1. Introduction
Antimicrobial resistance is one of the most pressing challenges facing modern medicine. However, the rate of discovery and introduction of new classes of antibiotics has slowed drastically over the decades, with only two introduced to the market since 1962 [
1]. The need for new antibiotic classes is only increasing as the potential for analogue development from existing classes is depleted. Pathogens that exhibit alarming resistance against current antimicrobial treatments are given the acronym of ESKAPE, including
Enterococcus faecium,
Staphylococcus aureus,
Klebsiella pneumoniae,
Acinetobacter baumannii,
Pseudomonas aeruginosa, and
Enterobacter species [
2]. Among them, methicillin and vancomycin-resistant
Staphylococcus aureus (MRSA/VRSA) are placed in the high-priority category by the World Health Organization to accelerate the development of new antimicrobial compounds for treating multi-drug-resistant
S. aureus [
3]. MRSA is usually acquired within a hospital environment and can cause various severe opportunistic infections and diseases [
4]. Vancomycin has long been regarded as a drug of last resort for MRSA treatment. Unfortunately, the emerging VRSA jeopardises the efficiency of clinical treatments and leads to a significant increase in staphylococcal bacteraemia mortality [
5].
Bacteriophages, especially lytic phages, are viruses that infect, propagate, and destroy bacteria. Throughout evolution, phages continuously adapt to confront their bacterial hosts for their own survival. This intertwined relationship endows the phages with highly specific mechanisms to reprogram bacterial cell metabolism. Phage-inspired antibacterial target discovery is another ascending approach to harnessing the antimicrobial activity of phages [
6]. Identifying novel bacterial targets through phage research could help screen and design new small molecular compounds that replicate the growth-inhibitory effects of the antibacterial phage proteins. Intriguingly, a substantial proportion of phage gene products have completely unknown functions since they have not been characterized and have sequences that do not correspond with any proteins of known function. Screening hypothetical proteins of unknown function (HPUFs) from bacteriophages for toxic activity against bacteria may provide new and potentially life-saving approaches to combat bacterial infections [
7,
8]. An earlier extensive study on mining 26
S. aureus phage genomes revealed that 31 polypeptide families showed toxicity towards the bacterial host, including ORF104 from phage 77 [
5]. The gene product of ORF104 interacted with bacterial essential protein DnaI, disrupting DNA synthesis and cell growth. The research team then used the protein pair to screen for small molecular inhibitors, and identified 36 of them interrupting the ORF104-DnaI interaction and 11 that were directly toxic against
S. aureus [
5].
Stab21 is a lytic
Staphylococcus phage recently discovered by Oduor et al. [
9]. The phage is taxonomically classified to the
Kayvirus genus of the Twortvirinae subfamily in the Herelleviridae family (accession number LR215719, [
9]). Stab21 has a wide host range and possesses no genes associated with antibiotic resistance or virulence [
9]; therefore, it is a good candidate for therapeutical applications. Of all 238 predicted genes in the Stab21 genome, 203 were preliminarily annotated to encode HPUFs. Many lytic phages could possess genes encoding proteins that directly or indirectly mediate the destruction of their host bacterial cells [
10]. Hence, among the HPUFs of Stab21, there is a great potential for the discovery of bactericidal proteins that alter bacterial pathways in an unprecedented manner.
The use of a next-generation sequencing (NGS)-based screening approach to screen for phage-encoded toxic proteins is shown to be as reliable as the alternative plating-based toxicity screening method. An earlier direct comparison was performed during the development of the NGS method, where it was concluded that the NGS-based assay not only provides similar screening results as the plating-based assays, but is also superior in efficiency, accuracy, and reliability [
11,
12].
In the study presented here, we investigated all hypothetical proteins and selected 96 true HPUFs for their bactericidal activity in E. coli using the NGS-based approach. We confirmed their bactericidal activities in E. coli and their potential anti-S. aureus abilities using various inducible expression systems. Five of the Stab21 gene products were shown to have cross-species toxicity towards E. coli. Through detailed in silico functional and structural analyses, we found that Gp024 shares high structural homology with a homing endonuclease. Our work not only underscores the potential of Stab21 HPUFs as antimicrobial agents, but also reinforces the efficacy of NGS in identifying these potential assets.
