

High Emergence of Multidrug-Resistant Sequence Type 131 Subclade C2 among Extended-Spectrum β-Lactamase (ESBL)-Producing Escherichia coli Isolated from the University Hospital Bratislava, Slovakia
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Patients’ Characteristics and Isolation of ESBL-Producing E. coli Strains
2.2. Occurrence of ST131 and non-ST131 ESBL-Producing E. coli Strains
2.3. Distribution of Clades and Subclades of ST131 ESBL-Producing E. coli Strains
2.4. Genetic Relatedness of ESBL-Producing Sequenced E. coli Strains
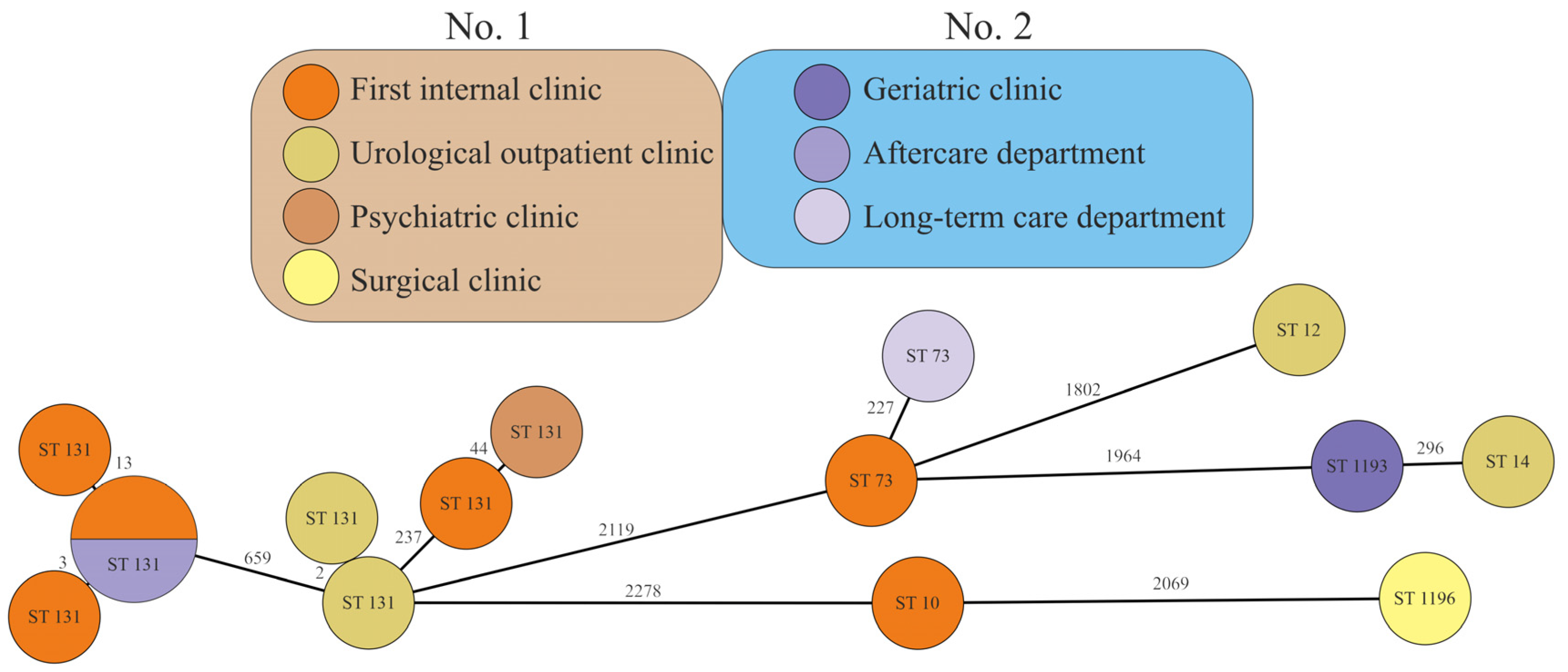
2.5. Occurrence of STs in Healthcare Facilities
2.6. Antibiotic Resistance of MDR and ESBL-Producing E. coli Strains
3. Discussion
4. Materials and Methods
4.1. Healthcare Facilities, Patients, and Specimens
4.2. Isolation, Proof of Bacterial Strains, and Susceptibility Testing
4.3. DNA Isolation and NGS Library Preparation
4.4. ST-131 Clade Multiplex PCR with fimH-30 Detection
4.5. Bioinformatic Processing and Genome Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tchesnokova, V.; Riddell, K.; Scholes, D.; Johnson, J.R.; Sokurenko, E.V. The Uropathogenic Escherichia coli Subclone Sequence Type 131-H30 Is Responsible for Most Antibiotic Prescription Errors at an Urgent Care Clinic. Clin. Infect. Dis. 2019, 68, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Gutiérrez, B.; Rodríguez-Baño, J. Current options for the treatment of infections due to extended-spectrum beta-lactamase-producing Enterobacteriaceae in different groups of patients. Clin. Microbiol. Infect. 2019, 8, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Geisinger, E.; Isberg, R.R. Interplay Between Antibiotic Resistance and Virulence During Disease Promoted by Multidrug-Resistant Bacteria. J. Infect. Dis. 2017, 215, S9–S17. [Google Scholar] [CrossRef] [Green Version]
- Tamma, P.D.; Rodriguez-Baňo, J. The Use of Noncarbapenem β-Lactams for the Treatment of Extended-Spectrum β-Lactamase Infections. Clin. Infect. Dis. 2017, 64, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Etayo, L.; González, D.; Vitas, A.I. Clonal Complexes 23, 10, 131 and 38 as Genetic Markers of the Environmental Spread of Extended-Spectrum β-Lactamase (ESBL)-Producing E. coli. Antibiotics 2022, 11, 1465. [Google Scholar] [CrossRef] [PubMed]
- Soncini, J.G.M.; Cerdeira, L.; Sano, E.; Koga, V.L.; Tizura, A.T.; Tano, Z.N.; Nakazato, G.; Kobayashi, R.K.T.; Aires, C.A.M.; Lincopan, N.; et al. Genomic insights of high-risk clones of ESBL-producing Escherichia coli isolated from community infections and commercial meat in southern Brazil. Sci. Rep. 2022, 12, 9354. [Google Scholar] [CrossRef] [PubMed]
