
Serotyping and Antimicrobial Resistance Profiling of Multidrug-Resistant Non-Typhoidal Salmonella from Farm Animals in Hunan, China
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Salmonella Strains Isolation
2.2. Prevalence of Salmonella Serovar
2.3. Antibiotic Susceptibility Testing
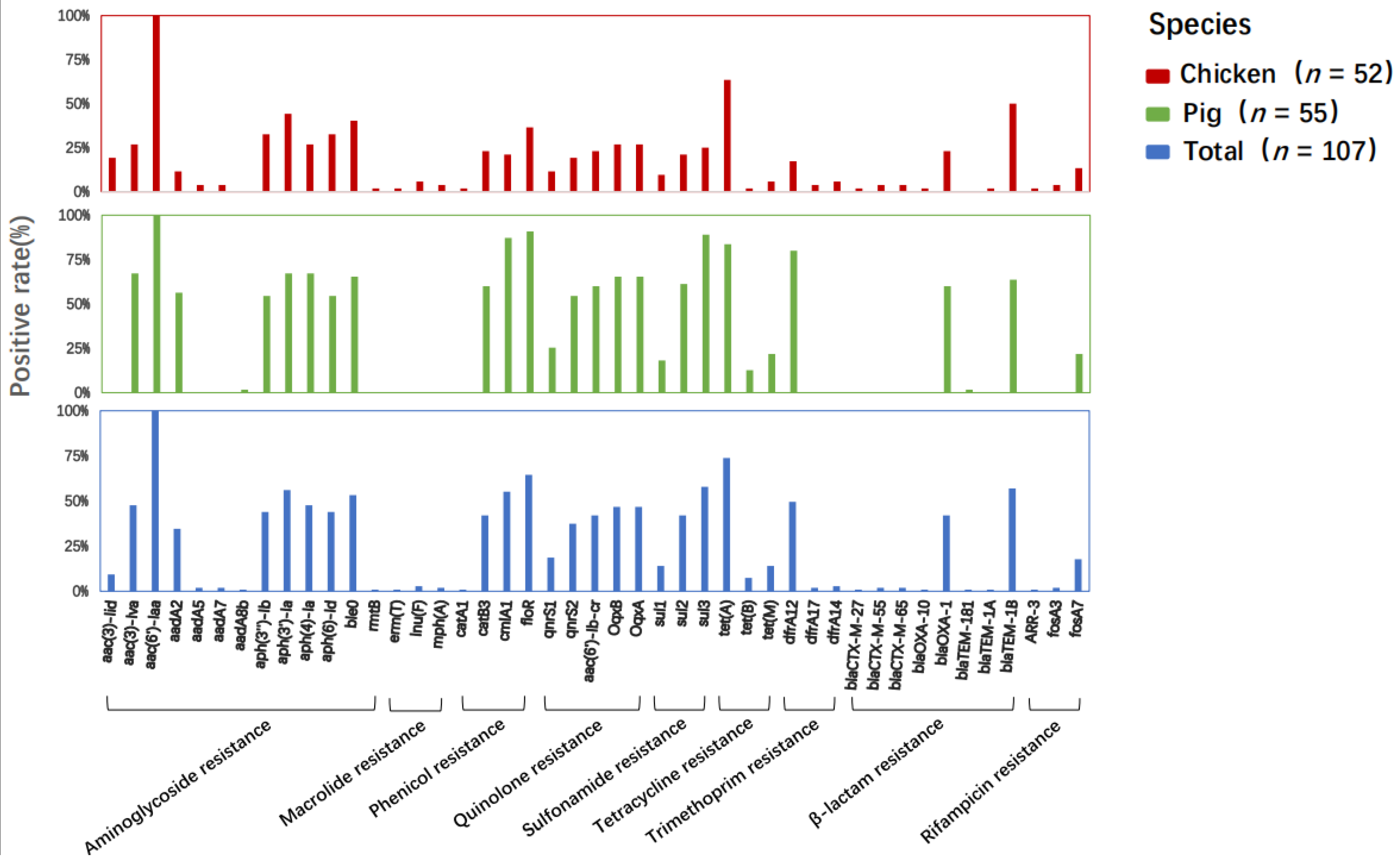
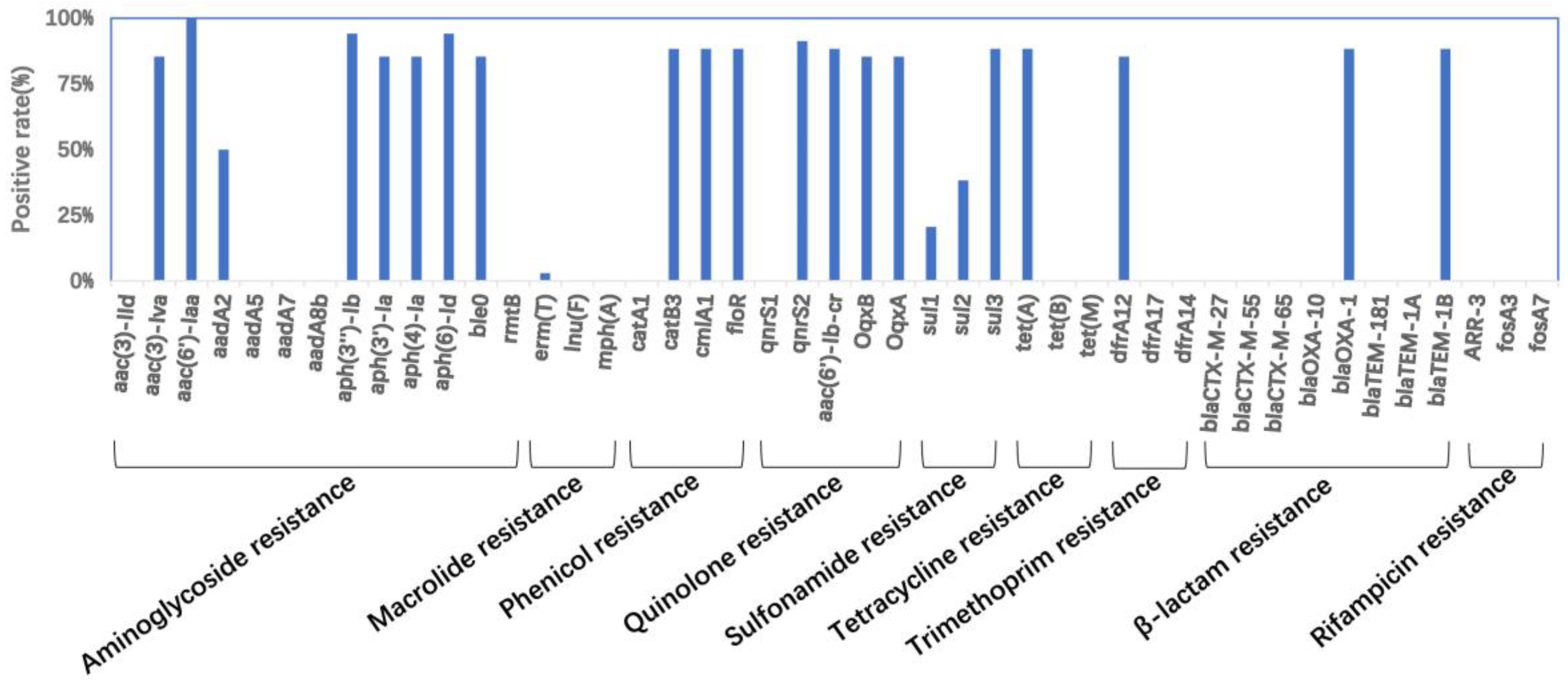
2.4. Antibiotic Resistance Gene and Plasmid Profiles
2.5. Characteristic of S. 4,[5],12:i:-
