

Advances in the Discovery of Efflux Pump Inhibitors as Novel Potentiators to Control Antimicrobial-Resistant Pathogens


Abstract
:1. Introduction
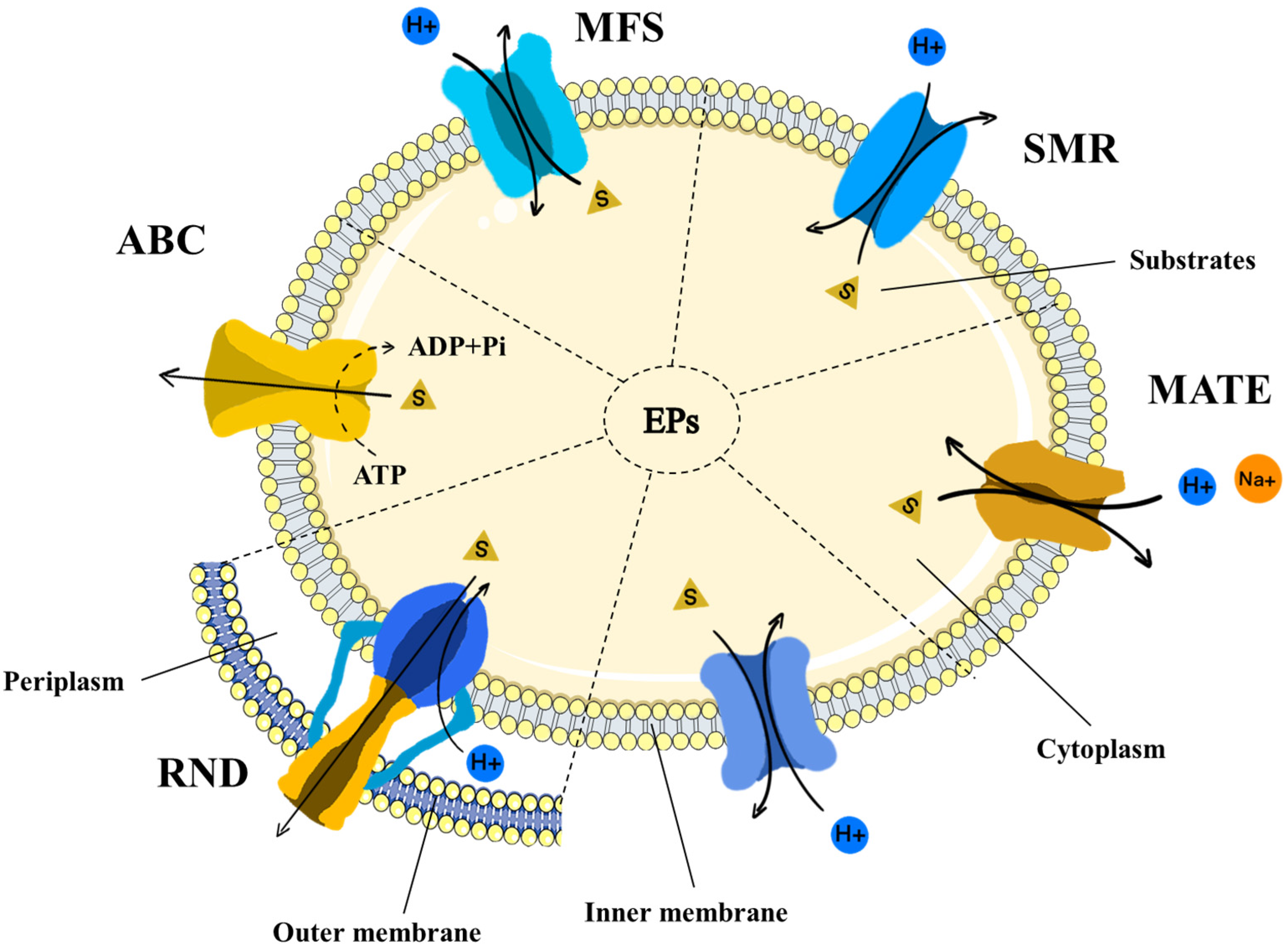
2. Classification of Efflux Pumps and Their Roles in Antibiotic Resistance
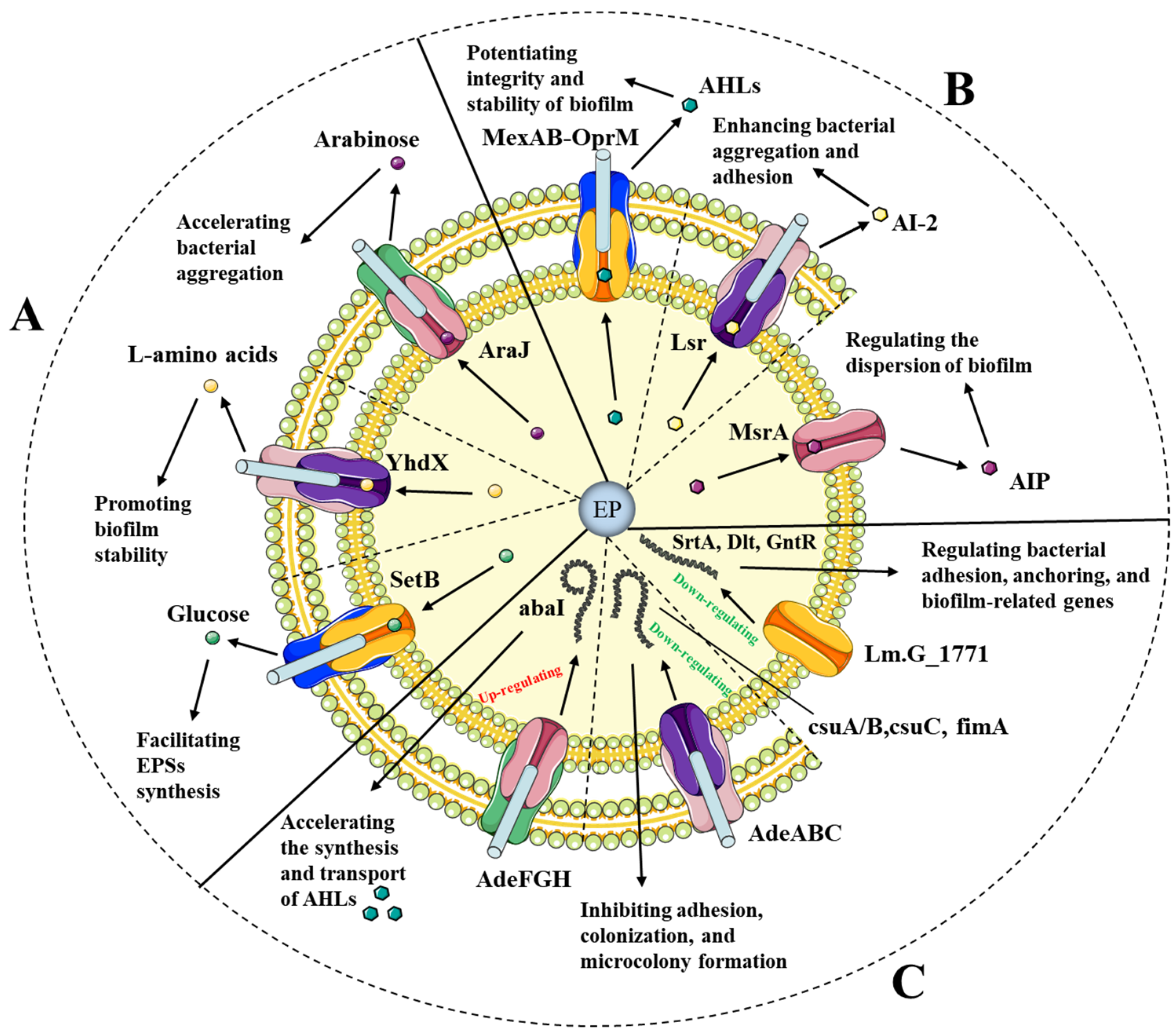
3. Efflux-Pump-Mediated Biofilm Formation
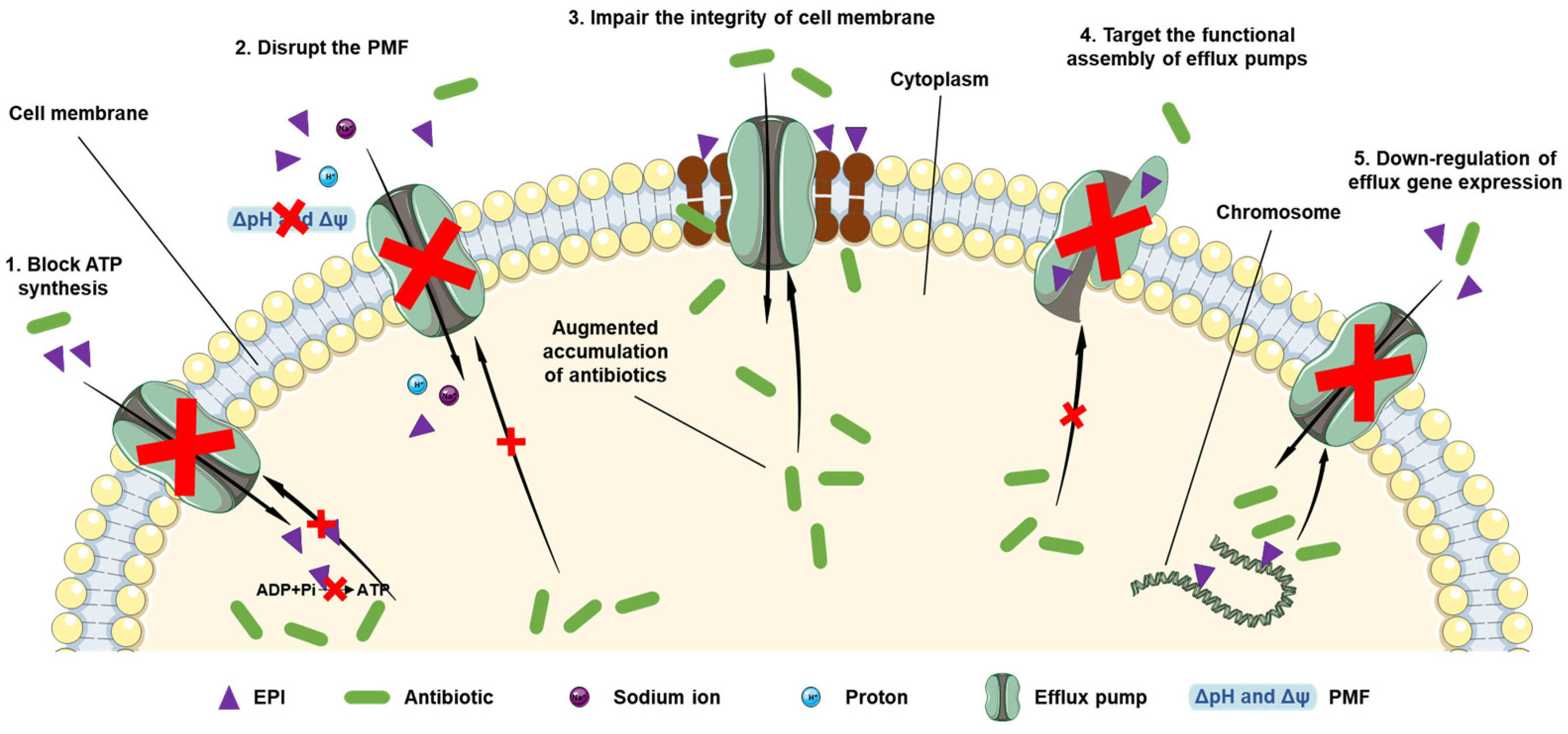
4. EPIs as a Promising Strategy to Combat Antimicrobial Resistance
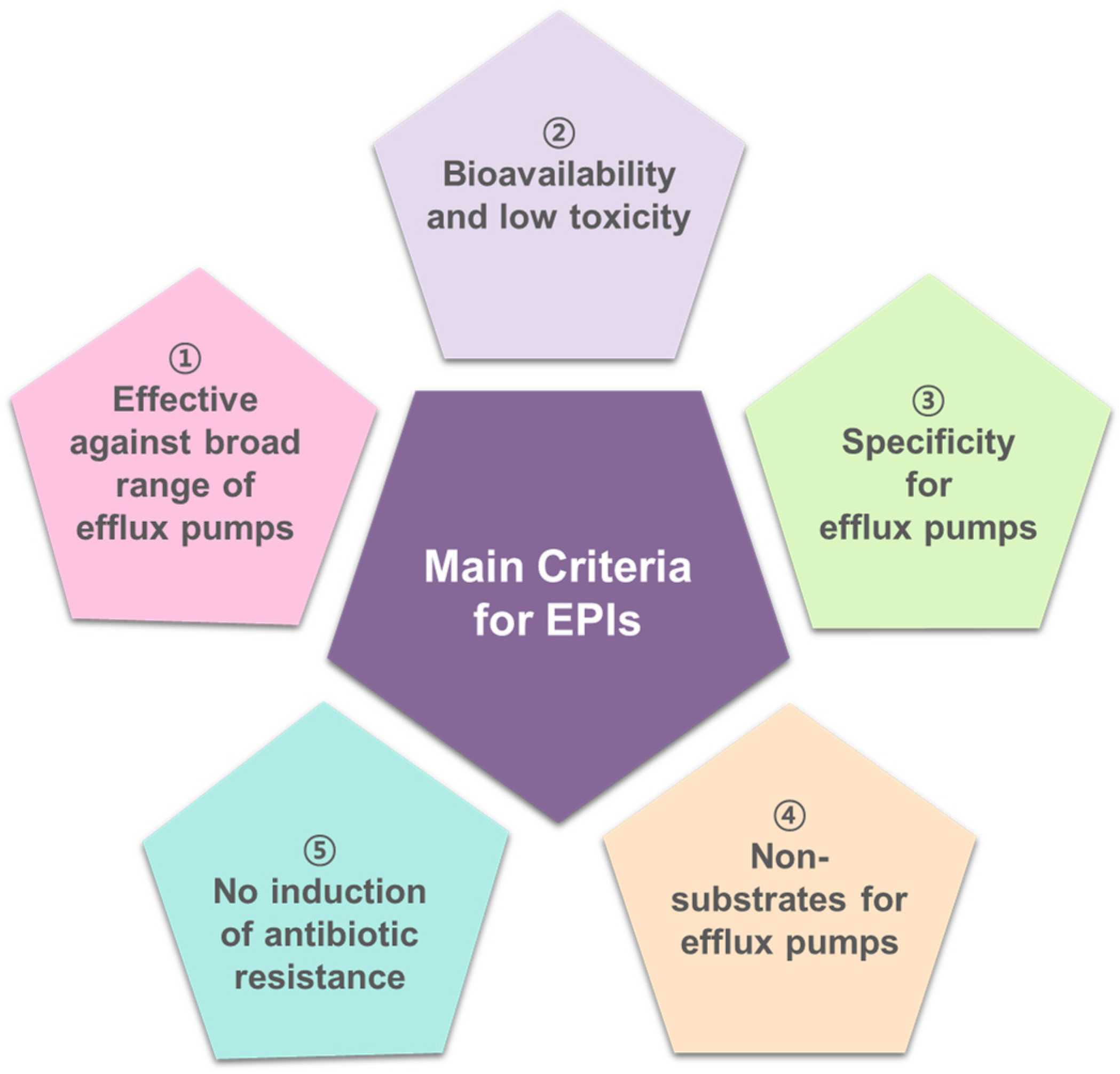
5. Sustainability Criteria for EPIs
6. Conventional and Synthetic EPIs
7. Discovery of Novel Natural EPIs
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jasovsky, D.; Littmann, J.; Zorzet, A.; Cars, O. Antimicrobial resistance—A threat to the world’s sustainable development. Ups. J. Med. Sci. 2016, 121, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Munita, J.; Arias, C. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4, 481–511. [Google Scholar] [CrossRef]
- Webber, M.A.; Piddock, L.J. The importance of efflux pumps in bacterial antibiotic resistance. J. Antimicrob. Chemother. 2003, 51, 9–11. [Google Scholar] [CrossRef]
- Piddock, L.J. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Ind. J. Med. Res. 2019, 149, 129–145. [Google Scholar]
- Kaatz, G.W.; DeMarco, C.E.; Seo, S.M. MepR, a repressor of the Staphylococcus aureus MATE family multidrug efflux pump MepA, is a substrate-responsive regulatory protein. Antimicrob. Agents Chemother. 2006, 50, 1276–1281. [Google Scholar] [CrossRef] [PubMed]
- Horiyama, T.; Yamaguchi, A.; Nishino, K. TolC dependency of multidrug efflux systems in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2010, 65, 1372–1376. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Corona, F.; Martínez, J.L. Biolog phenotype microarray is a tool for the identification of multidrug resistance efflux pump inducers. Antimicrob. Agents Chemother. 2018, 62, e01263-18. [Google Scholar] [CrossRef]
- Perez, A.; Poza, M.; Aranda, J.; Latasa, C.; Medrano, F.J.; Tomas, M.; Romero, A.; Lasa, I.; Bou, G. Effect of transcriptional activators SoxS, RobA, and RamA on expression of multidrug efflux pump AcrAB-TolC in Enterobacter cloacae. Antimicrob. Agents Chemother. 2012, 56, 6256–6266. [Google Scholar] [CrossRef] [PubMed]
- Llanes, C.; Hocquet, D.; Vogne, C.; Benali-Baitich, D.; Neuwirth, C.; Plesiat, P. Clinical strains of Pseudomonas aeruginosa overproducing MexAB-OprM and MexXY efflux pumps simultaneously. Antimicrob. Agents Chemother. 2004, 48, 1797–1802. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Beardmore, R.E.; Pena-Miller, R.; Gori, F.; Iredell, J. Antibiotic cycling and antibiotic mixing: Which one best mitigates antibiotic resistance? Mol. Biol. Evol. 2017, 34, 802–817. [Google Scholar] [CrossRef]
- Bergstrom, C.T.; Lo, M.; Lipsitch, M. Ecological theory suggests that antimicrobial cycling will not reduce antimicrobial resistance in hospitals. Proc. Natl. Acad. Sci. USA 2004, 101, 13285–13290. [Google Scholar] [CrossRef] [PubMed]
- Yeh, P.J.; Hegreness, M.J.; Aiden, A.P.; Kishony, R. Drug interactions and the evolution of antibiotic resistance. Nat. Rev. Microbiol. 2009, 7, 460–466. [Google Scholar] [CrossRef]
- AlMatar, M.; Albarri, O.; Makky, E.A.; Koksal, F. Efflux pump inhibitors: New updates. Pharmacol. Rep. 2021, 73, 1–16. [Google Scholar] [CrossRef]
- Bhattacharyya, T.; Sharma, A.; Akhter, J.; Pathania, R. The small molecule IITR08027 restores the antibacterial activity of fluoroquinolones against multidrug-resistant Acinetobacter baumannii by efflux inhibition. Int. J. Antimicrob. Agents 2017, 50, 219–226. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Warren, M.S.; Lee, A.; Galazzo, J.; Fronko, R.; Lee, M.; Blais, J.; Cho, D.; Chamberland, S.; Renau, T.; et al. Identification and characterization of inhibitors of multidrug resistance efflux pumps in Pseudomonas aeruginosa: Novel agents for combination therapy. Antimicrob. Agents Chemother. 2001, 45, 105–116. [Google Scholar] [CrossRef]
- Renau, T.; Léger, R.; Flamme, E.; Sangalang, J.; She, M.; Yen, R.; Gannon, C.; Griffith, D.; Chamberland, S.; Lomovskaya, O.; et al. Inhibitors of efflux pumps in Pseudomonas aeruginosa potentiate the activity of the fluoroquinolone antibacterial levofloxacin. J. Med. Chem. 1999, 42, 4928–4931. [Google Scholar] [CrossRef] [PubMed]
- Koncha, R.R.; Ramachandran, G.; Sepuri, N.B.V.; Ramaiah, K.V.A. CCCP-induced mitochondrial dysfunction—Characterization and analysis of integrated stress response to cellular signaling and homeostasis. FEBS J. 2021, 288, 5737–5754. [Google Scholar] [CrossRef]
- Kumar, A.; Schweizer, H.P. Bacterial resistance to antibiotics: Active efflux and reduced uptake. Adv. Drug Deliv. Rev. 2005, 57, 1486–1513. [Google Scholar] [CrossRef] [PubMed]
- Baugh, S.; Phillips, C.R.; Ekanayaka, A.S.; Piddock, L.J.; Webber, M.A. Inhibition of multidrug efflux as a strategy to prevent biofilm formation. J. Antimicrob. Chemother. 2014, 69, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.A.; Liu, Q.; Henderson, P.J.; Paulsen, I.T. Homologs of the Acinetobacter baumannii AceI transporter represent a new family of bacterial multidrug efflux systems. MBio 2015, 6, e01982-14. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F. Multiple molecular mechanisms for multidrug resistance transporters. Nature 2007, 446, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Bolla, J.R.; Howes, A.C.; Fiorentino, F.; Robinson, C.V. Assembly and regulation of the chlorhexidine-specific efflux pump AceI. Proc. Natl. Acad. Sci. USA 2020, 117, 17011–17018. [Google Scholar] [CrossRef]
- Takatsuka, Y.; Nikaido, H. Covalently linked trimer of the AcrB multidrug efflux pump provides support for the functional rotating mechanism. J. Bacteriol. 2009, 191, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- Baylay, A.J.; Piddock, L.J. Clinically relevant fluoroquinolone resistance due to constitutive overexpression of the PatAB ABC transporter in Streptococcus pneumoniae is conferred by disruption of a transcriptional attenuator. J. Antimicrob. Chemother. 2015, 70, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Alcalde-Rico, M.; Hernando-Amado, S.; Blanco, P.; Martinez, J.L. Multidrug efflux pumps at the crossroad between antibiotic resistance and bacterial virulence. Front. Microbiol. 2016, 7, 1483. [Google Scholar] [CrossRef]