- Isendahl, J.; Giske, C.G.; Hammar, U.; Sparen, P.; Tegmark Wisell, K.; Ternhag, A.; Nauclér, P. Temporal Dynamics and Risk Factors for Bloodstream Infection With Extended-spectrum β-Lactamase-producing Bacteria in Previously-colonized Individuals: National Population-based Cohort Study. Clin. Infect. Dis. 2019, 68, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Besser, T.E.; Call, D.R.; Weissman, S.J.; Jones, L.P.; Davis, M.A. Evaluation of two multi-locus sequence typing schemes for commensal Escherichia coli from dairy cattle in Washington State. J. Microbiol. Methods 2016, 124, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Ludden, C.; Coll, F.; Gouliouris, T.; Restif, O.; Blane, B.; Blackwell, G.A.; Kumar, N.; Naydenova, P.; Crawley, C.; Brown, N.M.; et al. Defining nosocomial transmission of Escherichia coli and antimicrobial resistance genes: A genomic surveillance study. Lancet Microbe 2021, 2, e472–e480. [Google Scholar] [CrossRef]
- Matsumura, Y.; Pitout, J.D.D.; Peirano, G.; DeVinney, R.; Noguchi, T.; Yamamoto, M.; Gomi, R.; Matsuda, T.; Nakano, S.; Nagao, M.; et al. Rapid Identification of Different Escherichia coli Sequence Type 131 Clades. Antimicrob. Agents Chemother. 2017, 61, e00179-17. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.P.; Kaye, K.S.; Evans, R.; Salzman, E.; Pogue, J.; Hayakawa, K.; Marchaim, D.; Awasthy, P.; Salim, M.; Martin, E.T. Clinical and Molecular Epidemiology of Extended-Spectrum Beta-Lactamase-Producing Escherichia Coli Infections in Metro Detroit: Early Dominance of the ST-131 Clone. Infect. Dis. Ther. 2020, 9, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Lovayová, V.; Čurová, K.; Hrabovský, V.; Nagyová, M.; Siegfried, L.; Toporová, A.; Rimárová, K.; Andraščíková, Š. Antibiotic resistance in the invasive bacteria Escherichia coli. Cent. Eur. J. Public Health 2022, 30, S75–S80. [Google Scholar] [CrossRef]
- Hrabák, J.; Empel, J.; Bergerová, T.; Fajfrlík, K.; Urbásková, P.; Kern-Zdanowicz, I.; Hryniewicz, W.; Gniadkowski, M. International clones of Klebsiella pneumoniae and Escherichia coli with extended-spectrum beta-lactamases in a Czech hospital. J. Clin. Microbiol. 2009, 47, 3353–3357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafolie, J.; Nicolas-Chanoine, M.H.; Grenouillet, F.; Hocquet, D.; Bertrand, X. Prevalence of Escherichia coli sequence type 131 and its H30 subclone among E. coli isolates in a French hospital. Int. J. Antimicrob. Agents 2014, 44, 466–468. [Google Scholar] [CrossRef]
- Roer, L.; Johannesen, T.B.; Hansen, F.; Stegger, M.; Tchesnokova, V.; Sokurenko, E.; Garibay, N.; Allesøe, R.; Thomsen, M.C.F.; Lund, O.; et al. CHTyper, a Web Tool for Subtyping of Extraintestinal Pathogenic Escherichia coli Based on the fumC and fimH Alleles. J. Clin. Microbiol. 2018, 56, e00063-18. [Google Scholar] [CrossRef] [Green Version]
