
Effects of Monensin and Rapamycin Combination Therapy on Tumor Growth and Apoptosis in a Xenograft Mouse Model of Neuroblastoma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
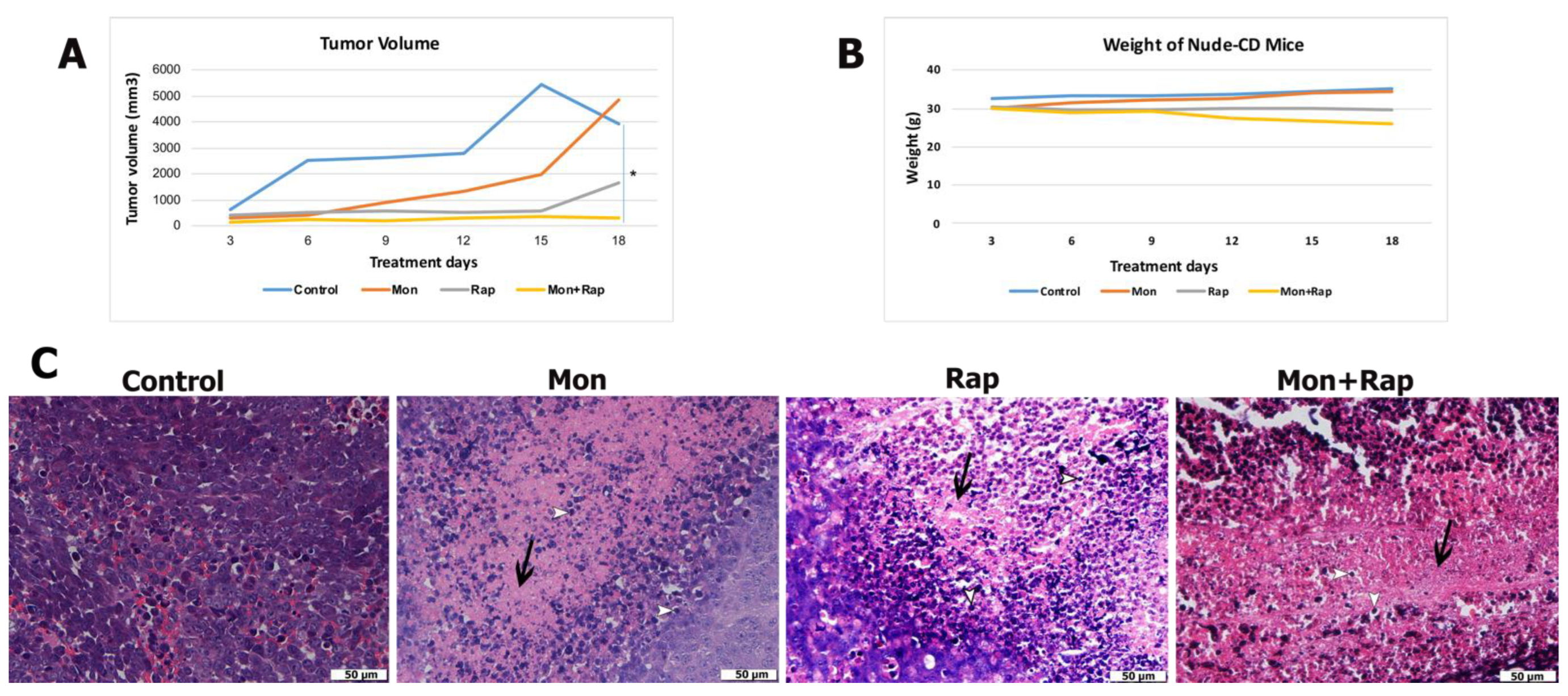
2.1. Combination Treatment of Monensin and Rapamycin Reduces Xenograft Neuroblastoma Tumor Development
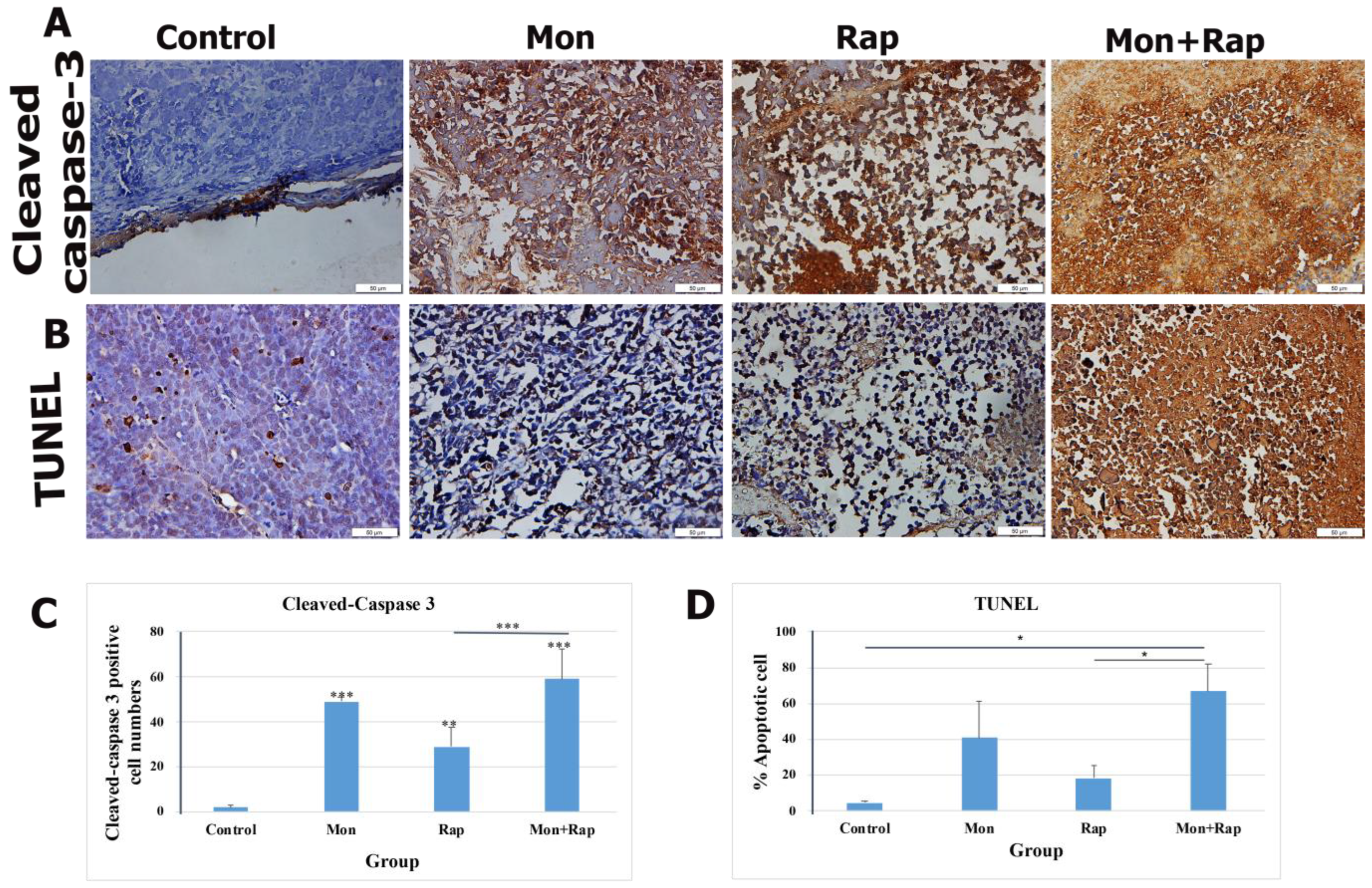
2.2. Combination Treatment of Monensin and Rapamycin Triggers Apoptosis in Xenograft Neuroblastoma Tumor Tissues
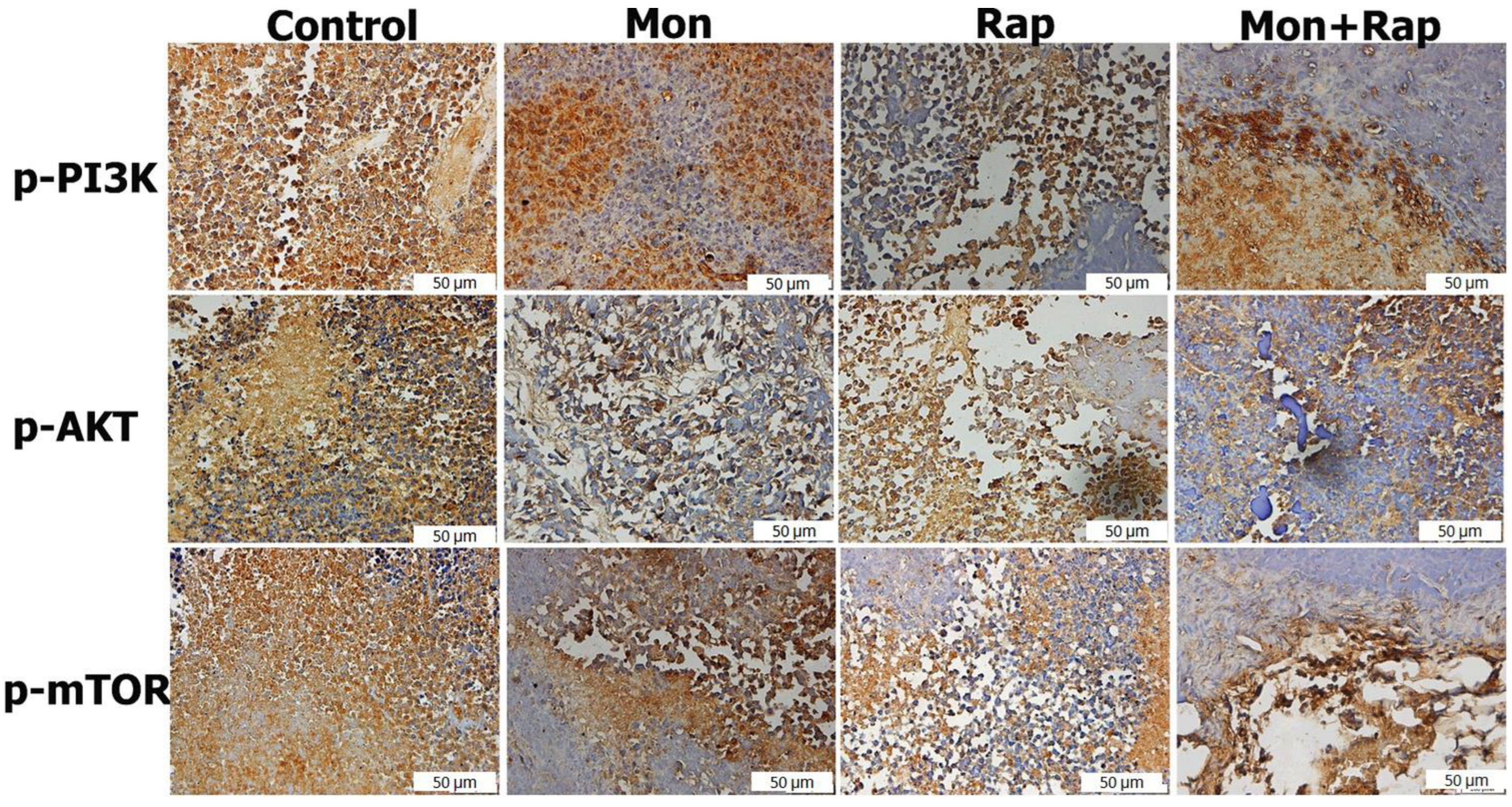
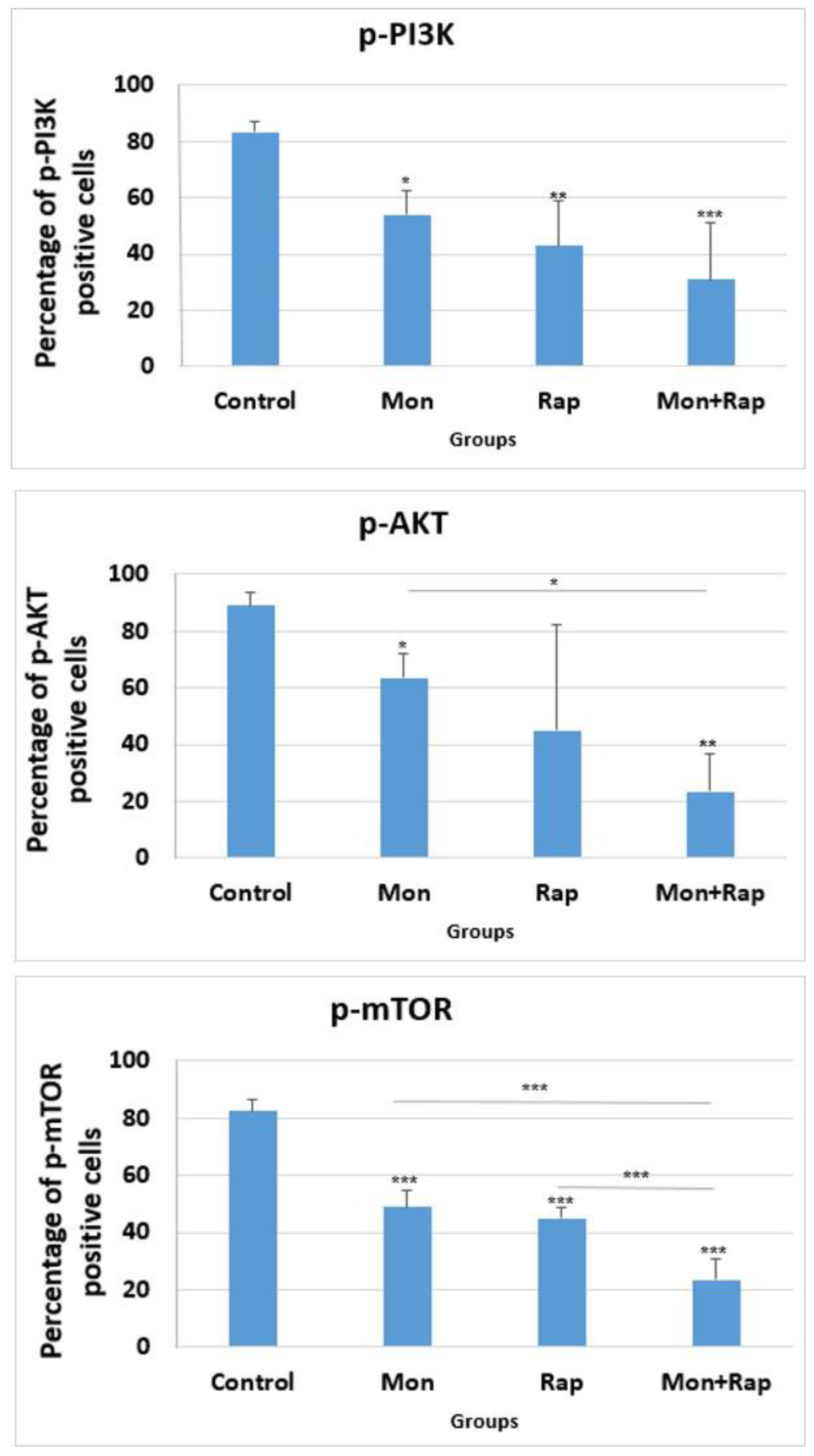
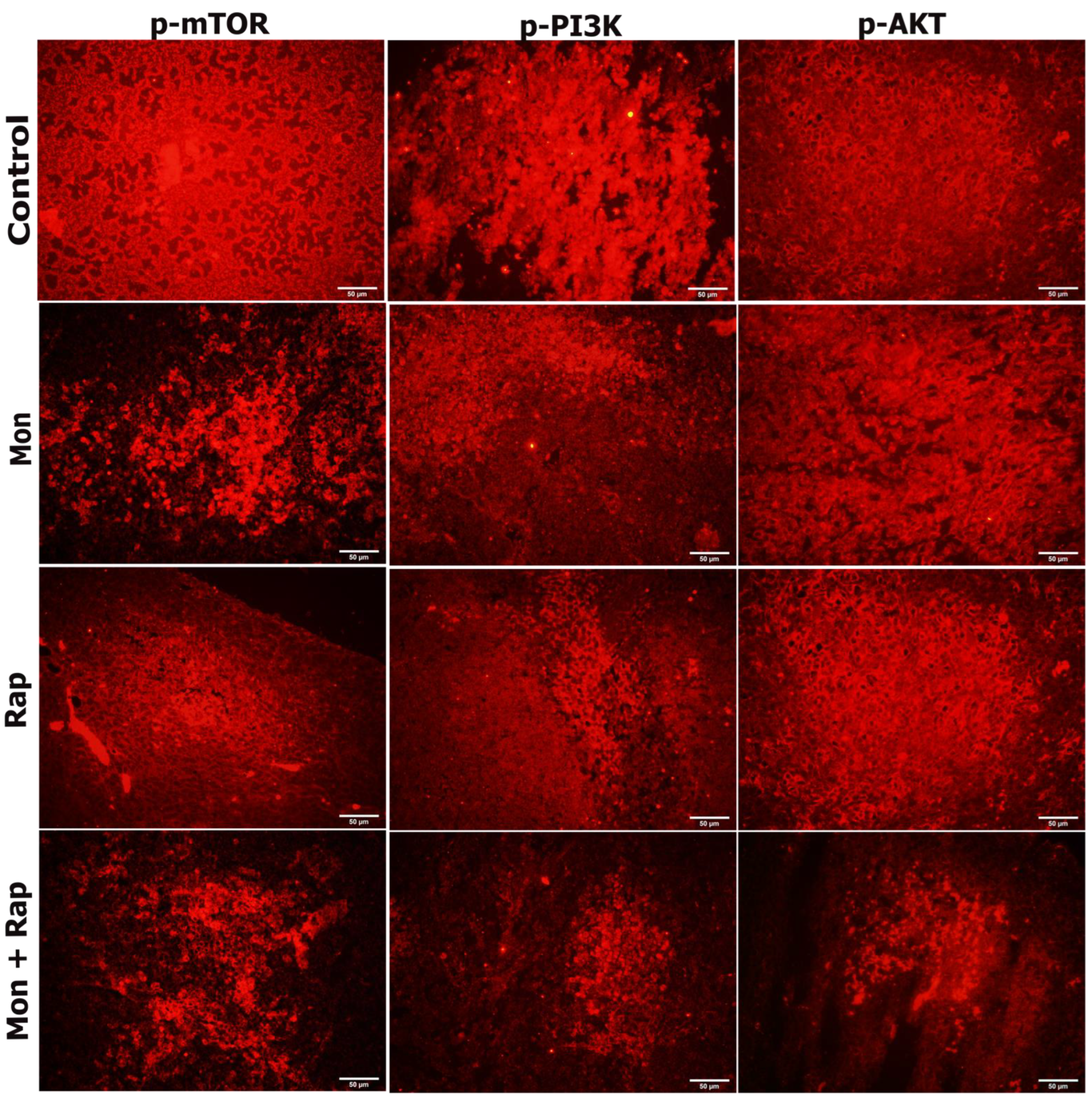
2.3. The Combination of Monensin and Rapamycin Reduces PI3K/AKT/mTOR Expression in Xenograft Neuroblastoma Tumor Tissues
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals
4.3. Establishment of a Xenograft Neuroblastoma Cancer Model
4.4. Formation of Experimental Groups
4.5. Histological Evaluation of Xenograft Neuroblastoma Tumor Tissues
4.6. Determination of Apoptosis by TUNEL Method in Xenograft Neuroblastoma Tumor Tissues
4.7. Evaluation of Cleaved-Caspase 3 and p-PI3K/p-AKT/p-mTOR Protein Expression by Immunohistochemistry in Xenograft Neuroblastoma Tumor Tissues
4.8. Evaluation of Cleaved-Caspase 3 and p-PI3K/p-AKT/p-mTOR Protein Expression by Immunofluorescence in Xenograft Neuroblastoma Tumor Tissues