- Kobayashi, N.; Nishino, K.; Yamaguchi, A. Novel macrolide-specific ABC-type efflux transporter in Escherichia coli. J. Bacteriol. 2001, 183, 5639–5644. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Villet, R.A.; Estabrooks, Z.A.; Hooper, D.C. Native efflux pumps contribute resistance to antimicrobials of skin and the ability of Staphylococcus aureus to colonize skin. J. Infect. Dis. 2014, 209, 1485–1493. [Google Scholar] [CrossRef] [PubMed]
- Lamut, A.; Peterlin Masic, L.; Kikelj, D.; Tomasic, T. Efflux pump inhibitors of clinically relevant multidrug resistant bacteria. Med. Res. Rev. 2019, 39, 2460–2504. [Google Scholar] [CrossRef]
- LaBreck, P.; Bochi-Layec, A.; Stanbro, J.; Dabbah-Krancher, G.; Simons, M.; Merrell, D. Systematic analysis of efflux pump-mediated antiseptic resistance in Staphylococcus aureus suggests a need for greater antiseptic stewardship. mSphere 2020, 5, e00959-19. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, M.; Bonaccorsi di Patti, M.C.; Fanelli, G.; Utsumi, R.; Eguchi, Y.; Trirocco, R.; Prosseda, G.; Grossi, M.; Colonna, B. Host-bacterial pathogen communication: The Wily role of the multidrug efflux pumps of the MFS family. Front. Mol. Biosci. 2021, 8, 723274. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Plesiat, P.; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Lu, J.; Chen, Z.; Nguyen, S.H.; Mao, L.; Li, J.; Yuan, Z.; Guo, J. Antidepressant fluoxetine induces multiple antibiotics resistance in Escherichia coli via ROS-mediated mutagenesis. Environ. Int. 2018, 120, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate specificities of MexAB-OprM, MexCD-OprJ, and MexXY-OprM efflux pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [PubMed]
- Sobel, M.L.; Hocquet, D.; Cao, L.; Plesiat, P.; Poole, K. Mutations in PA3574 (nalD) lead to increased MexAB-OprM expression and multidrug resistance in laboratory and clinical isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2005, 49, 1782–1786. [Google Scholar] [CrossRef] [PubMed]
- Alnaseri, H.; Kuiack, R.C.; Ferguson, K.A.; Schneider, J.E.T.; Heinrichs, D.E.; McGavin, M.J. DNA binding and sensor specificity of FarR, a novel TetR family regulator required for induction of the fatty acid efflux pump FarE in Staphylococcus aureus. J. Bacteriol. 2019, 201, e00602-18. [Google Scholar] [CrossRef]
- Marchand, I.; Damier-Piolle, L.; Courvalin, P.; Lambert, T. Expression of the RND-type efflux pump AdeABC in Acinetobacter baumannii is regulated by the AdeRS two-component system. Antimicrob. Agents Chemother. 2004, 48, 3298–3304. [Google Scholar] [CrossRef] [PubMed]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-nodulation-cell division-type efflux pump involved in aminoglycoside resistance in Acinetobacter baumannii strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef]
- Coyne, S.; Rosenfeld, N.; Lambert, T.; Courvalin, P.; Périchon, B. Overexpression of resistance-nodulation-cell division pump AdeFGH confers multidrug resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4389–4393. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, N.; Bouchier, C.; Courvalin, P.; Périchon, B. Expression of the resistance-nodulation-cell division pump AdeIJK in Acinetobacter baumannii is regulated by AdeN, a TetR-type regulator. Antimicrob. Agents Chemother. 2012, 56, 2504–2510. [Google Scholar] [CrossRef] [PubMed]
- He, G.X.; Kuroda, T.; Mima, T.; Morita, Y.; Mizushima, T.; Tsuchiya, T. An H+-coupled multidrug efflux pump, PmpM, a member of the MATE family of transporters, from Pseudomonas aeruginosa. J. Bacteriol. 2004, 186, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Schindler, B.D.; Kaatz, G.W. Multidrug efflux pumps of Gram-positive bacteria. Drug Resist. Uptat. 2016, 27, 1–13. [Google Scholar] [CrossRef]
- Bay, D.C.; Rommens, K.L.; Turner, R.J. Small multidrug resistance proteins: A multidrug transporter family that continues to grow. Biochim. Biophys. Acta 2008, 1778, 1814–1838. [Google Scholar] [CrossRef]
- Srinivasan, V.B.; Rajamohan, G. KpnEF, a new member of the Klebsiella pneumoniae cell envelope stress response regulon, is an SMR-type efflux pump involved in broad-spectrum antimicrobial resistance. Antimicrob. Agents Chemother. 2013, 57, 4449–4462. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug efflux pumps in Staphylococcus aureus: An update. Open Microbiol. J. 2013, 7, 59–71. [Google Scholar] [CrossRef]
- Buffet-Bataillon, S.; Tattevin, P.; Maillard, J.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Efflux pump induction by quaternary ammonium compounds and fluoroquinolone resistance in bacteria. Future Microbiol. 2016, 11, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Masaoka, Y.; Ueno, Y.; Morita, Y.; Kuroda, T.; Mizushima, T.; Tsuchiya, T. A two-component multidrug efflux pump, EbrAB, in Bacillus subtilis. J. Bacteriol. 2000, 182, 2307–2310. [Google Scholar] [CrossRef]
- Liu, Q.; Hassan, K.A.; Ashwood, H.E.; Gamage, H.; Li, L.; Mabbutt, B.C.; Paulsen, I.T. Regulation of the aceI multidrug efflux pump gene in Acinetobacter baumannii. J. Antimicrob. Chemother. 2018, 73, 1492–1500. [Google Scholar] [CrossRef]
- Davidson, A.L.; Dassa, E.; Orelle, C.; Chen, J. Structure, function, and evolution of bacterial ATP-binding cassette systems. Microbiol. Mol. Biol. Rev. 2008, 72, 317–364. [Google Scholar] [CrossRef]
- Chetri, S. The culmination of multidrug-resistant efflux pumps vs. meager antibiotic arsenal era: Urgent need for an improved new generation of EPIs. Front. Microbiol. 2023, 14, 1149418. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, M.; Grossi, M.; Zennaro, A.; Fanelli, G.; Micheli, G.; Barras, F.; Colonna, B.; Prosseda, G. The varied role of efflux pumps of the MFS family in the interplay of bacteria with animal and plant cells. Microorganisms 2019, 7, 285. [Google Scholar] [CrossRef] [PubMed]
