH3 |
| 28-OMe | 52.6, CH3 |
| Assay Type | (1) | (2) | (3) | (4) |
|---|---|---|---|---|
| Antibacterial–Gram-positive | ||||
| Staphylococcus aureus (ATCC 25923) | 1.8 | 6.6 | >30 | 3.7 |
| Staphylococcus aureus A (581101692:1) | 1.8 | 5.8 | >30 | 4.3 |
| Staphylococcus aureus B (AUS-RBWH-MRSA-01) | 2.4 | 6.4 | >30 | 3.7 |
| Enterococcus faecalis C (AUS-RBWH-VRE-01) | 2.6 | 3.9 | >30 | 3.7 |
| Enterococcus faecalis D (ACM 5184) | 1.3 | 1.8 | >30 | 0.5 |
| Antibacterial–Gram-negative | ||||
| Escherichia coli (ATCC 11775) | >30 | >30 | >30 | >30 |
| Antifungal | ||||
| Candida albicans (ATCC 10231) | 4.1 | >30 | >30 | >30 |
| Anticancer–cell cytotoxicity | ||||
| Human colon carcinoma (SW620) | 9.8 | >30 | >30 | >30 |
| Human lung carcinoma (NCI-H460) | 9.0 | >30 | >30 | >30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sritharan, T.; Salim, A.A.; Khalil, Z.G.; Capon, R.J. Jugiones A–D: Antibacterial Xanthone–Anthraquinone Heterodimers from Australian Soil-Derived Penicillium shearii CMB-STF067. Antibiotics 2024, 13, 97. https://doi.org/10.3390/antibiotics13010097
Sritharan T, Salim AA, Khalil ZG, Capon RJ. Jugiones A–D: Antibacterial Xanthone–Anthraquinone Heterodimers from Australian Soil-Derived Penicillium shearii CMB-STF067. Antibiotics. 2024; 13(1):97. https://doi.org/10.3390/antibiotics13010097
Chicago/Turabian StyleSritharan, Thulasi, Angela A. Salim, Zeinab G. Khalil, and Robert J. Capon. 2024. "Jugiones A–D: Antibacterial Xanthone–Anthraquinone Heterodimers from Australian Soil-Derived Penicillium shearii CMB-STF067" Antibiotics 13, no. 1: 97. https://doi.org/10.3390/antibiotics13010097