3. Discussion
The escalating prevalence of antibiotic resistance in critical pathogens such as
S. aureus demands the discovery of novel antimicrobial molecules [
20]. Phages, with their unique genetic diversity, present a promising reservoir of potential antimicrobials, yet the majority of their genomes remain uncharacterized. Current phage research identifies a range of bacteriotoxic molecules such as endolysins and polysaccharide depolymerases that could potentially be harnessed against antibiotic resistant bacterial infections [
21,
22,
23]. Given the wide genomic variation even among phages infecting the same host, there is significant potential for the discovery of new antimicrobial mechanisms and targets from the multitude of gene products currently deemed HPUF [
24]. Such discoveries may yield innovative antibiotic treatments capable of countering the ever-increasing threat of antimicrobial resistance [
6].
In this study, we employed an NGS-based screening assay to identify toxic proteins encoded by Staphylococcal phage Stab21. The HPUFs of Stab21 were screened for bacteriotoxic effects against E. coli and their bactericidal activities were confirmed in both E. coli and S. aureus. Our findings revealed that five gene products were toxic to E. coli DH10B using arabinose- and anhydrotetracycline-induced expression vectors. Crucially, this study verified that the application of this high-throughput NGS-based screening process in E. coli is not limited to examining HPUFs from only phages infecting Gram-negative bacteria.
Our study employed an already established protocol for NGS-based screening in
E. coli [
11,
12], recognizing the value of this model organism in providing insights into cellular mechanisms that transcend species boundaries [
25]. By adopting this protocol, we anticipate a rapid determination of any bacteriotoxic activity of HPUFs in both Gram-negative and Gram-positive strains.
In this context, we constructed the pCU1LK vector which contains the backbone of the
E. coli–
S. aureus shuttle vector pCU1 [
26] that can accommodate up to 6 kb fragments cloned under the
lac promoter. As the size of the inserted linker sequence was only 45 bp in pCU1LK, it should also allow cloning up to 6 kb fragments without problems to replicate freely in both hosts.
However, as the
lac promoter is notoriously known as leaky, we selected another tighter expression vector pRAB11 with two
tet operator
tetO sites [
27] to verify the toxicity of HPUFs. Nevertheless, we observed an unexpected growth inhibition of
S. aureus strain RN4220/pRAB11N in the presence of ATc on solid media (
Figures S3 and S4). This effect was not noticed in
E. coli DH10B/pRAB11N. We do not have a clear explanation for this phenomenon. Despite the expectation that ATc should induce the gene expression at lower concentrations and with less toxicity than its parent antibiotic, tetracycline (Tc) [
28], aged ATc has been found to generate toxic breakdown products that can interfere with
S. aureus growth [
29]. It is possible that these breakdown products are less effective on
E. coli than on
S. aureus, as observed in our study.
Though our study did not identify any toxHPUFs towards
S. aureus among the nine
E. coli -toxic candidates, further examination should be carried out for the remaining 87 HPUFs of Stab21, which were regarded as non-toxic to
E. coli. That could involve pooling pCU1LK-HPUFs or pRAB11N-HPUFs ligation mixtures and subsequently screening successful transformants for toxicity-presenting an alternative to the NGS-based screening method. Alternatively, an arsenite-inducible plasmid, pT0021, which has been previously employed effectively to screen for toxHPUFs in
S. aureus [
7], could serve as another viable option for this investigation.
Furthermore, for future screening of HPUFs from various phages derived from different bacteria, the NGS screening technique presented in our study can be fine-tuned depending on the bacterial host by applying different inducible plasmid vectors such as the Tn7-based integration vector pTNS2 [
30] for
Pseudomonas aeruginosa, or vectors derived from the IncQ plasmid for
Acinetobacter baumannii [
31].
Among the structural studies of the top five identified toxic HPUFs, only Gp024 yielded a reliable protein model, aligning with the
Bacillus phage SPO1 homing endonuclease I-HmuI. Although the predicted structure of Gp024 resembles different domains in HHpred and Phyre2, its function as a homing endonuclease cannot be completely ignored. Previous studies have identified phage HPUFs as homologs to homing endonucleases. A noteworthy example is found in the extensively researched
Escherichia phage T4, which houses 15 homing endonuclease encoding genes, showing the evolutionary link between these proteins and phage biology [
32]. Adding to the intrigue is the existence of colicins, DNases produced by
E. coli that demonstrate toxicity against other
E. coli strains. These colicins belong to the same H-N-H family of endonucleases [
33] as the suspected homing endonuclease Gp024 of Stab21. The analogous functions and structural similarity of these proteins could suggest that Gp024 could exhibit characteristics of homing endonucleases like colicins. However, further experiments such as single-site mutations and DNA nicking assays are needed to confirm its role as a homing endonuclease.