- Poelarends, G.; Mazurkiewicz, P.; Konings, W. Multidrug transporters and antibiotic resistance in Lactococcus lactis. Biochim. Biophys. Acta 2002, 1555, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Huang, C.; Su, H.; Zhang, Z.; Chen, M.; Wang, R.; Zhang, D.; Zhang, L.; Liu, M. Luteolin increases susceptibility to macrolides by inhibiting MsrA efflux pump in Trueperella pyogenes. Vet. Res. 2022, 53, 3. [Google Scholar] [CrossRef] [PubMed]
- Guelfo, J.R.; Rodriguez-Rojas, A.; Matic, I.; Blazquez, J. A MATE-family efflux pump rescues the Escherichia coli 8-oxoguanine-repair-deficient mutator phenotype and protects against H2O2 killing. PLoS Genet. 2010, 6, e1000931. [Google Scholar] [CrossRef] [PubMed]
- Tocci, N.; Iannelli, F.; Bidossi, A.; Ciusa, M.L.; Decorosi, F.; Viti, C.; Pozzi, G.; Ricci, S.; Oggioni, M.R. Functional analysis of pneumococcal drug efflux pumps associates the MATE DinF transporter with quinolone susceptibility. Antimicrob. Agents Chemother. 2013, 57, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Seukep, A.J.; Kuete, V.; Nahar, L.; Sarker, S.D.; Guo, M. Plant-derived secondary metabolites as the main source of efflux pump inhibitors and methods for identification. J. Pharmaceut. Anal. 2020, 10, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Takatsuka, Y. Mechanisms of RND multidrug efflux pumps. Biochim. Biophys. Acta 2009, 1794, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Kermani, A.A.; Macdonald, C.B.; Burata, O.E.; Ben Koff, B.; Koide, A.; Denbaum, E.; Koide, S.; Stockbridge, R.B. The structural basis of promiscuity in small multidrug resistance transporters. Nat. Commun. 2020, 11, 6064. [Google Scholar] [CrossRef] [PubMed]
- Spengler, G.; Kincses, A.; Gajdacs, M.; Amaral, L. New roads leading to old destinations: Efflux pumps as targets to reverse multidrug resistance in bacteria. Molecules 2017, 22, 468. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Hellwig, N.; Djahanschiri, B.; Khera, R.; Morgner, N.; Ebersberger, I.; Wang, J.; Michel, H. Assembly and functional role of PACE transporter PA2880 from Pseudomonas aeruginosa. Microbiol. Spectr. 2022, 10, e0145321. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.A.; Jackson, S.M.; Penesyan, A.; Patching, S.G.; Tetu, S.G.; Eijkelkamp, B.A.; Brown, M.H.; Henderson, P.J.; Paulsen, I.T. Transcriptomic and biochemical analyses identify a family of chlorhexidine efflux proteins. Proc. Natl. Acad. Sci. USA 2013, 110, 20254–20259. [Google Scholar] [CrossRef] [PubMed]
- Seukep, A.J.; Mbuntcha, H.G.; Kuete, V.; Chu, Y.; Fan, E.; Guo, M.Q. What approaches to thwart bacterial efflux pumps-mediated resistance? Antibiotics 2022, 11, 1287. [Google Scholar] [CrossRef] [PubMed]
- De Gaetano, G.V.; Lentini, G.; Fama, A.; Coppolino, F.; Beninati, C. Antimicrobial resistance: Two-component regulatory systems and multidrug efflux pumps. Antibiotics 2023, 12, 965. [Google Scholar] [CrossRef] [PubMed]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed]
- Subhadra, B.; Kim, D.H.; Woo, K.; Surendran, S.; Choi, C.H. Control of biofilm formation in healthcare: Recent advances exploiting quorum-sensing interference strategies and multidrug efflux pump inhibitors. Materials 2018, 11, 1676. [Google Scholar] [CrossRef]
- Reza, A.; Sutton, J.M.; Rahman, K.M. Effectiveness of efflux pump inhibitors as biofilm disruptors and resistance breakers in Gram-negative (ESKAPEE) bacteria. Antibiotics 2019, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K.; Furukawa, S.; Ogihara, H.; Morinaga, Y. Roles of multidrug efflux pumps on the biofilm formation of Escherichia coli K-12. Biocontrol Sci. 2011, 16, 69–72. [Google Scholar] [CrossRef]
- He, X.; Ahn, J. Differential gene expression in planktonic and biofilm cells of multiple antibiotic-resistant Salmonella Typhimurium and Staphylococcus aureus. FEMS Microbiol. Lett. 2011, 325, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Sionov, R.V.; Steinberg, D. Targeting the holy triangle of quorum sensing, biofilm formation, and antibiotic resistance in pathogenic bacteria. Microorganisms 2022, 10, 1239. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Miller, P.F.; Willard, J.; Olson, E.R. Functional and biochemical characterization of Escherichia coli sugar efflux transporters. J. Biol. Chem. 1999, 274, 22977–22984. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.P.; Delden, C.V.; Iglewski, B.H. Active efflux and diffusion are involved in transport of Pseudomonas aeruginosa cell-to-cell signals. J. Bacteriol. 1999, 4, 1203–1210. [Google Scholar] [CrossRef]
- Wang, J.; Jiao, H.; Meng, J.; Qiao, M.; Du, H.; He, M.; Ming, K.; Liu, J.; Wang, D.; Wu, Y. Baicalin inhibits biofilm formation and the quorum-sensing system by regulating the MsrA drug efflux pump in Staphylococcus saprophyticus. Front. Microbiol. 2019, 10, 2800. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, M.G.; Deziel, E. MexEF-OprN efflux pump exports the Pseudomonas quinolone signal (PQS) precursor HHQ (4-hydroxy-2-heptylquinoline). PLoS ONE 2011, 6, e24310. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, W.; Lametsch, R.; Aarestrup, F.; Shi, C.; She, Q.; Shi, X.; Knøchel, S. Phenotypic, proteomic, and genomic characterization of a putative ABC-transporter permease involved in Listeria monocytogenes biofilm formation. Foodborne Pathog. Dis. 2011, 8, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Chabane, Y.N.; Goussard, S.; Snesrud, E.; Courvalin, P.; De, E.; Grillot-Courvalin, C. Contribution of resistance-nodulation-cell division efflux systems to antibiotic resistance and biofilm formation in Acinetobacter baumannii. MBio 2015, 6, 10–128. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Lu, F.; Yuan, F.; Jiang, D.; Zhao, P.; Zhu, J.; Cheng, H.; Cao, J.; Lu, G. Biofilm formation caused by clinical Acinetobacter baumannii isolates is associated with overexpression of the AdeFGH efflux pump. Antimicrob. Agents Chemother. 2015, 59, 4817–4825. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Muir, T.W. Regulation of virulence in Staphylococcus aureus: Molecular mechanisms and remaining puzzles. Cell Chem. Biol. 2016, 23, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bian, Z.; Wang, Y. Biofilm formation and inhibition mediated by bacterial quorum sensing. Appl. Microbiol. Biotechnol. 2022, 106, 6365–6381. [Google Scholar] [CrossRef]