- Boll, E.J.; Overballe-Petersen, S.; Hasman, H.; Roer, L.; Ng, K.; Scheutz, F.; Hammerum, A.M.; Dungu, A.; Hansen, F.; Johannesen, T.B.; et al. Emergence of Enteroaggregative Escherichia coli within the ST131 Lineage as a Cause of Extraintestinal Infections. mBio 2020, 11, e00353-20. [Google Scholar] [CrossRef]
- Martischang, R.; François, P.; Cherkaoui, A.; Gaïa, N.; Renzi, G.; Agostinho, A.; Perez, M.; Graf, C.E.; Harbarth, S. Epidemiology of ESBL-producing Escherichia coli from repeated prevalence studies over 11 years in a long-term-care facility. Antimicrob. Resist. Infect. Control 2021, 10, 148. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, A.; Kiszakiewicz, C.; Kristiansson, E.; Yazdanshenas, S.; Kamenska, N.; Karami, N.; Åhrén, C. The impact of the ST131 clone on recurrent ESBL-producing E. coli urinary tract infection: A prospective comparative study. Sci. Rep. 2022, 12, 10048. [Google Scholar] [CrossRef]
- Mhaya, A.; Trabelsi, R.; Aillerie, S.; M’Zali, F.; Bégu, D.; Tounsi, S.; Gdoura, R.; Arpin, C. Detection of Clones B2-ST131-C2 and A-ST617 in Escherichia coli Producing Both CTX-M-15 and CTX-M-27 from Tunisian Community Patients. Antibiotics 2022, 11, 1329. [Google Scholar] [CrossRef]
- Ellaby, N.; Doumith, M.; Hopkins, K.L.; Woodford, N.; Ellington, M.J. Emergence of diversity in carbapenemase-producing Escherichia coli ST131, England, January 2014 to June 2016. Eurosurveillance 2019, 24, 1800627. [Google Scholar] [CrossRef] [Green Version]
- Olesen, B.; Hansen, D.S.; Nilsson, F.; Frimodt-Møller, J.; Leihof, R.F.; Struve, C.; Scheutz, F.; Johnston, B.; Krogfelt, K.A.; Johnson, J.R. Prevalence and characteristics of the epidemic multiresistant Escherichia coli ST131 clonal group among extended-spectrum beta-lactamase-producing E. coli isolates in Copenhagen, Denmark. J. Clin. Microbiol. 2013, 51, 1779–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weterings, V.; van den Bijllaardt, W.; Bootsma, M.; Hendriks, Y.; Kilsdonk, L.; Mulders, A.; Kluytmans, J. Duration of rectal colonization with extended-spectrum beta-lactamase-producing Escherichia coli: Results of an open, dynamic cohort study in Dutch nursing home residents (2013–2019). Antimicrob. Resist. Infect. Control 2022, 11, 98. [Google Scholar] [CrossRef]
- Feng, Y.C.; Liou, C.H.; Ng, W.V.; Chen, F.J.; Hung, C.H.; Liu, P.Y.; Liao, Y.C.; Wu, H.C.; Cheng, M.F. Distribution and Genomic Characterization of Third-Generation Cephalosporin-Resistant Escherichia coli Isolated from a Single Family and Home Environment: A 2-Year Longitudinal Study. Antibiotics 2022, 11, 1152. [Google Scholar] [CrossRef] [PubMed]
- van den Bunt, G.; Fluit, A.C.; Bootsma, M.C.J.; van Duijkeren, E.; Scharringa, J.; van Pelt, W.; Bonten, M.J.M. Dynamics of Intestinal Carriage of Extended-Spectrum Beta-lactamase-Producing Enterobacteriaceae in the Dutch General Population, 2014–2016. Clin. Infect. Dis. 2020, 71, 1847–1855. [Google Scholar] [CrossRef] [PubMed]