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Whittle, S.B.; Smith, V.; Doherty, E.; Zhao, S.; McCarty, S.; Zage, P.E. Overview and Recent Advances in the Treatment of Neuroblastoma. Expert Rev. Anticancer Ther. 2017, 17, 369–386. [Google Scholar] [CrossRef] [PubMed]
- Aaltonen, K.; Radke, K.; Adamska, A.; Seger, A.; Mañas, A.; Bexell, D. Patient-Derived Models: Advanced Tools for Precision Medicine in Neuroblastoma. Front. Oncol. 2022, 12, 1085270. [Google Scholar] [CrossRef] [PubMed]
- Colon, N.C.; Chung, D.H. Neuroblastoma. Adv. Pediatr. 2011, 58, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.C.; Clark, R.A.; Chung, D.H. High-Risk Neuroblastoma: A Surgical Perspective. Children 2023, 10, 388. [Google Scholar] [CrossRef]
- Alexander, F. Neuroblastoma. Urol. Clin. N. Am. 2000, 27, 383–392. [Google Scholar] [CrossRef]
- Ishola, T.A.; Chung, D.H. Neuroblastoma. Surg. Oncol. 2007, 16, 149–156. [Google Scholar] [CrossRef]
- Markowska, A.; Kaysiewicz, J.; Markowska, J.; Huczyński, A. Doxycycline, Salinomycin, Monensin and Ivermectin Repositioned as Cancer Drugs. Bioorg. Med. Chem. Lett. 2019, 29, 1549–1554. [Google Scholar] [CrossRef]