- Li, W.; Xue, M.; Yu, L.; Qi, K.; Ni, J.; Chen, X.; Deng, R.; Shang, F.; Xue, T. QseBC is involved in the biofilm formation and antibiotic resistance in Escherichia coli isolated from bovine mastitis. PeerJ 2020, 8, e8833. [Google Scholar] [CrossRef] [PubMed]
- Jani, S.; Seely, A.L.; Peabody, V.G.; Jayaraman, A.; Manson, M.D. Chemotaxis to self-generated AI-2 promotes biofilm formation in Escherichia coli. Microbiology 2017, 163, 1778–1790. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, H.; Pourhajibagher, M.; Alikhani, M.Y.; Bahador, A. The effect of antimicrobial photodynamic therapy on the expression of biofilm associated genes in Staphylococcus aureus strains isolated from wound infections in burn patients. Photodiagnosis Photodyn. Ther. 2019, 25, 406–413. [Google Scholar] [CrossRef]
- Mirzaie, A.; Ranjbar, R. Antibiotic resistance, virulence-associated genes analysis and molecular typing of Klebsiella pneumoniae strains recovered from clinical samples. AMB Express 2021, 11, 122. [Google Scholar] [CrossRef] [PubMed]
- Hajiagha, M.N.; Kafil, H.S. Efflux pumps and microbial biofilm formation. Infect. Genet. Evol. 2023, 112, 105459. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, H.; Jamshidi, S.; Sutton, J.; Rahman, K. Current advances in developing inhibitors of bacterial multidrug. Curr. Med. Chem. 2016, 23, 1062–1081. [Google Scholar] [CrossRef]
- Vargiu, A.V.; Ruggerone, P.; Opperman, T.J.; Nguyen, S.T.; Nikaido, H. Molecular mechanism of MBX2319 inhibition of Escherichia coli AcrB multidrug efflux pump and comparison with other inhibitors. Antimicrob. Agents Chemother. 2014, 58, 6224–6234. [Google Scholar] [CrossRef] [PubMed]
- Kvist, M.; Hancock, V.; Klemm, P. Inactivation of efflux pumps abolishes bacterial biofilm formation. Appl. Environ. Microbiol. 2008, 74, 7376–7382. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.A.; Mirza, Z.M.; Kumar, A.; Verma, V.; Qazi, G.N. Piperine, a phytochemical potentiator of ciprofloxacin against Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Doleans-Jordheim, A.; Veron, J.B.; Fendrich, O.; Bergeron, E.; Montagut-Romans, A.; Wong, Y.S.; Furdui, B.; Freney, J.; Dumontet, C.; Boumendjel, A. 3-Aryl-4-methyl-2-quinolones targeting multiresistant Staphylococcus aureus bacteria. ChemMedChem 2013, 8, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.C.; Ip, M.; Lau, C.B.; Lui, S.L.; Jolivalt, C.; Ganem-Elbaz, C.; Litaudon, M.; Reiner, N.E.; Gong, H.; See, R.H.; et al. Synergistic effects of baicalein with ciprofloxacin against NorA over-expressed methicillin-resistant Staphylococcus aureus (MRSA) and inhibition of MRSA pyruvate kinase. J. Ethnopharmacol. 2011, 137, 767–773. [Google Scholar] [CrossRef]
- Kaatz, G.W.; Moudgal, V.V.; Seo, S.M.; Kristiansen, J.E. Phenothiazines and thioxanthenes inhibit multidrug efflux pump activity in Staphylococcus aureus. Antimicrob. Agents Chemother. 2003, 47, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.K.; Mohanty, P. Bacterial efflux pumps involved in multidrug resistance and their inhibitors: Rejuvinating the antimicrobial chemotherapy. Recent Pat. Antiinfect. Drug Discov. 2012, 7, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Abdali, N.; Parks, J.; Haynes, K.; Chaney, J.; Green, A.; Wolloscheck, D.; Walker, J.; Rybenkov, V.; Baudry, J.; Smith, J.; et al. Reviving antibiotics efflux pump inhibitors that interact with AcrA, a membrane fusion protein of the AcrAB-TolC multidrug efflux pump. ACS Infect. Dis. 2017, 3, 89–98. [Google Scholar] [CrossRef]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pages, J.M.; Amaral, L.; Bolla, J.M. Geraniol restores antibiotic activities against multidrug-resistant isolates from gram-negative species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [PubMed]
- Guefack, M.F.; Messina, N.D.M.; Mbaveng, A.T.; Nayim, P.; Kuete, J.R.N.; Matieta, V.Y.; Chi, G.F.; Ngadjui, B.T.; Kuete, V. Antibacterial and antibiotic-potentiation activities of the hydro-ethanolic extract and protoberberine alkaloids from the stem bark of Enantia chlorantha against multidrug-resistant bacteria expressing active efflux pumps. J. Ethnopharmacol. 2022, 296, 115518. [Google Scholar] [CrossRef]
- Laws, M.; Shaaban, A.; Rahman, K.M. Antibiotic resistance breakers: Current approaches and future directions. FEMS Microbiol. Rev. 2019, 43, 490–516. [Google Scholar] [CrossRef] [PubMed]
- Borselli, D.; Lieutaud, A.; Thefenne, H.; Garnotel, E.; Pages, J.M.; Brunel, J.M.; Bolla, J.M. Polyamino-isoprenic derivatives block intrinsic resistance of P. aeruginosa to doxycycline and chloramphenicol in vitro. PLoS ONE 2016, 11, e0154490. [Google Scholar]
- Lamers, R.P.; Cavallari, J.F.; Burrows, L.L. The efflux inhibitor phenylalanine-arginine beta-naphthylamide (PAβN) permeabilizes the outer membrane of gram-negative bacteria. PLoS ONE 2013, 8, e60666. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Bostian, K.A. Practical applications and feasibility of efflux pump inhibitors in the clinic—A vision for applied use. Biochem. Pharmacol. 2006, 71, 910–918. [Google Scholar] [CrossRef]
- Opperman, T.J.; Nguyen, S.T. Recent advances toward a molecular mechanism of efflux pump inhibition. Front. Microbiol. 2015, 6, 421. [Google Scholar] [CrossRef]
- Compagne, N.; Vieira Da Cruz, A.; Muller, R.T.; Hartkoorn, R.C.; Flipo, M.; Pos, K.M. Update on the discovery of efflux pump inhibitors against critical priority Gram-negative bacteria. Antibiotics 2023, 12, 180. [Google Scholar] [CrossRef] [PubMed]