- Flament-Simon, S.C.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Nicolas-Chanoine, M.H.; Blanco, J. High Prevalence of ST131 Subclades C2-H30Rx and C1-M27 among Extended-Spectrum β-Lactamase-Producing Escherichia coli Causing Human Extraintestinal Infections in Patients from Two Hospitals of Spain and France during 2015. Front. Cell. Infect. Microbiol. 2020, 10, 125. [Google Scholar] [CrossRef] [PubMed]
- Tchesnokova, V.L.; Rechkina, E.; Larson, L.; Ferrier, K.; Weaver, J.L.; Schroeder, D.W.; She, R.; Butler-Wu, S.M.; Aguero-Rosenfeld, M.E.; Zerr, D.; et al. Rapid and Extensive Expansion in the United States of a New Multidrug-resistant Escherichia coli Clonal Group, Sequence Type 1193. Clin. Infect. Dis. 2019, 68, 334–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, L.W.; Hoi, L.T.; Khokhar, F.A.; Hoa, N.T.; Giang, T.V.; Bui, C.; Ninh, T.H.; Co, D.X.; Binh, N.G.; Long, H.B.; et al. Genomic characterisation of multidrug-resistant Escherichia coli, Klebsiella pneumoniae, and Acinetobacter baumannii in two intensive care units in Hanoi, Viet Nam: A prospective observational cohort study. Lancet Microbe 2022, 3, e857–e866. [Google Scholar] [CrossRef]
- Sadek, M.; Saad, A.M.; Nordmann, P.; Poirel, L. Genomic Characterization of an Extensively Drug-Resistant Extra-Intestinal Pathogenic (ExPEC) Escherichia coli Clinical Isolate Co-Producing Two Carbapenemases and a 16S rRNA Methylase. Antibiotics 2022, 11, 1479. [Google Scholar] [CrossRef]
- MacFadden, D.R.; Melano, R.G.; Coburn, B.; Tijet, N.; Hanage, W.P.; Daneman, N. Comparing Patient Risk Factor-, Sequence Type-, and Resistance Locus Identification-Based Approaches for Predicting Antibiotic Resistance in Escherichia coli Bloodstream Infections. J. Clin. Microbiol. 2019, 57, e01780–e18. [Google Scholar] [CrossRef] [Green Version]
- Cummins, E.A.; Snaith, A.E.; McNally, A.; Hall, R.J. The role of potentiating mutations in the evolution of pandemic Escherichia coli clones. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1–10. [Google Scholar] [CrossRef]
- van Hout, D.; Verschuuren, T.D.; Bruijning-Verhagen, P.C.J.; Bosch, T.; Schürch, A.C.; Willems, R.J.L.; Bonten, M.J.M.; Kluytmans, J.A.J.W. Extended-spectrum beta-lactamase (ESBL)-producing and non-ESBL-producing Escherichia coli isolates causing bacteremia in the Netherlands (2014–2016) differ in clonal distribution, antimicrobial resistance gene and virulence gene content. PLoS ONE 2020, 15, e0227604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 13.0. 2023. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_13.0_Breakpoint_Tables.pdf (accessed on 2 January 2023).