- Huczyński, A.; Ratajczak-Sitarz, M.; Stefańska, J.; Katrusiak, A.; Brzezinski, B.; Bartl, F. Reinvestigation of the Structure of Monensin A Phenylurethane Sodium Salt Based on X-Ray Crystallographic and Spectroscopic Studies, and Its Activity against Hospital Strains of Methicillin-Resistant S. Epidermidis and S. Aureus. J. Antibiot. 2011, 64, 249–256. [Google Scholar] [CrossRef]
- Wang, X.; Wu, X.; Zhang, Z.; Ma, C.; Wu, T.; Tang, S.; Zeng, Z.; Huang, S.; Gong, C.; Yuan, C.; et al. Monensin Inhibits Cell Proliferation and Tumor Growth of Chemo-Resistant Pancreatic Cancer Cells by Targeting the EGFR Signaling Pathway. Sci. Rep. 2018, 8, 17914. [Google Scholar] [CrossRef]
- Gu, J.; Huang, L.; Zhang, Y. Monensin Inhibits Proliferation, Migration, and Promotes Apoptosis of Breast Cancer Cells via Downregulating UBA2. Drug Dev. Res. 2020, 81, 745–753. [Google Scholar] [CrossRef]
- Yao, S.; Wang, W.; Zhou, B.; Cui, X.; Yang, H.; Zhang, S. Monensin Suppresses Cell Proliferation and Invasion in Ovarian Cancer by Enhancing MEK1 SUMOylation. Exp. Ther. Med. 2021, 22, 1390. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, K.Y.; Yu, S.N.; Park, S.G.; Yu, H.S.; Seo, Y.K.; Ahn, S.C. Monensin Induces PC-3 Prostate Cancer Cell Apoptosis via ROS Production and Ca2+ Homeostasis Disruption. Anticancer Res. 2016, 36, 5835–5843. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One Drug, Many Effects. Cell Metab. 2014, 19, 373. [Google Scholar] [CrossRef]