- Kaynak Onurdag, F.; Kayis, U.; Okten, S. Effect of phenylalanine-arginine-beta-naphthylamide to ciprofloxacin minimum inhibitory concentration values and expression of efflux pump system genes in Acinetobacter baumannii Isolates. Mikrobiol. Bul. 2021, 55, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Strahl, H.; Hamoen, L.W. Membrane potential is important for bacterial cell division. Proc. Natl. Acad. Sci. USA 2010, 107, 12281–12286. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Carbonel, A.; Mondragon, B.; Lopez-Chegne, N.; Pena-Tuesta, I.; Huayan-Davila, G.; Blitchtein, D.; Carrillo-Ng, H.; Silva-Caso, W.; Aguilar-Luis, M.A.; Del Valle-Mendoza, J. The effect of the efflux pump inhibitor carbonyl cyanide m-chlorophenylhydrazone (CCCP) on the susceptibility to imipenem and cefepime in clinical strains of Acinetobacter baumannii. PLoS ONE 2021, 16, e0259915. [Google Scholar] [CrossRef]
- Osei Sekyere, J.; Amoako, D.G. Carbonyl cyanide m-chlorophenylhydrazine (CCCP) reverses resistance to colistin, but not to carbapenems and tigecycline in multidrug-resistant Enterobacteriaceae. Front. Microbiol. 2017, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, A.; Steinke, P.; Bohnert, J.A.; Akova, M.; Jonas, D.; Kern, W.V. Effect of 1-(1-naphthylmethyl)-piperazine, a novel putative efflux pump inhibitor, on antimicrobial drug susceptibility in clinical isolates of Enterobacteriaceae other than Escherichia coli. J. Antimicrob. Chemother. 2006, 57, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Opperman, T.J.; Kwasny, S.M.; Kim, H.S.; Nguyen, S.T.; Houseweart, C.; D’Souza, S.; Walker, G.C.; Peet, N.P.; Nikaido, H.; Bowlin, T.L. Characterization of a novel pyranopyridine inhibitor of the AcrAB efflux pump of Escherichia coli. Antimicrob. Agents Chemother. 2014, 58, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xie, K.; Zou, D.; Meng, M.; Xie, M. Inhibitory effects of silybin on the efflux pump of methicillin-resistant Staphylococcus aureus. Mol. Med. Rep. 2018, 18, 827–833. [Google Scholar] [PubMed]
- Singh, S.; Kalia, N.P.; Joshi, P.; Kumar, A.; Sharma, P.R.; Kumar, A.; Bharate, S.B.; Khan, I.A. Boeravinone B, A novel dual inhibitor of NorA bacterial efflux pump of Staphylococcus aureus and human P-glycoprotein, reduces the biofilm formation and intracellular invasion of bacteria. Front. Microbiol. 2017, 8, 1868. [Google Scholar] [CrossRef] [PubMed]
- Cirino, I.C.; Menezes-Silva, S.M.; Silva, H.T.; de Souza, E.L.; Siqueira-Junior, J.P. The essential oil from Origanum vulgare L. and its individual constituents carvacrol and thymol enhance the effect of tetracycline against Staphylococcus aureus. Chemotherapy 2014, 60, 290–293. [Google Scholar] [CrossRef] [PubMed]
- de Morais Oliveira-Tintino, C.D.; Tintino, S.R.; Limaverde, P.W.; Figueredo, F.G.; Campina, F.F.; da Cunha, F.A.B.; da Costa, R.H.S.; Pereira, P.S.; Lima, L.F.; de Matos, Y.; et al. Inhibition of the essential oil from Chenopodium ambrosioides L. and alpha-terpinene on the NorA efflux-pump of Staphylococcus aureus. Food Chem. 2018, 262, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Alakomi, H.-L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; Wright, A.v. Characterization of the action of selected essential oil components on Gram-negative bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Lambert, R.; Skandamis, P.N.; Coote, P.; Nychas, G. A study of the minimum inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. J. Appl. Microbiol. 2001, 91, 453–462. [Google Scholar] [CrossRef]
- Miladi, H.; Zmantar, T.; Chaabouni, Y.; Fedhila, K.; Bakhrouf, A.; Mahdouani, K.; Chaieb, K. Antibacterial and efflux pump inhibitors of thymol and carvacrol against food-borne pathogens. Microb. Pathogen. 2016, 99, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Chovanová, R.; Mezovská, J.; Vaverková, Š.; Mikulášová, M. The inhibition the Tet(K) efflux pump of tetracycline resistant Staphylococcus epidermidis by essential oils from three Salvia species. Lett. Appl. Microbiol. 2015, 61, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Ghazal, T.S.A.; Schelz, Z.; Vidács, L.; Szemerédi, N.; Veres, K.; Spengler, G.; Hohmann, J. Antimicrobial, multidrug resistance reversal and biofilm formation inhibitory effect of Origanum majorana extracts, essential oil and monoterpenes. Plants 2022, 11, 1432. [Google Scholar] [CrossRef]
- Mouwakeh, A.; Kincses, A.; Nové, M.; Mosolygó, T.; Mohácsi-Farkas, C.; Kiskó, G.; Spengler, G. Nigella sativa essential oil and its bioactive compounds as resistance modifiers against Staphylococcus aureus. Phytother. Res. 2019, 33, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, A.; Mohammadzadeh, A.; Salehi, T.Z.; Mahmoodi, P.; Nourian, A. Cuminum cyminum L. essential oil: A promising antibacterial and antivirulence agent against multidrug-resistant Staphylococcus aureus. Front. Microbiol. 2021, 12, 667833. [Google Scholar] [CrossRef] [PubMed]
- Neyfakh, A.A.; Bidnenko, V.E.; Chen, L.B. Efflux-mediated multidrug resistance in Bacillus subtilis: Similarities and dissimilarities with the mammalian system. Proc. Natl. Acad. Sci. 1991, 88, 4781–4785. [Google Scholar] [CrossRef]
- Klyachko, K.A.; Schuldiner, S.; Neyfakh, A.A. Mutations affecting substrate specificity of the Bacillus subtilis multidrug transporter Bmr. J. Bacteriol. 1997, 179, 2189–2193. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, S.; Oluwatuyi, M.; Kaatz, G.W. A novel inhibitor of multidrug efflux pumps in Staphylococcus aureus. J. Antimicrob. Chemother. 2003, 51, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Wang, J. Berberine inhibits the MexXY-OprM efflux pump to reverse imipenem resistance in a clinical carbapenem-resistant Pseudomonas aeruginosa isolate in a planktonic state. Exp. Ther. Med. 2018, 15, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, Y.; Wang, X.; Guo, J.; Wang, H.; Zhang, H.; Du, F. Jatrorrhizine suppresses the antimicrobial resistance of methicillin-resistant Staphylococcus aureus. Exp. Ther. Med. 2019, 18, 3715–3722. [Google Scholar] [CrossRef] [PubMed]