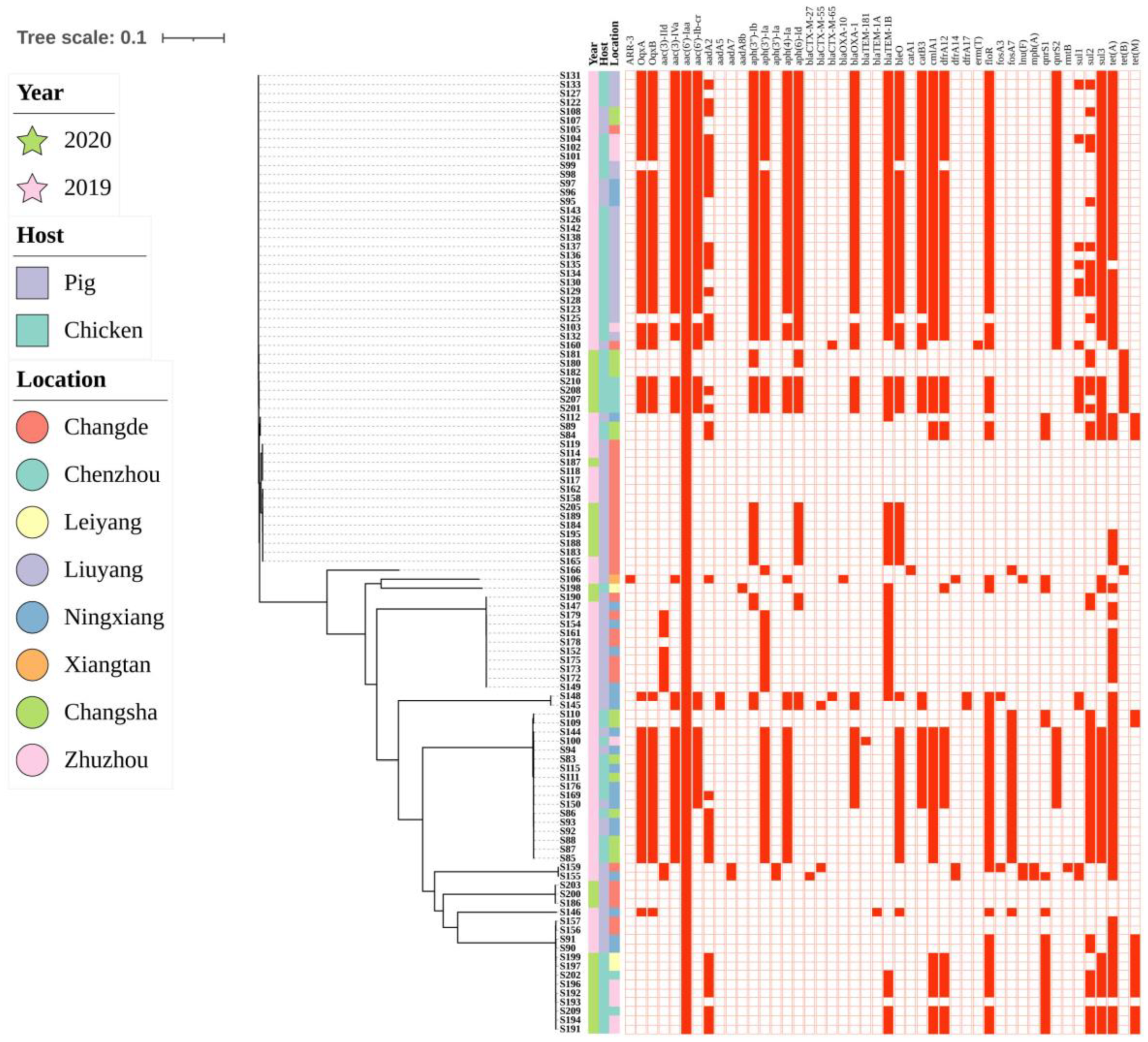
2.6. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Isolation of Salmonella Strains
4.2. Detection of the Salmonella Strain Serotypes
4.3. Antibiotic Susceptibility Testing
4.4. Whole-Genome Sequencing, De Novo Assembly, and Annotation
4.5. Antibiotic Resistance Genes and Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hendriksen, R.S.; Vieira, A.R.; Karlsmose, S.; Lo Fo Wong, D.M.; Jensen, A.B.; Wegener, H.C.; Aarestrup, F.M. Global monitoring of Salmonella serovar distribution from the World Health Organization Global Foodborne Infections Network Country Data Bank: Results of quality assured laboratories from 2001 to 2007. Foodborne Pathog. Dis. 2011, 8, 887–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. National Enteric Disease Surveillance: Salmonella Annual Report. 2016. Available online: https://www.census.gov/geo/pdfs/maps-data/maps/ (accessed on 24 April 2021).