- Benjamin, D.; Colombi, M.; Moroni, C.; Hall, M.N. Rapamycin Passes the Torch: A New Generation of MTOR Inhibitors. Nat. Rev. Drug Discov. 2011, 10, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Meric-Bernstam, F.; Esteva, F.J. Potential Role of Mammalian Target of Rapamycin Inhibitors in Breast Cancer Therapy. Clin. Breast Cancer 2005, 6, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Bu, L.M.; Wu, K.; Lu, L.G.; Wang, X.P. Rapamycin Inhibits the Proliferation of SW1990 Pancreatic Cancer Cell. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3072–3079. [Google Scholar]
- Raymond, E.; Alexandre, J.; Faivre, S.; Vera, K.; Materman, E.; Boni, J.; Leister, C.; Korth-Bradley, J.; Hanauske, A.; Armand, J.P. Safety and Pharmacokinetics of Escalated Doses of Weekly Intravenous Infusion of CCI-779, a Novel MTOR Inhibitor, in Patients with Cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2004, 22, 2336–2347. [Google Scholar] [CrossRef]
- Witzig, T.E.; Geyer, S.M.; Ghobrial, I.; Inwards, D.J.; Fonseca, R.; Kurtin, P.; Ansell, S.M.; Luyun, R.; Flynn, P.J.; Morton, R.F.; et al. Phase II Trial of Single-Agent Temsirolimus (CCI-779) for Relapsed Mantle Cell Lymphoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2005, 23, 5347–5356. [Google Scholar] [CrossRef]