- Kalia, N.P.; Mahajan, P.; Mehra, R.; Nargotra, A.; Sharma, J.P.; Koul, S.; Khan, I.A. Capsaicin, a novel inhibitor of the NorA efflux pump, reduces the intracellular invasion of Staphylococcus aureus. J. Antimicrob. Chemother. 2012, 67, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- Siriyong, T.; Srimanote, P.; Chusri, S.; Yingyongnarongkul, B.E.; Suaisom, C.; Tipmanee, V.; Voravuthikunchai, S.P. Conessine as a novel inhibitor of multidrug efflux pump systems in Pseudomonas aeruginosa. BMC Complement Altern. Med. 2017, 17, 405. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, G.R.; Tyagi, R.; Sanchita; Tripathi, S.; Pati, S.; Srivastava, S.K.; Darokar, M.P.; Sharma, A. Antibiotics potentiating potential of catharanthine against superbug Pseudomonas aeruginosa. J. Biomol. Struct. Dyn. 2018, 36, 4270–4284. [Google Scholar] [CrossRef] [PubMed]
- Yarlagadda, V.; Medina, R.; Wright, G.D. Venturicidin A, a membrane-active natural product inhibitor of ATP synthase potentiates aminoglycoside antibiotics. Sci. Rep. 2020, 10, 8134. [Google Scholar] [CrossRef]
- Dhanda, G.; Acharya, Y.; Haldar, J. Antibiotic adjuvants: A versatile approach to combat antibiotic resistance. ACS Omega 2023, 8, 10757–10783. [Google Scholar] [CrossRef]
- Lee, M.; Galazzo, J.; Staley, A.; Lee, J.; Warren, M.; Fuernkranz, H.; Chamberland, S.; Lomovskaya, O.; Miller, G. Microbial fermentation-derived inhibitors of efflux-pump-mediated drug resistance. Farmaco 2001, 56, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Tambat, R.; Jangra, M.; Mahey, N.; Chandal, N.; Kaur, M.; Chaudhary, S.; Verma, D.K.; Thakur, K.G.; Raje, M.; Jachak, S.; et al. Microbe-derived indole metabolite demonstrates potent multidrug efflux pump inhibition in Staphylococcus aureus. Front. Microbiol. 2019, 10, 2153. [Google Scholar] [CrossRef] [PubMed]
- Tambat, R.; Mahey, N.; Chandal, N.; Verma, D.K.; Jangra, M.; Thakur, K.G.; Nandanwar, H. A Microbe-Derived Efflux Pump Inhibitor of the Resistance-Nodulation-Cell Division Protein Restores Antibiotic Susceptibility in Escherichia coli and Pseudomonas aeruginosa. ACS Infect. Dis. 2022, 8, 255–270. [Google Scholar] [CrossRef]
- Whalen, K.E.; Poulson-Ellestad, K.L.; Deering, R.W.; Rowley, D.C.; Mincer, T.J. Enhancement of antibiotic activity against multidrug-resistant bacteria by the efflux pump inhibitor 3,4-dibromopyrrole-2,5-dione isolated from a Pseudoalteromonas sp. J. Nat. Prod. 2015, 78, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.T.; Kwasny, S.M.; Ding, X.; Cardinale, S.C.; McCarthy, C.T.; Kim, H.S.; Nikaido, H.; Peet, N.P.; Williams, J.D.; Bowlin, T.L.; et al. Structure-activity relationships of a novel pyranopyridine series of Gram-negative bacterial efflux pump inhibitors. Bioorg. Med. Chem. 2015, 23, 2024–2034. [Google Scholar] [CrossRef]
- Abreu, A.C.; Serra, S.C.; Borges, A.; Saavedra, M.J.; McBain, A.J.; Salgado, A.J.; Simoes, M. Combinatorial activity of flavonoids with antibiotics against drug-resistant Staphylococcus aureus. Microb. Drug Resist. 2015, 21, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Debattista, N.; Pappano, N. Antimicrobial activity and synergism of some substituted flavonoids. Folia Microbiol. 2008, 53, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Lechner, D.; Gibbons, S.; Bucar, F. Plant phenolic compounds as ethidium bromide efflux inhibitors in Mycobacterium smegmatis. J. Antimicrob. Chemother. 2008, 62, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Zloh, M.; Gibbons, S. Molecular similarity of MDR Inhibitors. Int. J. Mol. Sci. 2004, 5, 37–47. [Google Scholar] [CrossRef]
- Maia, G.L.; Falcao-Silva Vdos, S.; Aquino, P.G.; de Araujo-Junior, J.X.; Tavares, J.F.; da Silva, M.S.; Rodrigues, L.C.; de Siqueira-Junior, J.P.; Barbosa-Filho, J.M. Flavonoids from Praxelis clematidea R.M. King and Robinson modulate bacterial drug resistance. Molecules 2011, 16, 4828–4835. [Google Scholar] [CrossRef] [PubMed]
- Holler, J.G.; Christensen, S.B.; Slotved, H.C.; Rasmussen, H.B.; Guzman, A.; Olsen, C.E.; Petersen, B.; Molgaard, P. Novel inhibitory activity of the Staphylococcus aureus NorA efflux pump by a kaempferol rhamnoside isolated from Persea lingue Nees. J. Antimicrob. Chemother. 2012, 67, 1138–1144. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, R.K.; Dwivedi, P.; Ahmad, S. Screening of antibacterial activity of six plant essential oils against pathogenic bacterial strains. Asian J. Med. Sci. 2010, 2, 152–158. [Google Scholar]
- Mittal, R.P.; Rana, A.; Jaitak, V. Essential oils: An impending substitute of synthetic antimicrobial agents to overcome antimicrobial resistance. Curr. Drug Targets 2019, 20, 605–624. [Google Scholar] [CrossRef] [PubMed]
- Galvao, L.C.; Furletti, V.F.; Bersan, S.M.; da Cunha, M.G.; Ruiz, A.L.; de Carvalho, J.E.; Sartoratto, A.; Rehder, V.L.; Figueira, G.M.; Teixeira Duarte, M.C.; et al. Antimicrobial activity of essential oils against Streptococcus mutans and their antiproliferative effects. Evid. -Based Complement. Altern. Med. 2012, 2012, 751435. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.; Fernandes, J.A. Biological properties of medicinal plants: A review of their antimicrobial activity. J. Venom. Anim. Toxins Incl. Trop. Dis. 2010, 16, 402–413. [Google Scholar] [CrossRef]
- Lopez-Romero, J.C.; Gonzalez-Rios, H.; Borges, A.; Simoes, M. Antibacterial effects and mode of action of selected essential oils components against Escherichia coli and Staphylococcus aureus. Evid. -Based Complement. Altern. Med. 2015, 2015, 795435. [Google Scholar] [CrossRef]
- Oussalah, M.; Caillet, S.; Saucier, L.; Lacroix, M. Antimicrobial effects of selected plant essential oils on the growth of a Pseudomonas putida strain isolated from meat. Meat Sci. 2006, 73, 236–244. [Google Scholar] [CrossRef]