- EFSA; ECDC. European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC)The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Wu, B.; Pan, H.; Paudyal, N.; Jiang, J.; Zhang, L.; Li, Y.; Yue, M. ONE Health Approach to Address Zoonotic Brucellosis: A Spatiotemporal Associations Study Between Animals and Humans. Front. Vet. Sci. 2020, 7, 521. [Google Scholar] [CrossRef]
- Arnold, M.E.; Papadopoulou, C.; Davies, R.H.; Carrique-Mas, J.J.; Evans, S.J.; Hoinville, L.J. Estimation of Salmonella prevalence in UK egg-laying holdings. Prev. Vet. Med. 2010, 94, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Dopfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhou, X.; Jiang, Z.; Qi, Y.; Ed-Dra, A.; Yue, M. Epidemiological Investigation and Antimicrobial Resistance Profiles of Salmonella Isolated From Breeder Chicken Hatcheries in Henan, China. Front. Cell. Infect. Microbiol. 2020, 10, 497. [Google Scholar] [CrossRef]
- Bonardi, S. Salmonella in the pork production chain and its impact on human health in the European Union. Epidemiol. Infect. 2017, 145, 1513–1526. [Google Scholar] [CrossRef] [Green Version]
- WHO Guidelines Approved by the Guidelines Review Committee. In WHO Guidelines on Use of Medically Important Antimicrobials in Food-Producing Animals; World Health Organization: Geneva, Switzerland, 2017.
- Cuypers, W.L.; Jacobs, J.; Wong, V.; Klemm, E.J.; Deborggraeve, S.; Van Puyvelde, S. Fluoroquinolone resistance in Salmonella: Insights by whole-genome sequencing. Microb. Genom. 2018, 4, e000195. [Google Scholar] [CrossRef]
- Ito, J.; Sugimoto, R.; Nakaoka, H.; Yamada, S.; Kimura, T.; Hayano, T.; Inoue, I. Systematic identification and characterization of regulatory elements derived from human endogenous retroviruses. PLoS Genet. 2017, 13, e1006883. [Google Scholar] [CrossRef] [Green Version]
- Campos, J.; Mourao, J.; Peixe, L.; Antunes, P. Non-typhoidal Salmonella in the Pig Production Chain: A Comprehensive Analysis of Its Impact on Human Health. Pathogens 2019, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Wan, Y.; Du, P.; Bai, L. The Epidemiology of Monophasic Salmonella Typhimurium. Foodborne Pathog. Dis. 2020, 17, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kang, X.; Ed-Dra, A.; Zhou, X.; Jia, C.; Müller, A.; Liu, Y.; Kehrenberg, C.; Yue, M. Genome-Based Assessment of Antimicrobial Resistance and Virulence Potential of Isolates of Non-Pullorum/Gallinarum Salmonella Serovars Recovered from Dead Poultry in China. Microbiol. Spectr. 2022, 10, e0096522. [Google Scholar] [CrossRef] [PubMed]
- China Agriculture Statistical Report; Ministry of Agriculture and Rural Affairs: Beijing, China, 2021.