Our findings that five HPUFs from Stab21 exhibit cross-species toxicity towards E. coli have profound implications. It is possible that these HPUFs share a conserved molecular target in both E. coli and S. aureus, significantly broadening the potential applicability of these bactericidal agents. In conclusion, our research opens a promising avenue for the discovery of novel, potent antimicrobial agents, providing hope in the fight against growing antimicrobial resistance.
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, Phage and Culture Conditions
All bacterial strains and plasmids used for experimentation are listed in
Table 1 and
Table S2. Commercial electrocompetent
Escherichia coli DH10B cells (Thermo Fischer Scientific, Waltham, MA, USA), in-house prepared electrocompetent
E. coli DH5ɑ and in-house prepared electrocompetent
Staphylococcus aureus RN4220 were used as expression hosts.
E. coli DH5ɑ, DH10B and their derivatives were grown in Lysogeny broth (LB; 10 g/L Tryptone (Neogen, Lansing, MI, USA, Cat no. NCM02118A), 5 g/L Yeast Extract (Neogen, Cat no. NCM0218A), 10 g/L NaCl) or on agar (LA, LB supplemented with 1.5% Bacto agar). For toxicity tests, E. coli strains were grown in M9 minimal media (KH2PO4 3 g/L, 0.5 g/L NaCl, 6.78 g/L Na2HPO4, 1 g/L NH4Cl, casamino acid 0.2% (v/v), MgSO4 2 mM, CaCl2 0.1 mM, thiamine 1 mg/L) supplemented with antibiotic to maintain the plasmid, and glucose for repression and arabinose for induction. S. aureus strains were grown in Tryptic Soy Broth (TSB; VWR Chemicals, Radnor, PA, USA, Cat. No. 470015-844) or on Tryptic Soy Agar (TSA, Vegitone, Sigma-Aldrich, St. Louis, MA, USA, Cat. No. 14432) or broth (Dehydrated TSB, VWR Chemicals). Liquid cultures were grown at 37 °C overnight with 200 RPM shaking unless stated otherwise. Solid cultures were incubated at 37 °C overnight. To maintain the plasmids, broth or agar was supplemented with either 100 µg/mL of ampicillin (Amp100) or 30 µg/mL of chloramphenicol (Cm30) unless stated otherwise.
4.2. DNA Manipulations
For plasmid isolations, individual colonies were obtained on a streak plate, and 1 colony was used for inoculation to obtain overnight cultures. Plasmids from E. coli strains were extracted, purified, and precipitated with either NucleoBond™ Xtra Midi kit and NucleoBond™ Finalizers (Machery-Nagel, Düren, Germany) from 200 mL overnight cultures or with the NuceloSpin Plasmid EasyPure Kit (Machery-Nagel) for 1 mL cultures, according to manufacturers’ instructions. To isolate the plasmids from Gram-positive S. aureus, lysostaphin lyophilized powder from Staphylococcus staphylolyticus (Merck KGaA, Darmstadt, Germany) was added to the final concentration of 20 µg/mL and incubated for 1 h at 37 °C before using NucleoSpin Plasmid EasyPure Kit according to the manufacturer’s manual. For the Illumina sequencing, plasmid pools were extracted with NucleoBond™ Xtra Midi kit, and eluted in a 200 µL Tris/HCl pH 8.5 elution buffer.
E. coli and S. aureus genomic DNA was isolated using the JetFlex Genomic DNA Purification Kit (Thermo Fischer Scientific) following the bacterial gDNA isolation protocol for Gram-negative and Gram-positive bacteria, the latter using a 20 µg/mL lysostaphin pre-treatment. The isolated DNA was rehydrated at 22 °C for 16 h.
DNA fragments were amplified by PCR using primers listed in
Table S3. Stab21 phage DNA [
14] was used as a template for amplification of HPUFs.