- Caillet, S.; Lacroix, M. Effect of gamma radiation and oregano essential oil on murein and ATP concentration of Listeria monocytogenes. J. Food Prot. 2006, 69, 2961–2969. [Google Scholar] [CrossRef]
- Damjanović-Vratnica, B. Herbal extracts—Possibility of preventing food-borne infection. In Significance, Prevention and Control of Food Related Diseases; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; de Martino, L.; Coppola, R.; de Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Lammari, N.; Louaer, O.; Meniai, A.H.; Elaissari, A. Encapsulation of essential oils via nanoprecipitation process: Overview, progress, challenges and prospects. Pharmaceutics 2020, 12, 431. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, Y.; Feng, J.; Wang, H.; Yang, Y.; Ai, Q.; Zhang, Z.; Chu, S.; Chen, N. CZK, a novel alkaloid derivative from Clausena lansium, alleviates ischemic stroke injury through Nrf2-mediated antioxidant effects. Sci. Rep. 2023, 13, 6053. [Google Scholar] [CrossRef]
- Purwaningsih, I.; Maksum, I.P.; Sumiarsa, D.; Sriwidodo, S. A review of Fibraurea tinctoria and its component, berberine, as an antidiabetic and antioxidant. Molecules 2023, 28, 1294. [Google Scholar] [CrossRef]
- He, X.; Jin, Y.; Kong, F.; Yang, L.; Zhu, M.; Wang, Y. Discovery, antitumor activity, and fermentation optimization of roquefortines from Penicillium sp. OUCMDZ-1435. Molecules 2023, 28, 3180. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, Y.; Vong, C.T.; Zhu, Y.; Xu, B.; Ruan, C.C.; Wang, Y.; Cheang, W.S. Jatrorrhizine improves endothelial function in diabetes and obesity through suppression of endoplasmic reticulum stress. Int. J. Mol. Sci. 2022, 23, 12064. [Google Scholar] [CrossRef]
- Ning, N.; He, K.; Wang, Y.; Zou, Z.; Wu, H.; Li, X.; Ye, X. Hypolipidemic effect and mechanism of palmatine from coptis chinensis in hamsters fed high-fat diet. Phytother. Res. 2015, 29, 668–673. [Google Scholar] [CrossRef]
- Avci, F.G.; Atas, B.; Aksoy, C.S.; Kurpejovic, E.; Gulsoy Toplan, G.; Gurer, C.; Guillerminet, M.; Orelle, C.; Jault, J.M.; Sariyar Akbulut, B. Repurposing bioactive aporphine alkaloids as efflux pump inhibitors. Fitoterapia 2019, 139, 104371. [Google Scholar] [CrossRef] [PubMed]
- Chagnon, F.; Guay, I.; Bonin, M.A.; Mitchell, G.; Bouarab, K.; Malouin, F.; Marsault, E. Unraveling the structure-activity relationship of tomatidine, a steroid alkaloid with unique antibiotic properties against persistent forms of Staphylococcus aureus. Eur. J. Med. Chem. 2014, 80, 605–620. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, X.Y.; Li, M.K.; Hou, Z.; Zhou, Y.; Chen, Z.; Meng, J.R.; Luo, X.X.; Tang, H.F.; Xue, X.Y. A novel pregnane-type alkaloid from Pachysandra terminalis inhibits methicillin-resistant Staphylococcus aureus in vitro and in vivo. Phytother. Res. 2015, 29, 373–380. [Google Scholar] [CrossRef]
- Veale, C.G.; Zoraghi, R.; Young, R.M.; Morrison, J.P.; Pretheeban, M.; Lobb, K.A.; Reiner, N.E.; Andersen, R.J.; Davies-Coleman, M.T. Synthetic analogues of the marine bisindole deoxytopsentin: Potent selective inhibitors of MRSA pyruvate kinase. J. Nat. Prod. 2015, 78, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Hanawa, F.; Fokialakis, N.; Skaltsounis, A.L. Photo-activated DNA binding and antimicrobial activities of furoquinoline and pyranoquinolone alkaloids from rutaceae. Planta Med. 2004, 70, 531–535. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Efflux Pump Family | Efflux Pump Regulator | Strain | Substrate | Reference |
|---|---|---|---|---|
| ABC (PATA/B) | Streptococcus pneumoniae | Ciprofloxacin, levofloxacin, and norfloxacin (hydrophilic fluoroquinolones) | [26] | |
| ABC (MacAB-TolC) | BaeSR (−) | Escherichia coli | Lipopolysaccharides, polypeptide virulence factors, and macrolides | [27,28] |
| MFS (Tet38) | TetR21, MgrA (−) | Staphylococcus aureus | Glycerol-3-phosphate, fosfomycin, tetracycline, and certain unsaturated fatty acids | [29] |
| MFS (NorA) | NorR (+) MgrA (−) | Staphylococcus aureus | Fluoroquinolones, reserpine, dyes, pentamidine, phenothiazines, and omeprazole | [30] |
| MFS (QacA) | QacR (−) | Staphylococcus aureus | Bisbiguanides, quaternary ammonium compounds (QACs), diamides, and aromatic dyes | [31] |
| MFS (KpnGH) | Klebsiella pneumoniae | Detergents, cationic dyes, bile salts, and antiseptic chemicals | [32] | |
| RND (AcrAB-TolC) | RamA, AcrR (+) MarR, SoxR (−) | Escherichia coli | Tetracycline, levofloxacin, chloramphenicol, norfloxacin, bile salts, organic solvents, fatty acids, and dyes | [33,34] |
| RND (MexAB-oprM) | BrlR, CpxR (+) mexR, nalD (−) | Pseudomonas aeruginosa | β-lactams, chloramphenicol, fluoroquinolones, macrolides, novobiocin, tetracycline, trimethoprim, detergents, organic solvents, and dyes | [35,36] |
| RND (FarE) | farR (−) | Staphylococcus aureus | Linoleic acid, fatty acid, and rhodomyrtone | [37] |
| RND (AdeABC) | AdeRS (−) | Acinetobacter baumannii | Aminoglycosides, β-lactams, chloramphenicol, erythromycin, tetracyclines, and EtBr | [38,39] |
| RND (AdeFGH) | AdeL (−) | Acinetobacter baumannii | Clindamycin, fluoroquinolones, and tigecycline | [40] |
| RND (AdeIJK) | AdeN (−) | Acinetobacter baumannii | β-lactam, fluoroquinolones, tetracyclines, tigecycline, lincosamides, rifampin, chloramphenicol, co-trimoxazole, novobiocin, and fusidic acid | [41] |
| MATE (PmpM) | Pseudomonas aeruginosa | Tetraphenylphosphonium chloride, acriflavine, EtBr, benzalkonium chloride, and fluoroquinolones | [42] | |
| MATE (MepA) | mepR (−) | Staphylococcus aureus | Tigecycline, hydrophilic fluoroquinolones, dyes, and fungicides | [43] |
| SMR (EmrE) | Escherichia coli | Benzalkonium, EtBr, tetraphenylphosphonium, methyl viologen, betaine, and choline | [44] | |
| SMR (KpnEF) | CpxR (+) | Klebsiella pneumoniae | Erythromycin, ceftriaxone, tetracycline, cefepime, rifampin, SDS, EtBr, chlorhexidine, benzalkonium chloride, triclosan, and acriflavine | [45] |
| SMR (QacC) | Staphylococcus aureus | Quaternary ammonium compound, chlorhexidine, and EtBr | [46,47] | |