- Mondesire, W.H.; Jian, W.; Zhang, H.; Ensor, J.; Hung, M.C.; Mills, G.B.; Meric-Bernstam, F. Targeting Mammalian Target of Rapamycin Synergistically Enhances Chemotherapy-Induced Cytotoxicity in Breast Cancer Cells. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 7031–7042. [Google Scholar] [CrossRef]
- Marimpietri, D.; Brignole, C.; Nico, B.; Pastorino, F.; Pezzolo, A.; Piccardi, F.; Cilli, M.; Di Paolo, D.; Pagnan, G.; Longo, L.; et al. Combined Therapeutic Effects of Vinblastine and Rapamycin on Human Neuroblastoma Growth, Apoptosis, and Angiogenesis. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 3977–3988. [Google Scholar] [CrossRef]
- Johnsen, J.I.; Segerström, L.; Orrego, A.; Elfman, L.; Henriksson, M.; Kågedal, B.; Eksborg, S.; Sveinbjörnsson, B.; Kogner, P. Inhibitors of Mammalian Target of Rapamycin Downregulate MYCN Protein Expression and Inhibit Neuroblastoma Growth in Vitro and in Vivo. Oncogene 2008, 27, 2910–2922. [Google Scholar] [CrossRef]
- Tumova, L.; Pombinho, A.R.; Vojtechova, M.; Stancikova, J.; Gradl, D.; Krausova, M.; Sloncova, E.; Horazna, M.; Kriz, V.; Machonova, O.; et al. Monensin Inhibits Canonical Wnt Signaling in Human Colorectal Cancer Cells and Suppresses Tumor Growth in Multiple Intestinal Neoplasia Mice. Mol. Cancer Ther. 2014, 13, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. Apoptosis Pathways and Neuroblastoma Therapy. Curr. Pharm. Des. 2009, 15, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Kocoglu, S.S.; Secme, M.; Oy, C.; Korkusuz, G.; Elmas, L. Monensin, an Antibiotic Isolated from Streptomyces Cinnamonensis, Regulates Human Neuroblastoma Cell Proliferation via the PI3K/AKT Signaling Pathway and Acts Synergistically with Rapamycin. Antibiotics 2023, 12, 546. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular Mechanisms of Cell Death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Han, L.; Weng, J.; Wang, K.; Chen, T. Rapamycin Inhibits Proliferation and Induces Autophagy in Human Neuroblastoma Cells. Biosci. Rep. 2018, 38, BSR20181822. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. The PI3K/Akt/MTOR Pathway as Therapeutic Target in Neuroblastoma. Curr. Cancer Drug Targets 2009, 9, 729–737. [Google Scholar] [CrossRef]
- Opel, D.; Poremba, C.; Simon, T.; Debatin, K.M.; Fulda, S. Activation of Akt Predicts Poor Outcome in Neuroblastoma. Cancer Res. 2007, 67, 735–745. [Google Scholar] [CrossRef]