S. aureus Newman gDNA was used to amplify the toxic control gene ORF104. Plasmid DNA and colonies were used as templates for confirmation of correct gene insertion. Phusion High-Fidelity DNA Polymerase (Thermo Fischer Scientific) was used for generating DNA fragments with the highest accuracy for cloning, while DreamTaq DNA Polymerase (Thermo Fisher Scientific) was used for screening the presence of a certain DNA fragment either from the colony or from the ligation mixture. The PCRs were run in a T100™ or iCycler Thermal Cycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA) following standard manufacturer protocol for the polymerases.
All restriction enzymes used in this study were obtained from Thermo Fisher Scientific or New England Biolabs (MA, USA). Plasmid vectors were linearised with restriction enzymes as stated according to the manufacturer’s instructions, and dephosphorylated with FastAP™ Thermosensitive Alkaline Phosphatase (Thermo Fisher Scientific) at 37 °C for 30 min followed by a 15 min heat inactivation at 65 °C.
Sticky-end ligation of double-digested individual HPUF-encoding gene fragments to linearised and dephosphorylated pCU1LK or pRAB11N vectors was carried out at a 1:3 vector to insert molar ratio, and the total DNA concentration was adjusted to 10 ng/µL. T4 DNA Ligase (5 U) (Thermo Fischer Scientific) was used for all the ligation reactions. The ligation reaction was incubated at room temperature overnight (15 h) before heat inactivation at 65 °C for 10 min.
NucleoSpin Gel and PCR Clean-up XS kit (Machery-Nagel) was used to purify and concentrate DNA after PCRs and enzymatic reactions.
For the NGS screening assay, the Stab21 HPUF-encoding genes and pCU1LK vector were first double digested with restriction enzymes NotI and NheI or KpnI FastDigest™ enzymes (Thermo Fisher Scientific) depending on the insertion fragments (
Table S3) [
11]. Every 16 ligation mixtures of the HPUF gene and vector pCU1LK were pooled before concentration by kit, and an elution volume of 20 µL in Baxter Sterile Water (Baxter Corporation, Deerfield, IL, USA) was used per pool. One microliter (ca. 200 ng) of each ligation pool was transferred to 50 µL of
E. coli DH10B cells through electroporation. Plasmids from transformation reactions were isolated from a 3 h culture inoculated with all colonies formed on the transformation plates. DNA samples from both the ligation pool and the plasmid pool were sequenced with the 150 bp paired-end protocol in the Illumina HiSeq platform at NovoGene Company Ltd. (Cambridge, UK) as described by Kasurinen et al. [
11].
4.3. Electroporation and Transformation
Electroporation was performed with a Gene Pulser™ apparatus (Bio-Rad Laboratories) using 0.2 mm cuvettes. For the transformation of E. coli strains, the parameters of 200 Ω resistance, 25 mF capacitance and 2.5 kV voltage resulted in a time constant between 4.5 and 5.0 ms. Transformed E. coli cells were recovered in a 1 mL super optimal broth (SOC; 2% Tryptone, 0.5% Yeast Extract, 10 mM NaCl, 2.5 mM KCI, 10 mM MgCl2, 10 mM MgSO4, 20 mM glucose) and incubated at 37 °C with a 200 rpm shaking for 45 min before being plated on LB Amp100 agar plates using 10 µL, 100 µL, and the remainder of cells collected through centrifugation. For NGS screening, every 50 µL of the recovered cells from each pool were spread onto LB Amp100 plates, resulting in 20 plates. The plates were incubated at 37 °C overnight.
For the transformation of S. aureus RN4220, the parameters of the Gene Pulser electroporator were set with a resistance of 100 Ω, a capacitance of 25 µF, and a voltage of 2.3 kV resulting in a time constant between 2.0 and 2.4 ms. Transformed S. aureus were recovered in a 850 µL TSB and a 150 µL 2 M sucrose and incubated at 37 °C with a 200 rpm shaking for 90 min. The same plating scheme and incubation conditions were used as those for E. coli electroporation.