| SMR (EbrAB) | Bacillus subtilis | Cationic lipophilic dyes, including safranin O, pyronine Y, EtBr, and acriflavine | [48] | |
| PACE (AceI) | AceR (+) | Acinetobacter baumannii | Proflavine, chlorhexidine, acriflavine, dequalinium, and benzalkonium | [49] |
| Biofilm Factor | Strain | Efflux Pump | Target Component | Function | Reference |
|---|---|---|---|---|---|
| EPS matrix | Escherichia coli | MFS (SetB) | Glucose | EPS matrix synthesis | [71] |
| E. coli | ABC (YhdX) | L-amino acids | Biofilm stability | [72] | |
| E. coli | MFS (AraJ) | Arabinose | Bacterial aggregation | [65] | |
| QS signals | Pseudomonas aeruginosa | RND (MexAB-OprM) | N-3-oxododecanoyl-l-homoserine lactone | Biofilm formation | [73] |
| Staphylococcus aureus | ABC (MsrA) | agrA and sarA | Biofilm formation | [74] | |
| P. aeruginosa | RND (MexEF-OprN) | 4-hydroxy-2-heptylquinoline (HHQ) | Quorum sensing quencher | [75] | |
| Biofilm-associated genes | Listeria monocytogenes | ABC (Lm.G_1771) | SrtA, Dlt, and GntR | Biofilm-associated gene suppression | [76] |
| Enterobacteriaceae | RND (AdeABC, AdeIJK) | csuA/B, csuC, and fimA | Adhesion and colonization interruption | [77] | |
| Acinetobacter baumannii | RND (AdeG) | abaI | AHL synthesis and transport | [78] | |
| Salmonella typhimurium | RND (AcrAB-TolC) | curli | Curli expression | [21] |
| Origin | Efflux Pump Inhibitor | Chemical Structure | Target Strain and Effective Substrate | Mechanism | Reference |
|---|---|---|---|---|---|
| Synthetic EPIs | PAβN |  | P. aeruginosa (MexAB-OprM transporters)—levofloxacin | Competitive inhibition, downregulation of efflux-related genes, and adjustment of membrane permeability | [17,99,101,105] |
| CCCP |  | A. baumannii—imipenem and cefepime | Interference with ATP synthesis and electrochemical gradients | [106,108] | |
| NMP |  | Enterobacteriaceae—oxacillin, linezolid, and rifampicin | Interruption of functional assembly of efflux pumps | [30,89,99,109] | |
| MBX2319 |  | E. coli—levofloxacin | Competitive inhibition and blockage of access to the substrate-binding sites | [30,110] | |
| Natural EPIs | Silybin |  | MRSA—ciprofloxacin and benzalkonium chloride | Downregulation of efflux-related genes | [111] |
| Curcumin |  | Clinical MRSA—ciprofloxacin | Downregulation of efflux-related genes | [93] | |
| Luteolin |  | T. pyogenes—macrolides | Interference with ATP synthesis and downregulation of efflux-related genes | [54] | |
| Boeravinone B |  | S. aureus—ciprofloxacin, EtBr | Interaction with the active sites of efflux pumps | [112] | |
| Baicalin |  | S. saprophyticus—EtBr | Interference with ATP synthesis | [74] | |
| Origanum vulgare L. EO | S. aureus—tetracycline | Downregulation of efflux-related genes | [113] | ||
| C. ambrosioides L. EO | MRSA—tetracycline and ethidium bromide | Disruption of the proton transport and adjustment of membrane permeability | [114] | ||
| Thymol and carvacrol | Gram-negative bacteria—tetracycline and benzalkonium chloride | Impairment of membrane integrity and induction of ion leakage | [115,116,117] | ||
| Salvia fruticosa EO | S. aureus—tetracycline | Downregulation of efflux-related genes | [118] | ||
| Origanum Majorana L. EO | S. aureus and E. coli—EtBr | Not mentioned | [119] | ||
| Nigella sativa EO | MRSA—tetracycline, ciprofloxacin, and EtBr | Downregulation of efflux-related genes | [120] | ||
| Cuminum cyminum L. EO | S. aureus—EtBr | Induction of conformational changes in efflux pump structures | [121] | ||
| Reserpine |  | B. subtilis—tetracycline S. aureus—norfloxacin | Interference with proton gradients and interaction with efflux pump proteins | [122,123,124] | |
| Berberine |  | P. aeruginosa—imipenem | Downregulation of efflux-related genes | [125] | |
| Jatrorrhizine |  | MRSA—norfloxacin | Downregulation of efflux-related genes | [126] | |
| Columbamine |  | E. coli, P. aeruginosa, and K. pneumoniae—streptomycin, erythromycin, norfloxacin, ampicillin, ciprofloxacin, doxycycline, and chloramphenicol | Interference with ATP synthesis and generation of proton electrochemical gradients | [98] | |
| Capsaicin |  | S. aureus—ciprofloxacin, EtBr | Docking to the active site | [127] | |
| Conessine |  | P. aeruginosa—cefotaxime, levofloxacin, tetracycline, erythromycin, novobiocin, and rifampicin | Competitive inhibition and/or blockage of access to the substrate-binding sites | [128] | |
| Catharanthine |  | P. aeruginosa—tetracycline and streptomycin | Docking to the active site | [129] | |
| Venturicidin A |  | Aminoglycoside-resistant MRSA—gentamicin | Interference with ATP synthesis and proton gradients | [130,131] | |
| EA-371α EA-371δ |  EA-371α EA-371α EA-371δ EA-371δ | P. aeruginosa—levofloxacin | Downregulation of efflux-related genes | [132] | |
| 2-(2-Aminophenyl) indole (RP2) |  | S. aureus—ciprofloxacin, tetracycline and erythromycin | Blockage of active site/channel | [133] | |
| Ethyl 4-bromopyrrole-2-carboxylate (RP1) |  | E. coli and P. aeruginosa—cloxacillin, ceftazidime, chloramphenicol, ciprofloxacin, erythromycin, levofloxacin, piperacillin, tetracycline, and tigecycline | Competitive inhibition | [134] | |
| 3,4-dibromopyrrole-2,5-dione |  | E. coli—ciprofloxacin, levofloxacin, kanamycin, erythromycin, piperacillin, tetracycline, and chloramphenicol | Not mentioned | [135] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Wang, J.; Ahn, J. Advances in the Discovery of Efflux Pump Inhibitors as Novel Potentiators to Control Antimicrobial-Resistant Pathogens. Antibiotics 2023, 12, 1417. https://doi.org/10.3390/antibiotics12091417
Zhang S, Wang J, Ahn J. Advances in the Discovery of Efflux Pump Inhibitors as Novel Potentiators to Control Antimicrobial-Resistant Pathogens. Antibiotics. 2023; 12(9):1417. https://doi.org/10.3390/antibiotics12091417
Chicago/Turabian StyleZhang, Song, Jun Wang, and Juhee Ahn. 2023. "Advances in the Discovery of Efflux Pump Inhibitors as Novel Potentiators to Control Antimicrobial-Resistant Pathogens" Antibiotics 12, no. 9: 1417. https://doi.org/10.3390/antibiotics12091417