- Vico, J.P.; Lorenzutti, A.M.; Zogbi, A.P.; Aleu, G.; Sanchez, I.C.; Caffer, M.I.; Rosmini, M.R.; Mainar-Jaime, R.C. Prevalence, associated risk factors, and antimicrobial resistance profiles of non-typhoidal Salmonella in large scale swine production in Cordoba, Argentina. Res. Vet. Sci. 2020, 130, 161–169. [Google Scholar] [CrossRef]
- Sia, C.M.; Baines, S.L.; Valcanis, M.; Lee, D.Y.J.; Gonçalves da Silva, A.; Ballard, S.A.; Easton, M.; Seemann, T.; Howden, B.P.; Ingle, D.J.; et al. Genomic diversity of antimicrobial resistance in non-typhoidal Salmonella in Victoria, Australia. Microb. Genom. 2021, 7, 000725. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, J.; Ed-Dra, A.; Li, X.; Peng, X.; Xia, L.; Guo, Q.; Yao, G.; Yue, M. Prevalence and genomic investigation of Salmonella isolates recovered from animal food-chain in Xinjiang, China. Food. Res. Int. 2021, 142, 110198. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Chen, W.; Elbediwi, M.; Pan, H.; Wang, L.; Zhou, C.; Zhao, B.; Xu, X.; Li, D.; Yan, X.; et al. Characterization of Salmonella Resistome and Plasmidome in Pork Production System in Jiangsu, China. Front. Vet. Sci. 2020, 7, 617. [Google Scholar] [CrossRef]
- Jiang, Z.; Paudyal, N.; Xu, Y.; Deng, T.; Li, F.; Pan, H.; Peng, X.; He, Q.; Yue, M. Antibiotic Resistance Profiles of Salmonella Recovered From Finishing Pigs and Slaughter Facilities in Henan, China. Front. Microbiol. 2019, 10, 1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Song, J.; Zeng, X.; Chen, D.; Chen, R.; Qiu, C.; Zhou, K. National Prevalence of Salmonella enterica Serotype Kentucky ST198 with High-Level Resistance to Ciprofloxacin and Extended-Spectrum Cephalosporins in China, 2013 to 2017. mSystems 2021, 6, e00935-20. [Google Scholar] [CrossRef]
- Elbediwi, M.; Li, Y.; Paudyal, N.; Pan, H.; Li, X.; Xie, S.; Rajkovic, A.; Feng, Y.; Fang, W.; Rankin, S.C.; et al. Global Burden of Colistin-Resistant Bacteria: Mobilized Colistin Resistance Genes Study (1980–2018). Microorganisms 2019, 7, 461. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Sun, F.; Sun, D.; Wang, Z.; Jin, S.; Pan, Z.; Xu, Z.; Chen, X.; Jiao, X. Multidrug resistance and prevalence of quinolone resistance genes of Salmonella enterica serotypes 4,[5],12:i:- in China. Int. J. Food Microbiol. 2020, 330, 108692. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, X.; Kuang, D.; Shi, X.; Xiao, W.; Zhang, J.; Gu, Z.; Xu, X.; Meng, J. Prevalence of antimicrobial resistance of non-typhoidal Salmonella serovars in retail aquaculture products. Int. J. Food Microbiol. 2015, 210, 47–52. [Google Scholar] [CrossRef]