- Nordmann, P.; Poirel, L.; Dortet, L. Rapid Detection of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2012, 18, 1503–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gattringer, R.; Nikš, M.; Ostertág, R.; Schwarz, K.; Medvedovic, H.; Graninger, W.; Georgopoulos, A. Evaluation of MIDITECH automated colorimeter MIC reading for antimicrobial susceptibility testing. J. Antimicrob. Chemother. 2002, 49, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Prjibelski, A.D.; Antipov, D.; Meleshko, D.; Lapidus, A.L.; Korobeynikov, A.I. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.F.; Mohamed, K.; Fan, Y.; Agama Study Group; Achtman, M. The EnteroBase user’s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny, and Escherichia core genomic diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Total of (354) | ST131 (263) | Non-ST131 (91) | |||||
|---|---|---|---|---|---|---|---|
| Gender and Age | n | % | n | % | n | % | |
| Male | 119 | 33.6 | 86 | 32.7 | 33 | 36.3 | |
| Mean years (range) 75.0 ± 14.4 (25–96) | |||||||
| Median years 79 | |||||||
| Female | 235 | 66.4 | 177 | 67.3 | 58 | 63.7 | |
| Mean years (range) 80.4 ± 11.9 (9–98) * | |||||||
| Median years 83 | |||||||
| ≥65 years | 310 | 87.6 | 236 | 89.7 ** | 74 | 81.3 | |
| ˂65 years | 44 | 12.4 | 27 | 10.3 | 17 | 18.7 | |
| Setting | |||||||
| No. 1 | First internal clinic | 190 | 53.7 | 142 | 54.0 | 48 | 52.7 |
| Surgical clinic | 20 | 5.6 | 13 | 4.9 | 7 | 7.7 | |
| Neurological clinic | 7 | 2.0 | 6 | 2.3 | 1 | 1.1 | |
| Psychiatric clinic | 5 | 1.4 | 5 | 1.9 | 0 | 0 | |
| Dermatovenerological clinic | 8 | 2.3 | 6 | 2.3 | 2 | 2.2 | |
| Urological outpatient clinic | 10 | 2.8 | 5 | 1.9 | 5 | 5.5 | |
| Institute of Pathological Anatomy | 3 | 0.8 | 0 | 0 | 3 | 3.3 | |
| No. 2 | Geriatric clinic | 76 | 21.5 | 58 | 22.1 | 18 | 19.8 |
| Long-term care department | 18 | 5.1 | 14 | 5.3 | 4 | 4.4 | |
| Aftercare department | 12 | 3.4 | 11 | 4.2 | 1 | 1.1 | |
| Other outpatient offices | 5 | 1.4 | 3 | 1.1 | 2 | 2.2 | |
| Total of (260) | A/B (20) | C0 (17) | C1 (3) | C2 (220) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gender | n | % | n | % | n | % | n | % | n | % | |
| Male | 84 | 32.3 | 10 | 50 | 4 | 23.5 | 2 | 66.7 | 68 | 30.9 | |
| Female | 176 | 67.7 | 10 | 50 | 13 | 76.5 | 1 | 33.3 | 152 | 69.1 | |
| Age | |||||||||||
| ≥65 years | 235 | 90.4 | 16 | 80 | 16 | 94.1 | 3 | 100 | 200 | 90.9 | |
| ˂65 years | 25 | 9.6 | 4 | 20 | 1 | 5.9 | 20 | 9.1 | |||
| Setting | |||||||||||
| No. 1 | First internal clinic | 140 | 53.8 | 12 | 60 | 12 | 70.6 | 116 | 52.7 | ||
| Surgical clinic | 13 | 5 | 1 | 5 | 12 | 5.4 | |||||
| Neurological clinic | 6 | 2.3 | 6 | 2.7 | |||||||
| Psychiatric clinic | 5 | 1.9 | 5 | 2.3 | |||||||
| Dermatovenerological clinic | 6 | 2.3 | 6 | 2.7 | |||||||
| Urological outpatient clinic | 5 | 1.9 | 2 | 10 | 3 | 1.4 | |||||
| Institute of Pathological Anatomy | |||||||||||
| No. 2 | Geriatric clinic | 58 | 22.3 | 4 | 20 | 3 | 17.6 | 2 | 66.7 | 49 | 22.3 |
| Long-term care department | 14 | 5.4 | 2 | 11.8 | 12 | 5.5 | |||||
| Aftercare department | 10 | 3.8 | 1 | 5 | 1 | 33.3 | 8 | 3.6 | |||
| Other outpatient offices | 3 | 1.2 | 3 | 1.4 | |||||||