4.4. Construction of Vectors pCU1LK and pRAB11N
To construct a vector suitable for screening the toxicity of all the 96 HPUFs in
E. coli, we added a linker to the pCU1 plasmid. The KpnI-PstI linker containing restriction sites for BamHI, XbaI, NheI, NcoI, and NotI was constructed by annealing oligonucleotides NOTup and NOTdown (
Table S3) with a final concentration of 50 μM in a 20 μL linker solution (50 mM Tris-HCl pH 8.0, 100 mM NaCl, 1 mM EDTA). The reaction mixture was incubated at 95 °C for 2 min, followed by 10 min at 52 °C. The annealed linker was phosphorylated with T4 polynucleotide kinase (T4 PNK, Thermo Fisher Scientific) before ligation to gel-purified pCU1 linearised with KpnI and PstI digestions. The resultant vector pCU1LK was used for the preliminary screening of the toxicity of all HPUFs in
E. coli.
The
E. coli–S. aureus shuttle vector pRAB11N was re-constructed to make an exact copy of the plasmid pRAB11, which could no longer be obtained from any source. The plasmid pRMC2 [
34] was used as a template in plasmid-PCR using pRAB-fw and pRAB-rev as primers (
Table S3). This PCR added a second
tet operator to the
tetR promoter region. A 30 µL aliquot of the obtained PCR product was digested with a 50 U DpnI (New England BioLabs, cat. no. R0176) in a reaction volume of 50 µL to eliminate the pRMC2 template before ligation. One of the confirmed transformants was named pRAB11N, and it was used as a shuttle expression vector in both
E. coli and
S. aureus.
The correctness of the obtained plasmid vectors was confirmed by Sanger sequencing at the Finnish Institute for Molecular Medicine (FIMM) Genomics Sequencing (Biomedicum, Tukholmankatu 8, Helsinki, Finland) using primers Puc19-F and Puc19-R for pCU1LK, and fR-346 for pRAB11N. Primers used in this study are listed in
Table S3.
4.5. Bioinformatics
The NGS-based toxicity screening of HPUF encoding genes was carried out using the protocol described previously [
11]. The DNA samples used for NGS are described in
Section 2.3. For each pool of 16 HPUFs, the reads containing the four expected ligation joint sequences (VGF, GVF, VGR, and GVR,
Figure 7, and
Table S1) were identified and extracted from both the pooled ligation mixture DNA and the pooled plasmid DNA samples using the script and workflow described earlier [
11]. The total number of the four ligation joint sequences for each HPUF-encoding gene was calculated and used to represent their total read coverage (N joint reads, Formula (1)). The relative number of joint sequence reads was calculated for all genes in the pools by dividing the total read coverage of a single gene by the total number of joint sequence reads for all genes in the pool and expressed as a percentage (relative gene percentage, Formula (1)). As described by Kasurinen et al., a low ratio (Formula (2)) between the relative joint sequence reads of individual genes from plasmid pools and those from the corresponding ligation mixture indicates the presence of toxHPUF gene, owing to the elimination of transformants carrying a toxic gene [
11].
On the contrary, a ratio close to 1 or above indicates a non-toxic gene, reflecting the successful replication of the recombinant plasmid. In theory, a HPUF could be considered as bactericidal if this ratio is less than 1. In our study presented here, gene products exhibiting ratios between 0.5 and 1.0 were considered potentially mildly toxic, while those with a ratio under 0.5 were potentially bactericidal.
4.6. Toxicity Confirmation
The toxicity of the potentially toxic HPUFs towards host bacteria was primarily tested using a drop test on agar plates. Potentially toxic HPUFs genes were cloned either into the pBAD33 vector under an arabinose-inducible promoter (using the KpnI-XbaI restriction sites) or pRAB11N under an ATc-inducible promoter (using the BglII-KpnI sites). The correct gene insertions were confirmed by colony PCR using primers pBADF and pBADRev for pBAD33 constructions and pRAB11-F and fR-346 for pRAB11N constructions. The resultant PCR products were further analysed and identified by Sanger sequencing.
Subsequently,
E. coli or
S. aureus transformants carrying the desired recombinant plasmids (
Table S2) were grown on suitable agar plates to obtain single colonies. Three isolated colonies containing the recombinant vector and positive-control colonies were resuspended and diluted to optical density at 600 nm (OD
600) 0.2 with sterile phosphate-buffered saline (PBS) pH 7.4. Five microliters of the serial dilutions from 10
−1 to 10
−8 of each culture were spotted onto both induced and non-induced LA or TSA plates and let dry before incubation at 37 °C overnight. Varying inducing and non-inducing conditions of the plates are listed in
Table S4.