- Hyeon, J.Y.; Li, S.; Mann, D.A.; Zhang, S.; Kim, K.J.; Lee, D.H.; Deng, X.; Song, C.S. Whole-Genome Sequencing Analysis of Salmonella Enterica Serotype Enteritidis Isolated from Poultry Sources in South Korea, 2010–2017. Pathogens 2021, 10, 45. [Google Scholar] [CrossRef]
- Vidovic, S.; An, R.; Rendahl, A. Molecular and Physiological Characterization of Fluoroquinolone-Highly Resistant Salmonella Enteritidis Strains. Front. Microbiol. 2019, 10, 729. [Google Scholar] [CrossRef]
- Hauser, E.; Tietze, E.; Helmuth, R.; Junker, E.; Blank, K.; Prager, R.; Rabsch, W.; Appel, B.; Fruth, A.; Malorny, B. Pork contaminated with Salmonella enterica serovar 4,[5],12:i:-, an emerging health risk for humans. Appl. Environ. Microbiol. 2010, 76, 4601–4610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbediwi, M.; Pan, H.; Biswas, S.; Li, Y.; Yue, M. Emerging colistin resistance in Salmonella enterica serovar Newport isolates from human infections. Emerg. Microbes. Infect. 2020, 9, 535–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaconu, E.L.; Alba, P.; Feltrin, F.; Di Matteo, P.; Iurescia, M.; Chelli, E.; Donati, V.; Marani, I.; Giacomi, A.; Franco, A.; et al. Emergence of IncHI2 Plasmids With Mobilized Colistin Resistance (mcr)-9 Gene in ESBL-Producing, Multidrug-Resistant Salmonella Typhimurium and Its Monophasic Variant ST34 From Food-Producing Animals in Italy. Front. Microbiol. 2021, 12, 705230. [Google Scholar] [CrossRef]
- Neuert, S.; Nair, S.; Day, M.R.; Doumith, M.; Ashton, P.M.; Mellor, K.C.; Jenkins, C.; Hopkins, K.L.; Woodford, N.; de Pinna, E.; et al. Prediction of Phenotypic Antimicrobial Resistance Profiles From Whole Genome Sequences of Non-typhoidal Salmonella enterica. Front. Microbiol. 2018, 9, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Yue, M.; Rankin, S.; Weill, F.X.; Frey, J.; Schifferli, D.M. One-Step Identification of Five Prominent Chicken Salmonella Serovars and Biotypes. J. Clin. Microbiol. 2015, 53, 3881–3883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guibourdenche, M.; Roggentin, P.; Mikoleit, M.; Fields, P.I.; Bockemuhl, J.; Grimont, P.A.; Weill, F.X. Supplement 2003–2007 (No. 47) to the White-Kauffmann-Le Minor scheme. Res. Microbiol. 2010, 161, 26–29. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th Edn.CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Yang, J.; Chen, W.; Liu, Z.; Zhao, Q.; Yang, H.; Sun, Z.; Chen, X.; Li, J. Prevalence and Characteristics of mcr-1-Producing Escherichia coli in Three Kinds of Poultry in Changsha, China. Front. Microbiol. 2022, 13, 840520. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antibiotic Agent | 2019 (n = 79) | 2020 (n = 28) |