| Antibiotics | Total | ST131 Strains | Non-ST131 Strains | p * | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | R | (%) | n | R | (%) | n | R | (%) | ||
| Ampicillin/sulbactam | 321 | 245 | 76.3 | 244 | 193 | 79.1 | 77 | 52 | 67.5 | 0.037 |
| Piperacillin/tazobactam | 317 | 60 | 18.9 | 241 | 49 | 20.3 | 76 | 11 | 14.5 | 0.002 |
| Cefuroxime | 320 | 317 | 99.1 | 243 | 242 | 99.6 | 77 | 75 | 97.4 | |
| Cefotaxime | 319 | 314 | 98.4 | 243 | 241 | 99.2 | 76 | 73 | 96.1 | 0.043 |
| Ceftazidime | 320 | 251 | 78.4 | 244 | 199 | 81.6 | 76 | 52 | 68.4 | 0.042 |
| Ceftazidime/avibactam | 347 | 0 | 0 | 262 | 0 | 0 | 85 | 0 | 0 | |
| Cefoperazone/sulbactam | 280 | 18 | 6.4 | 214 | 15 | 7 | 66 | 3 | 4.5 | |
| Cefepime | 281 | 234 | 83.3 | 214 | 182 | 85 | 67 | 52 | 77.6 | |
| Ertapenem | 280 | 3 | 1.1 | 214 | 1 | 0.5 | 66 | 2 | 3 | |
| Meropenem | 319 | 0 | 0 | 243 | 0 | 0 | 76 | 0 | 0 | |
| Gentamicin | 319 | 78 | 24.5 | 242 | 54 | 22.3 | 77 | 24 | 31.2 | |
| Tobramycin | 280 | 154 | 55 | 214 | 121 | 56.5 | 66 | 33 | 50 | |
| Amikacin | 320 | 14 | 4.4 | 244 | 10 | 4.1 | 76 | 4 | 5.3 | 0.038 |
| Tetracycline | 299 | 190 | 63.5 | 226 | 145 | 64.2 | 73 | 45 | 61.6 | |
| Tigecycline | 280 | 3 | 1.1 | 214 | 3 | 1.4 | 66 | 0 | 0 | |
| Ciprofloxacin | 321 | 301 | 93.8 | 244 | 236 | 96.7 | 77 | 65 | 84.4 | <0.001 |
| Colistin | 276 | 1 | 0.4 | 211 | 1 | 0.5 | 65 | 0 | 0 | |
| Trimethoprim/sulfamethoxazole | 319 | 241 | 75.5 | 242 | 186 | 76.9 | 77 | 55 | 71.4 | |
| Nitrofurantoin | 72 | 1 | 1.4 | 51 | 1 | 2 | 21 | 0 | 0 | |
| Fosfomycin | 69 | 2 | 2.9 | 49 | 1 | 2 | 20 | 1 | 5 | |
| Primer | Sequence | Length of Product |
|---|---|---|
| ST-131_F | AGCAACGATATTTGCCCATT | 580 bp |
| ST-131_R | GGCGATAACAGTACGCCATT | |
| Clade C1_F | TGAATCAAAGGTCCGAGCTG | 232 bp |
| Clade C1_R | TATGGCTGGCAGATGCTTTA | |
| Clade C2_F | ACGGATTCAGGTAGACGATT | 164 bp |
| Clade C2_R | CCTCACCAAAGTTGCGATTAC | |
| FimH-30_F | CCGCCAATGGTACCGCTATT | 354 bp |
| FimH-30_R | CAGCTTTAATCGCCACCCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koreň, J.; Andrezál, M.; Ozaee, E.; Drahovská, H.; Wawruch, M.; Liptáková, A.; Maliar, T. High Emergence of Multidrug-Resistant Sequence Type 131 Subclade C2 among Extended-Spectrum β-Lactamase (ESBL)-Producing Escherichia coli Isolated from the University Hospital Bratislava, Slovakia. Antibiotics 2023, 12, 1209. https://doi.org/10.3390/antibiotics12071209
Koreň J, Andrezál M, Ozaee E, Drahovská H, Wawruch M, Liptáková A, Maliar T. High Emergence of Multidrug-Resistant Sequence Type 131 Subclade C2 among Extended-Spectrum β-Lactamase (ESBL)-Producing Escherichia coli Isolated from the University Hospital Bratislava, Slovakia. Antibiotics. 2023; 12(7):1209. https://doi.org/10.3390/antibiotics12071209
Chicago/Turabian StyleKoreň, Ján, Michal Andrezál, Elham Ozaee, Hana Drahovská, Martin Wawruch, Adriána Liptáková, and Tibor Maliar. 2023. "High Emergence of Multidrug-Resistant Sequence Type 131 Subclade C2 among Extended-Spectrum β-Lactamase (ESBL)-Producing Escherichia coli Isolated from the University Hospital Bratislava, Slovakia" Antibiotics 12, no. 7: 1209. https://doi.org/10.3390/antibiotics12071209