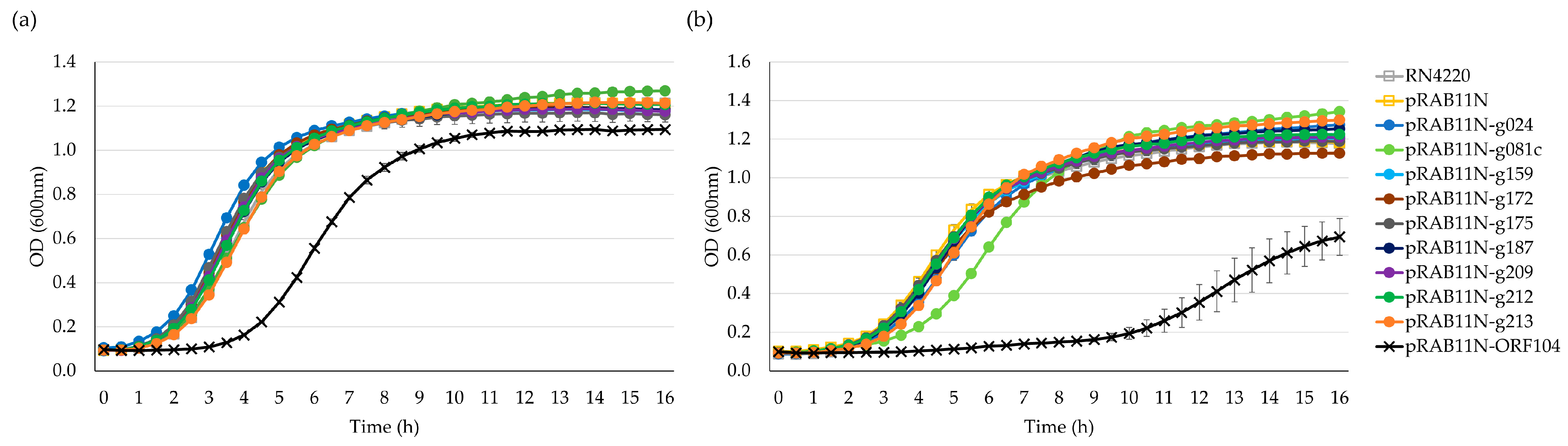
To confirm toxicity of the HPUFs and visualise the time-specific impact of toxHPUFs, the growth curves of the strains carrying the pRAB11N-HPUF plasmids were determined with and without induction. Three colonies of each strain were inoculated in a 1 mL TSB supplemented with Amp100 for E. coli, and TSB supplemented with Cm30 for S. aureus and incubated at 37 °C overnight with a 200 RPM shaking. Cells were resuspended and washed with an equal volume of TSB, and OD600 was measured. The washed cells were used as a starting inoculum after resuspending to an OD600 of 0.01 into fresh TSB, with and without the inducer (ATc 0.4 µM). The bacteria were grown in Bioscreen Honeycomb plates (Oy Growth Curves Ab Ltd., Helsinki, Finland) in triplicate. The OD600 was measured using the Bioscreen C MBR (Oy Growth Curves Ab Ltd., Helsinki, Finland) every 30 min for 16 h with settings of continuous shaking, high amplitude, and normal speed. The shaking was stopped 10 s before each OD600 measurement. The mean values and standard deviations were calculated using triplicate data points.
4.7. Genomic DNA Sequencing
Bacterial gDNA isolated from
E. coli DH10B/pRAB11N-HPUF clones was sequenced at Novogene Company Ltd. Contigs were assembled using BV-BRC (
https://www.bv-brc.org/app/Assembly2, accessed on 26 June 2023), and a nucleotide BLAST (Basic Local Alignment Search Tool, version 2.14.0,
https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 26 June 2023) search was used to align the sequence assemblies with the individual pRAB11N-HPUF and
E. coli DH10B genome sequences. Protein BLAST was used to align sequences to known proteins.
4.8. Structural and Functional Analysis of Toxic Proteins
Phyre2, HHpred, and AlphaFold2 were used to predict the functions and structures of the toxic hypothetical proteins. Comparisons were made between the sequence identities of the proteins and known protein structures, with cut-offs of 30% identity and confidence levels over 90% for Phyre2 and similar parameters for HHpred. Functional and structural protein database files were superimposed using the molecular visualisation system PyMOL utilising the ‘super’ function (The PyMOL Molecular Graphics System, version 2.5, Schrödinger, LCC). RMSD scores were calculated using PyMOL to measure structural alignment and overlap of the predicted protein structures.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}