|---|---|---|
| Penicillin: Ampicillin | 56 (70.9%) | 20 (71.4%) |
| β-lactams combination: Amoxicillin/Clavulanic | 3 (3.9%) | 2 (7.1%) |
| Aminoglycosides: Gentamicin | 36 (45.6%) | 2 (7.1%) |
| Spectinomycin | 23 (29.1%) | 13 (46.4%) |
| Tetracyclines: Tetracycline | 27 (34.2%) | 21 (75.0%) |
| Florfenicol | 59 (74.7%) | 13 (46.4%) |
| Sulfafurazole | 52 (65.8%) | 16 (57.1%) |
| Trimethoprim/sulfamethoxazole | 47 (59.5%) | 13 (46.4%) |
| Ceftiofur | 9 (11.4%) | - |
| Ceftazidime | 2 (2.5%) | - |
| Enrofloxacin | 43 (54.4%) | 4 (14.3%) |
| Ofloxacin | 17 (21.5%) | - |
| Meropenem | 2 (2.5%) | - |
| Apramycin | - | - |
| Polymyxins: Colistin | 5 (6.3%) | - |
| Mequindox | - |
| Serotypes | In Total | Pig | Poultry |
|---|---|---|---|
| Potential monophasic variant of Typhimurium | 34 | 27 | 7 |
| Typhimurium | 20 | 6 | 15 |
| Derby | 17 | 12 | 5 |
| Rissen | 13 | 9 | 4 |
| Enteritidis | 11 | - | 11 |
| Apeyeme | 3 | - | 3 |
| Kentucky | 2 | - | 2 |
| Indiana | 2 | - | 2 |
| London | 1 | 1 | - |
| Meleagridis | 1 | - | 1 |
| Thompson | 1 | - | 1 |
| Reading | 1 | - | 1 |
| In total | 107 | 55 | 52 |
| Antibiotic Agent | Abbreviation | Antibiotic Concentration Range (μg/mL) | Breakpoint Interpretive Criteria (μg/mL) | Results in Percentage (%) | ||||
|---|---|---|---|---|---|---|---|---|
| S | I | R | S | I | R | |||
| Penicillin: Ampicillin | AMP | 0–512 | ≤8 | 16 | ≥32 | 32 (29.9%) | 0 | 75 (70%) |
| β-lactams combination: Amoxicillin/Clavulanic | AMC | 0.5/0.25–256/128 | ≤8/4 | 16/8 | ≥32/16 | 76 (71%) | 26 (24.3%) | 5 (4.7%) |
| Aminoglycosides: Gentamicin | GEN | 0.25–128 | ≤4 | 8 | ≥16 | 51 (47.7%) | 19 (17.8%) | 37 (34.6%) |
| Spectinomycin | STP | 0–512 | ≤32 | 64 | ≥128 | 14 (13%) | 26 (24%) | 67 (62.6%) |
| Tetracyclines: Tetracycline | TET | 0–512 | ≤4 | 8 | ≥16 | 17 (15.9%) | 0 | 90 (84.1%) |
| Florfenicol | FFC | 0–256 | ≤4 | 8 | ≥16 | 36 (33.6%) | 0 | 71 (66.4%) |
| Sulfafurazole | SOX | 0–512 | ≤256 | - | ≥512 | 39 (36.5%) | - | 68 (63.6%) |
| Co-trimoxazole | SXT | 0–32/608 | ≤2/38 | - | ≥4/76 | 47 (43.9%) | - | 60 (56%) |
| Ceftiofur | EFT | 0.12–256 | ≤2 | 4 | ≥8 | 96 (89.7%) | 1 (0.9%) | 10 (9.4%) |
| Ceftazidime | CAZ | 0.12–256 | ≤4 | 8 | ≥16 | 97 (90.7%) | 4 (3.7%) | 6 (5.6%) |
| Enrofloxacin | ENR | 0.01–32 | ≤0.25 | 0.5–1 | ≥2 | 18 (16.8%) | 42 (39.3%) | 47 (43.9%) |
| Ofloxacin | OFX | 0.03–64 | ≤2 | 4 | ≥8 | 57 (53.3%) | 33 (30.8%) | 17 (15.9%) |
| Meropenem | MEM | 0.03–10 | ≤1 | 2 | ≥4 | 104 (97.2%) | 0 | 3 (2.8%) |
| Apramycin | APR | 0–64 | - | - | - | - | - | - |
| Polymyxins: Colistin | CL | 0.12–256 | ≤2 | - | ≥4 | 95 (88.8%) | - | 12 (11.2%) |