- King, D.; Yeomanson, D.; Bryant, H.E. PI3King the Lock: Targeting the PI3K/Akt/MTOR Pathway as a Novel Therapeutic Strategy in Neuroblastoma. J. Pediatr. Hematol. Oncol. 2015, 37, 245–251. [Google Scholar] [CrossRef]
- Morgensztern, D.; McLeod, H.L. PI3K/Akt/MTOR Pathway as a Target for Cancer Therapy. Anti-Cancer Drugs 2005, 16, 797–803. [Google Scholar] [CrossRef]
- Li, X.; Tang, Y.; Yu, F.; Sun, Y.; Huang, F.; Chen, Y.; Yang, Z.; Ding, G. Inhibition of Prostate Cancer DU-145 Cells Proliferation by Anthopleura Anjunae Oligopeptide (YVPGP) via PI3K/AKT/MTOR Signaling Pathway. Mar. Drugs 2018, 16, 325. [Google Scholar] [CrossRef] [PubMed]
- Goyal, A.; Wang, Y.; Graham, M.M.; Doseff, A.I.; Bhatt, N.Y.; Marsh, C.B. Monocyte Survival Factors Induce Akt Activation and Suppress Caspase-3. Am. J. Respir. Cell Mol. Biol. 2002, 26, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, H.L.; Wang, Y.C.; Lu, Z.Y.; Yu, X.F.; Sui, D.Y. 20(S)-Protopanaxadiol-Induced Apoptosis in MCF-7 Breast Cancer Cell Line through the Inhibition of PI3K/AKT/MTOR Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 1053. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Al-Sammarraie, N.; DiPette, D.J.; Singh, U.S. Metformin Impairs Rho GTPase Signaling to Induce Apoptosis in Neuroblastoma Cells and Inhibits Growth of Tumors in the Xenograft Mouse Model of Neuroblastoma. Oncotarget 2014, 5, 11697–11709. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, J.; Wang, Z.; Yan, Z.; Qiao, M.; Ye, J.; Wei, Q.; Wang, J.; Wang, X.; Zhao, L.; et al. Antibiotic Monensin Synergizes with EGFR Inhibitors and Oxaliplatin to Suppress the Proliferation of Human Ovarian Cancer Cells. Sci. Rep. 2015, 5, 17523. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serter Kocoglu, S.; Sunay, F.B.; Akkaya, P.N. Effects of Monensin and Rapamycin Combination Therapy on Tumor Growth and Apoptosis in a Xenograft Mouse Model of Neuroblastoma. Antibiotics 2023, 12, 995. https://doi.org/10.3390/antibiotics12060995
Serter Kocoglu S, Sunay FB, Akkaya PN. Effects of Monensin and Rapamycin Combination Therapy on Tumor Growth and Apoptosis in a Xenograft Mouse Model of Neuroblastoma. Antibiotics. 2023; 12(6):995. https://doi.org/10.3390/antibiotics12060995
Chicago/Turabian StyleSerter Kocoglu, Sema, Fatma Bahar Sunay, and Pakize Nur Akkaya. 2023. "Effects of Monensin and Rapamycin Combination Therapy on Tumor Growth and Apoptosis in a Xenograft Mouse Model of Neuroblastoma" Antibiotics 12, no. 6: 995. https://doi.org/10.3390/antibiotics12060995