| Mequindox | NA | 1–512 | - | - | - | - | - | - |
| Antimicrobial Agents | Pig (n = 55) | Chicken (n = 52) | p-Value |
|---|---|---|---|
| Ampicillin | 40 (72.7%) | 35 (67.3%) | 0.6903 |
| Amoxicillin/Clavulanic | 2 (3.6%) | 3 (5.8%) | 0.6013 |
| Gentamicin | 17 (30.9%) | 21 (40.4%) | 0.3059 |
| Spectinomycin | 50 (90.9%) | 18 (34.6%) | 1.47 |
| Tetracycline | 54 (98.2%) | 37 (71.2%) | <0.0005 |
| Florfenicol | 51 (92.7%) | 21 (40.4%) | <0.0005 |
| Sulfafurazole | 50 (90.9%) | 19 (36.5%) | <0.0005 |
| Trimethoprim/sulfamethoxazole | 46 (83.6%) | 14 (26.9%) | <0.0005 |
| Ceftiofur | 1 (1.8%) | 8 (15.4%) | 0.0115 |
| Ceftazidime | 0 (0%) | 2 (3.9%) | 0.1420 |
| Enrofloxacin | 33 (60.0%) | 14 (26.9%) | 0.0005 |
| Ofloxacin | 7 (12.7%) | 10 (19.2%) | 0.3576 |
| Meropenem | 0 (0%) | 2 (3.9%) | 0.1420 |
| Apramycin | - | - | |
| Colistin | 0 (0%) | 11 (21.2%) | 0.0003 |
| Mequindox | - | - |
| Numbers | 4,[5],12:i:- (n = 34) | |
|---|---|---|
| Antibiotic agent | Drug resistant | Multidrug resistant (n = 31) |
| Tetracyclines: Tetracycline | 34 (100%) | 31 |
| β-lactam: Ampicillin | 31 (91.2%) | 31 |
| Chloramphenicols: Florfenicol | 31 (91.2%) | 31 |
| Aminocyclitols: Spectinomycin | 31 (91.2%) | 31 |
| Sulfonamides: trimethoprim/sulfamethoxazole | 30 (88.2%) | 30 |
| Fluoroquinolones: Enrofloxacin | 26 (76.5%) | 26 |
| Sulfonamides: Sulfafurazole | 24 (70.6%) | 22 |
| Aminoglycosides: Gentamicin | 16 (47.1%) | 16 |
| Fluoroquinolones: Ofloxacin | 7 (20.6%) | 7 |
| Cephalosporins: Ceftiofur | 1 (2.9%) | 1 |
| Polymyxins: Colistin | 1(2.9%) | 1 |
| β-lactams combination: Amoxicillin/Clavulanic | - | - |
| Carbapenems: Meropenem | - | - |
| Cephalosporins: Ceftazidime | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Li, J.; Zhou, R.; Xu, Q.; Qu, S.; Lin, H.; Wang, Y.; Li, P.; Zheng, X. Serotyping and Antimicrobial Resistance Profiling of Multidrug-Resistant Non-Typhoidal Salmonella from Farm Animals in Hunan, China. Antibiotics 2023, 12, 1178. https://doi.org/10.3390/antibiotics12071178
Zhang Z, Li J, Zhou R, Xu Q, Qu S, Lin H, Wang Y, Li P, Zheng X. Serotyping and Antimicrobial Resistance Profiling of Multidrug-Resistant Non-Typhoidal Salmonella from Farm Animals in Hunan, China. Antibiotics. 2023; 12(7):1178. https://doi.org/10.3390/antibiotics12071178
Chicago/Turabian StyleZhang, Zhuohui, Jiyun Li, Rushun Zhou, Qianqian Xu, Shiyin Qu, Hongguang Lin, Yan Wang, Pishun Li, and Xiaofeng Zheng. 2023. "Serotyping and Antimicrobial Resistance Profiling of Multidrug-Resistant Non-Typhoidal Salmonella from Farm Animals in Hunan, China" Antibiotics 12, no. 7: 1178. https://doi.org/10.3390/antibiotics12071178